Основные типы морфологической структуры тела водорослей (М. М. Голлербах)
В отличие от высших растений, целиком и полностью характеризующихся одним листостебельным типом строения (другая структура у них вызвана вторичным упрощением), водоросли в пределах слоевцового типа строения обнаруживают исключительное морфологическое разнообразие. Тело водорослей, как уже упоминалось, может быть всех четырех степеней сложности, вообще известных для организмов,- одноклеточным, колониальным, многоклеточным и неклеточным. Размеры их в пределах каждой из этих форм отличаются огромным диапазоном - от микроскопических до очень крупных. Так, некоторые виды одноклеточной сине-зеленой водоросли синехоцистис (Synechocystis) едва достигают 1 мкм, одноклеточные зеленые водоросли из рода хлорелла (Chlorella) могут быть в 2 мкм, а длина клеток, образующих междоузлия в стеблевидных талломах харовых водорослей, часто составляет 15-20 см.
Однако самыми крупными размерами отличаются многоклеточные морские бурые водоросли, слоевища которых у отдельных видов, например у макроцистиса (Macrocystis pyrifera), могут достигать в длину 30-45 м.
Водоросли поражают многообразием своего внешнего облика. Вместе с тем все это исключительное многообразие имеет в своей основе несколько хорошо обособленных типов морфологической структуры, являющихся выражением главнейших ступеней морфологической дифференциации тела водорослей в процессе эволюции. Важно отметить, что эти ступени то в большей, то в меньшей степени повторяются в разных отделах водорослей, что свидетельствует об известном параллелизме эволюционного развития в пределах этих отделов.
В настоящее время различают 9 основных типов морфологической структуры тела водорослей. Из них 4 относятся к одноклеточным формам, 1 - к неклеточным, остальные 4 - к многоклеточным (колониальные формы, будучи существенным этапом на пути усложне-ния организации водорослей, все же являются лишь разновидностью одноклеточного строения).
1. Амебоидная структура представлена одноклеточными организмами, лишенными твердой клеточной оболочки и постоянной формы тела (рис. 15). Такие организмы, выпуская псевдоподии, способны совершать ползающие движения, подобно простейшим животным - амебам. Псевдоподии часто бывают длинными и тонкими, и тогда их называют ризоподиями (отсюда другое название этой структуры - ризоподиальная). Иногда несколько клеток подобного строения соединяются своими ризоподиями или даже сливаются в плазмодии.
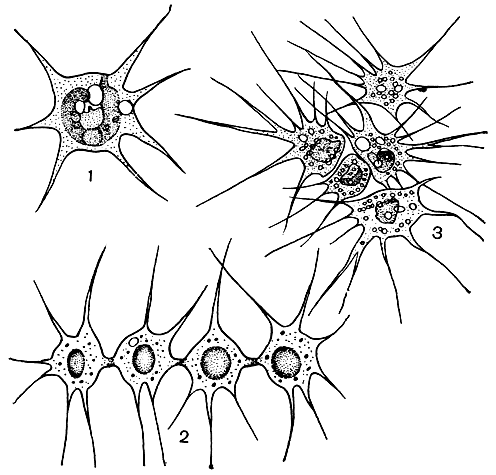
Рис. 15. Амебоидная структура у золотистых водорослей: 1 - одиночные клетки Chrysamoeba; 2 - рядовое объединение клеток Chrysidiastrum; 3 - групповое объединение клеток Rhizochrysis
Амебоидное строение могут временно приобретать некоторые одноклеточные водоросли, обладающие жгутиками (монадная структура, см. ниже), путем сбрасывания их или втягивания внутрь. Амебоидная структура свойственна ряду представителей пирофитовых, золотистых и желто-зеленых водорослей.
Амебоидная структура является наипростейшим типом строения одноклеточного организма, и есть все основания считать, что в процессе эволюции она была самым первым этапом при возникновении клеточного уровня жизни. Это не исключает, однако, того, что в некоторых случаях у современных водорослей она может быть следствием вторичного упрощения строения монадных форм.
2. Монадная структура свойственна одноклеточным организмам с твердой клеточной оболочкой или достаточно прочным перипластом и характеризуется наличием у таких клеток одного, двух или нескольких жгутиков, с помощью которых они активно двигаются в воде (рис. 16). Некоторые монадные формы вторично лишены жгутиков, и тогда, как правило, их тело способно менять свою форму (метаболия).
