Клетка водорослей (Т. В. Седова)
Клетка - основная структурная единица тела водорослей, представленных либо одноклеточными, либо многоклеточными формами. Совершенно уникальную группу составляют сифоновые водоросли: у них талломы не поделены на клетки, однако в цикле развития имеются одноклеточные стадии. Вполне очевидно, что клетка и здесь сохраняет свое значение как основной элемент, развитие и дифференциация которого приводят к формированию необычного слоевища.
Особенность одноклеточных форм определяется тем, что здесь организм состоит всего из одной клетки, поэтому в ее строении и физиологии сочетаются клеточные и организменные
черты. Это наложило отпечаток на характер изменчивости одноклеточных форм, наследование признаков, образование популяций и т. д. Представляя собой автономную систему, обладающую способностью к росту и самовоспроизведению, мелкая, не видимая простым глазом одноклеточная водоросль выполняет роль своеобразной фабрики, которая добывает сырье (поглощает из окружающей среды растворы минеральных солей и углекислоты), его перерабатывает и производит такие ценные соединения, как белки, углеводы и жиры. Кроме того, важным продуктом ее деятельности считается кислород. Таким образом, она активно участвует в круговороте веществ в природе. Одноклеточные водоросли иногда образуют временные или постоянные скопления в виде ценобиев и колоний.
Многоклеточные формы возникли после того, как клетка проделала длительный и сложный путь развития в качестве самостоятельного организма. В современных растениях сохранились следы этой истории. Переход от одноклеточного к многоклеточному состоянию сопровождался потерей индивидуальности и связанными с этим изменениями в структуре и функциях клетки. Внутри талломов многоклеточных водорослей складываются качественно иные отношения, чем между клетками одноклеточных водорослей. С возникновением многоклеточности связаны дифференцировка и специализация клеток в талломе, что следует рассматривать как первый шаг на пути становления тканей (гистогенез) и органов (органогенез). В зависимости от расположения клеток в талломе многоклеточные водоросли могут быть представлены нитчатыми или пластинчатыми формами.
Несмотря на более чем 300-летнюю историю изучения, клетка до сих пор не раскрыла всех своих тайн, так как "сцена", на которой здесь разыгрываются события, имеет микроскопические размеры, а действующие "герои" измеряются микрометрами и ангстремами. Этот миниатюрный, труднодоступный мир сказочно красив и удивительно гармоничен. Прижизненные наблюдения дают самые общие представления о клетке и ее строении. Для выявления деталей приходится прибегать к специальной обработке материала (фиксации, окрашиванию, удалению воды) и только после этого приступать к изучению в световом или электронном микроскопе (рис. 2).
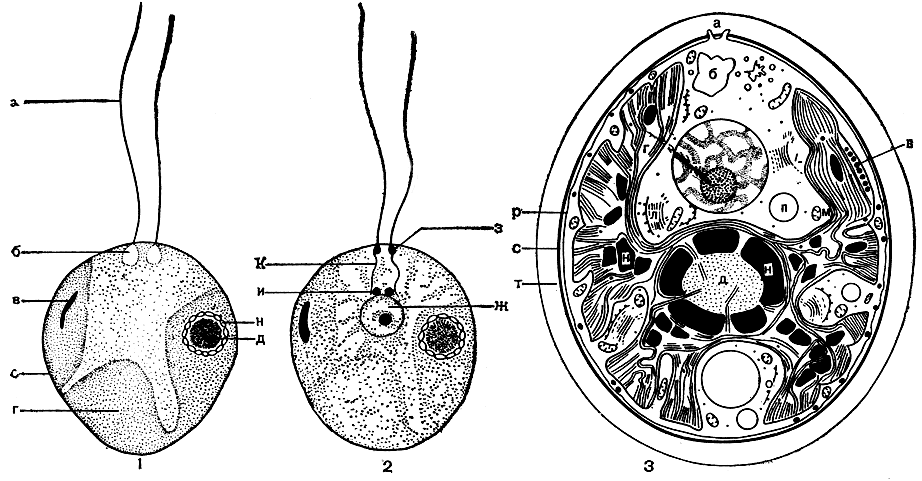
Рис. 2. Схематическое изображение клеточной организации одноклеточных зеленых подвижных водорослей при разных методах исследований: 1 - в прижизненном состоянии (световой микроскоп); 2 - после фиксации и окрашивания (световой микроскоп); 3 - по данным электронной микроскопии: а - жгутики, б - пульсирующие вакуоли, в - стигма, г - лопастной постенный хлоропласт, д - пиреноид, ж - ядро с ядрышком в центре, з - базальные тела, и - центриоль, к - ризопласты, л - диктиосома, м - митохондрии, н - крахмальные зерна, о - эндоплазматическая сеть, п - вакуоля, р - плазмалемма, с - оболочка, т - слизистая капсула
Даже при беглом знакомстве с водорослями бросается в глаза чрезвычайное разнообразие как формы (шаровидные, грушевидные, яйцевидные, веретеновидные, спиралевидные, дисковидные, бочонковидные, цилиндрические и сложные, не поддающиеся краткому описанию), так и размеров (всего от долей микрометра, как у сине-зеленых, до нескольких десятков сантиметров, как у харовых) их клеток. Наибольшая пестрота картин обнаруживается у свободноживущих одноклеточных водорослей.