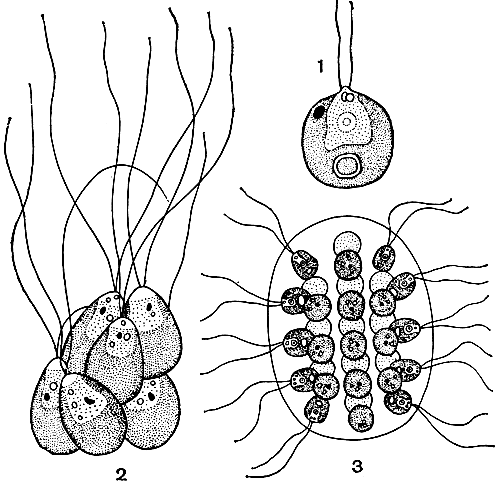
Рис. 16. Монадная структура у зеленых водорослей: 1 - одиночная клетка Chlamydomonas; 2 - колония Pyrobotrys, образованная срастанием клеток; 3 - колония Eudorina, образованная слизью
Монадная структура очень широко распространена в мире водорослей - она свойственна многим представителям в отделах пирофитовых, золотистых, эвгленовых, желто-зеленых и зеленых водорослей, причем у первых трех она является преобладающей.
Среди водорослей этого уровня организации часто встречаются колониальные формы, представляющие собой собрание отдельных клеток монадного строения, объединенных слизью в одно целое. Форма таких колоний и строение слизи у разных водорослей могут существенно различаться; в некоторых случаях клетки бывают связаны друг с другом плазмодесмами.
У большинства колониальных форм монадного строения клетки внутри колоний не обнаруживают различий, только у немногих из них наблюдается разделение на вегетативные, отличающиеся меньшими размерами, и на более крупные, служащие для размножения (например, у вольвокса и плевдорины из отдела зеленых водорослей).
Наконец, у многих водорослей во всех отделах, за исключением сине-зеленых и красных, независимо от структуры талломов в вегетативном состоянии, монадным строением обладают подвижные воспроизводительные клетки, служащие для бесполого размножения (зооспоры), а при наличии полового процесса - игаметы (мужские, а в некоторых случаях и женские).
Монадная структура многими альгологами принимается за первичную и исходную в филогенезе водорослей, однако имеется достаточно веских оснований считать ее вторичной, образовавшейся из амебоидной. Во всяком случае, несомненно, что монадная структура в процессе эволюции водорослей оказалась весьма прогрессивной и сохранилась у них на стадиях размножения вплоть до наиболее сложно организованных форм.
3. Коккоидная структура характеризуется отдельными клетками, снабженными твердой оболочкой и в вегетативном состоянии постоянно лишенными жгутиков или псевдоподиев (рис. 17). Вследствие такого устройства водоросли коккоидной структуры не способны к активному движению (исключение составляют только десмидиевые и диатомовые водоросли, но аппарат движения у них совершенно другой - выделение слизи), они или свободно живут и могут лишь пассивно переноситься водой, или прикрепляются к субстрату. Форма таких клеток необычайно разнообразна - от простой шаровидной до самой причудливой; оболочка у них гладкая или с различными выростами, цельная или пористая и т. д. Многим водорослям коккоидной структуры свойственно образование колоний различной формы, со слизью или без нее.
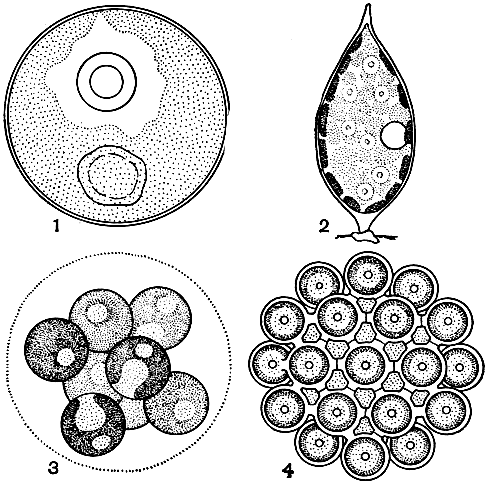
Рис. 17. Коккоидная структура у зеленых водорослей: 1 - одиночная клетка Chlorococcum; 2 - одиночная клетка Characium; 3 - колония Sphaerocystis, образованная слизью; 4 - колония Coelastrum, образованная срастанием клеток
Коккоидная структура чрезвычайно широко распространена у водорослей и в большей или меньшей степени наблюдается во всех отделах, где имеются одноклеточные формы. В отделе диатомовых водорослей, очень многочисленных и широко распространенных, она является единственной.