У водорослей, в отличие от высших растений, встречаются клетки, содержимое которых (протопласт) окружено лишь тонкой, порядка 100 Å, пограничной мембраной (плазмалеммой). Такие клетки обычно называют голыми. Они не способны сохранять свою форму и постоянно находятся в метаболирующем, амебоидном состоянии. Подобного рода клетки встречаются как среди одноклеточных, так и многоклеточных водорослей, чаще всего на стадии гамет и зооспор.
Клетки некоторых водорослей (эвгленовых, перидиней, желто-зеленых), помимо плазмалеммы, окружены кожистым, эластичным слоем, толщина которого колеблется от 80 до 250 Å. Этот слой получил название пелликулы, или перипласта. Он состоит из фибриллярного вещества, в основном полисахаридной природы (главным образом из пектоз) и имеет сложную, многослойную организацию. Клетки с тонкой пелликулой обычно очень изменчивы по форме. Только толстая, похожая на панцирь пелликула может прочно ее зафиксировать. На поверхности пелликулы иногда возникают складки, выросты в виде зубцов или утолщения, называемые чешуйками. Эти структуры в различных сочетаниях образуют самые причудливые узоры, придавая организму неповторимый облик. Но главная их функция - повышение прочности клеточного покрова.
У представителей своеобразной группы кокколитофорид, входящих в состав золотистых водорослей, пелликула снаружи окружена дополнительным слоем округлых известковых телец - кокколитов. Они лежат в слизи, выделяемой протопластом.
Иногда в пелликуле обнаруживаются особые структуры в виде блях или шестиугольников, получившие название трихоцист. Обладая способностью скручиваться и раскручиваться, нити трихоцист при определенных условиях могут мгновенно распрямляться, "выстреливая" наружу. Предполагают, что трихоцисты выполняют в клетке защитную функцию и, может быть, даже участвуют в ее передвижении.
Еще более прочным, чем пелликула, оказался клеточный покров в виде теки. У некоторых вольвоксовых и хризомонад тека напоминает причудливой формы домик, в котором находится клетка. Она лежит там свободно, занимая только часть пространства (рис. 69, 3, 4; 195, 196). В структурном отношении тека - сложный мембранный комплекс, а по химическому составу представляет собой углеводное соединение, в основном состоящее из галактозы, галактуроновой кислоты и арабинозы. Часто, например, у перидиниевых водорослей, на наружном слое теки появляются шестиугольной формы пластинки, покрытые множеством мелких шипиков, бородавочек и других выростов. Пластинки располагаются в строгом порядке, срастаются своими краями и минерализуются, образуя сооружение, напоминающее панцирь. В месте срастания пластинок возникают швы - единственные участки поверхности, сохранившие способность к росту. В теке имеются поры, которые служат для циркуляции веществ между клеткой и внешней средой, для выхода наружу жгутиков и трихоцист.
В делении клетки тека не участвует. Обычно возникающие внутри ее дочерние клетки (или только одна из них) выходят наружу и образуют новую теку.
Важным событием, во многом предопределившим судьбу всего последующего развития растительной клетки, следует признать появление на ее поверхности покрова в виде оболочки - сначала пектиновой, а затем и целлюлозной природы. Преимущество этого образования состоит в том, что оно удачно сочетает защитную и опорную функции с возможностью ростовых процессов и проницаемостью.
Клеточные оболочки водорослей весьма разнообразны как по своему строению, так и по химическому составу. Толщина оболочки варьирует не только от вида к виду, но даже в пределах одного вида в зависимости от возраста клетки, ее функционального состояния и условий окружающей среды. Как правило, покоящиеся клетки имеют более толстые оболочки по сравнению с репродуктивными или молодыми вегетативными клетками (у зигнемы, например, они соответственно равны 1,6 и 0,8 мкм). Оболочки значительно утолщаются при дефиците влаги.
У подавляющего большинства водорослей оболочки оказываются сплошными, хотя, как у желто-зеленых, десмидиевых и диатомовых, встречаются и составные, слагающиеся из двух и более частей.