В эволюционном отношении коккоидная структура интересна как такая ступень морфологической дифференциации водорослей, на основе которой стало возможно возникновение многоклеточных талломов.
4. Пальмеллоидная структура представляет собой усложненный вариант коккоидной структуры, выражающийся в образовании водорослями достаточно крупных, преимущественно прикрепленных к субстрату слизистых тел определенной формы, содержащих внутри множество коккоидных клеток (рис. 18). Клетки объединяются в этой слизи чисто механически и плазматических связей не имеют. Такое строение широко распространено среди водорослей, но о пальмеллоидной структуре говорят лишь тогда, когда оно является постоянной формой их вегетативного роста; если же только временной стадией, то тогда его называют пальмеллевидным состоянием.
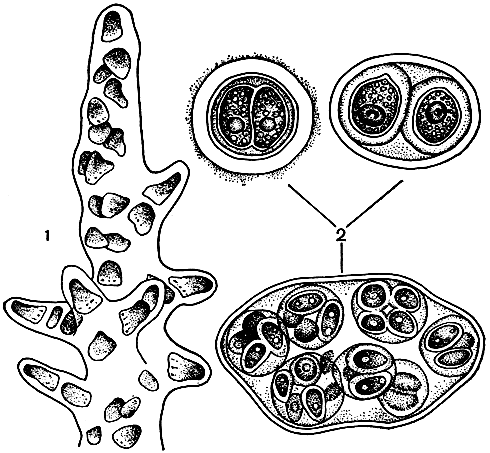
Рис. 18. Пальмеллоидная структура и пальмеллевидное состояние: 1 - пальмеллоидная структура у золотистой водоросли Hydrurus (часть таллома); 2 - пальмеллевидное состояние у зеленой водоросли Chlamydomonas
В пальмеллевидное состояние могут переходить многие одноклеточные подвижные и неподвижные водоросли (преимущественно при наступлении неблагоприятных условий); в меньшей степени оно наблюдается у водорослей иной структуры. Образующиеся при этом слизистые тела не достигают крупных размеров и не имеют определенной формы.
5. Нитчатая структура, как следует из названия, представлена талломами, состоящими из нескольких, нередко очень многих клеток, расположенных в форме нити. Нити могут быть простыми и разнообразно ветвящимися, свободноживущими, прикрепленными и объединенными в слизистые колонии (рис. 19).
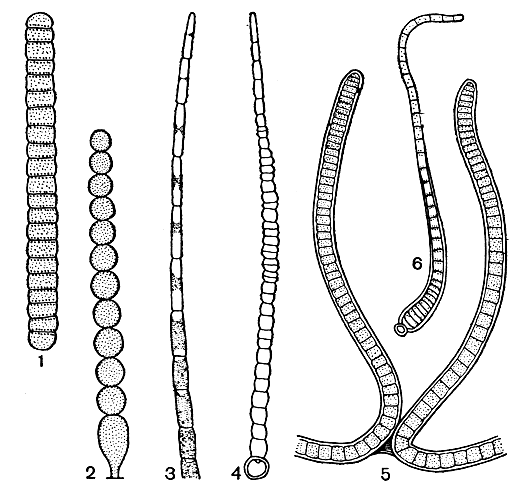
Рис. 19. Нитчатая структура у сине-зеленых водорослей: 1 - простейшее строение нити с диффузным ростом у Oscillatoria; 2 - нить с дифференцированным основанием у Endonema; 3 - верхушка нити у Rivularia, вытянутая в волосок; 4 - интеркалярный рост у Gloeotrichia; 5 - апикальный рост на концах ветвей у Scytonema; 6 - базальный рост у Calothrix
Нитчатая структура в мире водорослей является простейшей формой многоклеточного слоевища и свойственна огромному количеству представителей из разных отделов. Клетки в нитчатых слоевищах тесно связаны друг с другом, во многих случаях здесь доказано наличие пор и плазмодесм, проходящих через поперечные клеточные перегородки. Вместе с тем распадение нитей на участки и даже на отдельные клетки является обычным способом вегетативного размножения многих нитчатых водорослей.
В простейших случаях талломы нитчатой структуры слагаются из одного ряда клеток, вполне подобных друг другу. Наряду с этим многочисленны водоросли, у которых нити изменяются к концам - утончаются, или расширяются, или имеют конечные клетки, отличающиеся по форме от остальных. Различия в строении концов нитей бывают наиболее отчетливыми при прикрепленном образе жизни, определяющем полярность нити. При этом нередко нижняя клетка превращается в ризоид или стопу, а к вершине нити меняется форма клеток.
Рост нитчатых талломов происходит по-разному. Если способностью делиться обладают все клетки нити, то рост называют диффузным. В других случаях делятся не все клетки, а только часть их, и тогда рост нити происходит в определенных ее участках, называемых зоной роста или меристемальной зоной. В зависимости от положения меристемальной зоны на нити различают, кроме диффузного, еще три типа роста: интеркалярный, если зона роста расположена в средней части нити (обычно ближе к ее вершине), апикальный рост, осуществляемый делением преимущественно конечных клеток, и базальный рост, происходящий за счет деления клеток, расположенных у основания нити.
Нитчатая структура в эволюционном отношении интересна как исходная ступень для образования ряда более сложных многоклеточных структур, присущих многим водорослям, которые могут быть правильно поняты только как производное нити.
6. Разнонитчатая структура (иногда ее называют гетеротрихальной). Талломы данной структуры, будучи нитчатыми в своей основе, подразделяются на две части: стелющуюся по субстрату, горизонтальную, и прямо стоящую, вертикальную (рис. 20). В исходном варианте горизонтальная часть состоит из ветвящихся, обычно тесно расположенных нитей, которые или отчетливо различимы, или полностью смыкаются в сплошную псевдопаренхиматическую клеточную пластинку, нарастающую по периферии и уже не имеющую следов нитчатого строения. Вертикальную часть образует одна или многие, часто ветвящиеся нити, на которых обычно развиваются органы размножения.
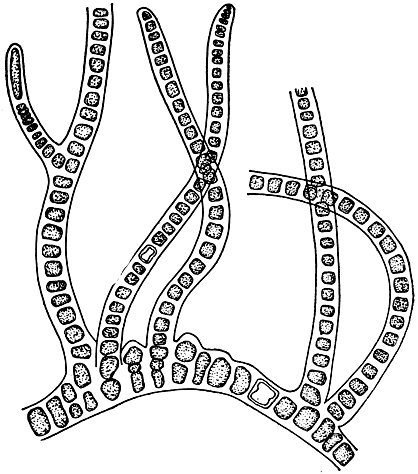
Рис. 20. Разнонитчатая структура у сине-зеленой водоросли Fischerella
В результате вторичных изменений - редукции или чрезмерного развития одной из этих частей - такое строение может резко нарушаться, что в некоторых случаях приводит к полной утрате характерных черт разнонитчатого строения. Так, при тесном соединении горизонтально расположенных нитей и более или менее полном исчезновении вертикальных нитей (иногда они сохраняются только в форме волосков) слоевище приобретает форму простого однослойного диска, целиком прикрепленного к субстрату.
В других случаях недоразвивается или полностью выпадает горизонтальная часть слоевища, тогда как вертикальные нити оказываются сильно развитыми и хорошо дифференцированными.
Разнонитчатая структура в том или ином выражении представлена в отделах сине-зеленых, зеленых, золотистых, бурых и красных водорослей, где она является или постоянной формой существования, или временным состоянием.
7. Пластинчатая структура характеризуется многоклеточными слоевищами в форме пластинок, состоящих из одного, двух или нескольких слоев клеток (рис. 21). Образование их всегда начинается с нити и происходит в результате продольного деления клеток, составляющих нить. Если возникающие продольные перегородки располагаются строго в одном направлении, то пластинка сохраняется однослойной; при возникновении перегородок, параллельных первоначальной пластинке, образуется два и более клеточных слоев. Двухслойные пластинчатые слоевища у некоторых водорослей еще на начальных стадиях их формирования превращаются в трубку или мешок вследствие расхождения слоев в середине при сохранении их связи по краям. В результате этого внутри образуется полость, а стенки становятся однослойными. Трубчатые слоевища по мере разрастания могут ветвиться.

Рис. 21. Пластинчатая структура у зеленой водоросли Prasiola: однорядная нить, разрастающаяся в однослойную пластинку
Пластинчатые водоросли растут либо свободно распростертыми по субстрату, либо прикрепленными к нему в одном месте края пластинки. Они хорошо представлены в отделах зеленых, бурых и красных водорослей.
8. Сифональная структура представляет собой особый тип строения, свойственный только некоторым водорослям и нередко называемый неклеточным. Отличительной чертой его является отсутствие внутри слоевищ клеточных перегородок при наличии большого количества ядер. Такие слоевища иногда достигают довольно крупных размеров, а внешнее их расчленение может быть очень сложным (рис. 22).
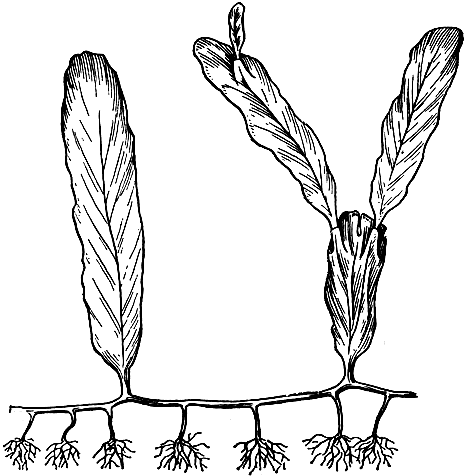
Рис. 22. Синофальная структура у морской зеленой водоросли Caulerpa (часть таллома)
Пресноводные водоросли сифональной структуры имеют вид или слабоветвящихся нитей, различимых простым глазом, или шаровидных телец размером с булавочную головку, снабженных разветвленными ризоидами. Сифональные представители морских водорослей отличаются большим разнообразием внешнего облика и подчас очень сложным расчленением слоевищ, принимающих у некоторых видов подобие деления на стебли, листья и корни. У таких сложных форм внутри обычно образуются впячивания стенок, придающие всему сифону механическую прочность.
Сифональная структура хорошо выражена в отделах зеленых и желто-зеленых водорослей. Предположения о путях ее образования до сих пор еще различны. Можно допустить независимое происхождение ее двумя путями: от водорослей коккоидной и нитчатой структуры, т. е. в результате усложнения одноклеточного и упрощения многоклеточного строения.
9. Харофитная структура, свойственная только харовым водорослям (отдел Charophyta), характеризуется крупными многоклеточными слоевищами линейно-членистого строения, состоящими из главного побега с ветвями и отходящими снизу ризоидами и сидящих на нем мутовками членистых боковых побегов, у некоторых форм ветвящихся (рис. 23). Места расположения мутовок на главном побеге называют узлами, части главного побега между узлами - междоузлиями. Междоузлия и членики боковых побегов всегда образованы только одной сильно вытянутой клеткой, но у многих видов рода хара (Chara) эти клетки снаружи обрастают еще одним слоем линейно расположенных дополнительных клеток, слагающих так называемую кору.
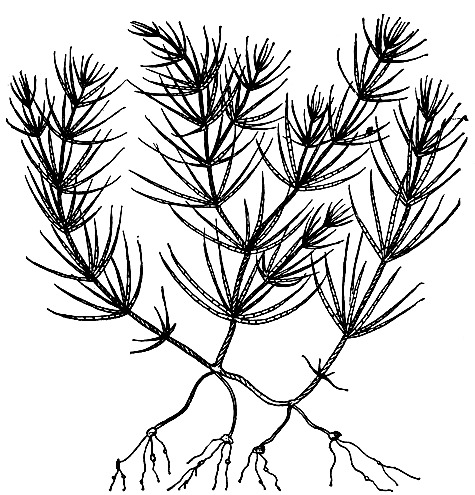
Рис. 23. Харофитная структура: молодой экземпляр Chara
Харофитная структура настолько своеобразна, что пути ее возникновения до сих пор остаются неясными, а в эволюционном отношении она является, несомненно, тупиковой.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'