По времени заложения и особенностям роста различают первичные и вторичные оболочки (рис. 3, 2, 3). В активно делящихся клетках обычно образуется только первичная оболочка. Ее рост идет в двух направлениях: увеличивается преимущественно поверхность и в меньшей мере толщина. Переход клеток к покоящемуся состоянию сопровождается заложением снаружи от плазмалеммы вторичной оболочки, которая растет главным образом в толщину, отодвигает первичную оболочку от протопласта и вскоре становится преобладающей (рис. 3, 3). Она придает клетке прочность и окончательную форму. При возобновлении в клетке ростовых процессов вторичная оболочка подвергается гидратации, становится эластичной и получает возможность растягиваться.
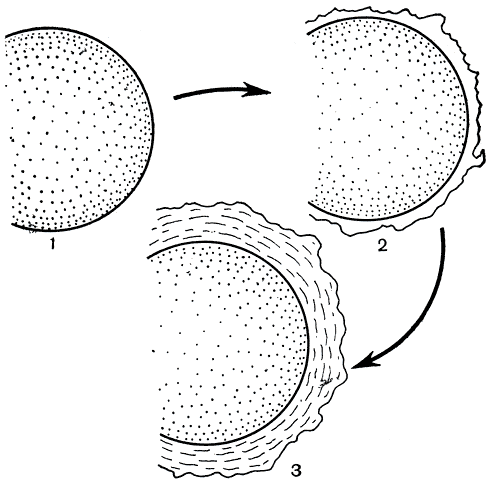
Рис. 3. Последовательные этапы формирования клеточной оболочки: 1 - молодая вегетативная клетка, лишенная оболочки; 2 - заложение первичной оболочки; 3 - появление под ней вторичной оболочки
На долю оболочки в среднем приходится 13-15% сухой массы клетки. У гидродиктиона, например, этот показатель еще выше и равен приблизительно 39%, при этом жиры составляют 7,2%, а остальное - белки и углеводы. Таким образом, белково-углеводный комплекс является основой оболочки, причем у водорослей соотношение этих компонентов колеблется в очень широких пределах (у сине-зеленых водорослей они представлены в одинаковой пропорции, а у хламидомонад отмечается сдвиг в сторону увеличения белков). Помимо этого, обращает на себя внимание разнообразный состав полисахаридов. В частности, у хламидомонады они образованы галактозами, арабинозами, в меньшей мере - ксилозами и рамнозами; у ацетабулярии - маннозами, глюкозами, галактозами и рамнозами; у ооцистиса - глюкозами, а галактозы, арабинозы и другие находятся в виде следов; у валонии - исключительно глюкозами. Создается впечатление, что природа решила поставить на водорослях грандиозный эксперимент по выявлению из множества полисахаридов того, который лучше всего подходил бы в качестве материала клеточных оболочек.
Даже при беглом взгляде бросается в глаза неоднородность, слоистость оболочки водорослевой клетки. Слои отличаются друг от друга по толщине, плотности и даже по химическому составу. Как правило, внутренние слои оказываются целлюлозными, а наружный - пектиновым. Он надежно защищает клетку от губительного влияния кислот и других столь же сильных реагентов (конечно, если это воздействие не будет слишком долгим).
Изучение оболочки водорослевых клеток в электронном микроскопе расширило наши представления относительно ее организации. Оболочка у подавляющего большинства видов оказалась двухкомпонентной системой, состоящей из аморфного основного вещества (стромы) и погруженных в него фибрилл целлюлозной природы с сечением 200 - 300 Å, которые выполняют роль опорного элемента и повышают прочность всей конструкции (рис. 4, 1, 2, 3).
Общее количество фибрилл и характер их ориентации варьируют не только в зависимости от формы клетки, но и от слоя к слою в пределах одной и той же клетки. Все попытки найти этому объяснение, исходя из теории механических напряжений, окончились неудачно, поскольку не учитывались "творческие способности" самой клетки, живущей по биологическим законам.
Изучение оболочек на ультраструктурном уровне позволяет произвести и более точное расчленение оболочек на слои. В большинстве случаев их не больше двух-трех, но бывают и отклонения. У сине-зеленых водорослей оболочка, как правило, четырехслойная, причем у осциллатории все слои одинаковой толщины (100 Å), а у анабены существуют различия, и весьма значительные (I -100 Å; II - 140 Å; III - 150-300 Å; IV - 140 Å). Многослойные оболочки обнаружены у десмидиевых водорослей. Однако не у всех водорослей слоистость оболочки выявляется четко, что, вероятно, служит причиной появления порой весьма противоречивых данных. Так, одни исследователи сообщают о существовании у ценобиальной зеленой водоросли сценедесмус всего двух слоев, а другие - трех.
Отсутствие целлюлозы делает оболочки гомогенными, что нередко наблюдается у вольвоксовых (гематококк, вольвокс), улотриксовых (спонгоморфа, акросифония), сифоновых (кодиум).
Иногда там, где фибриллярный компонент отсутствует или развит очень слабо, оболочки интенсивно пропитываются (инкрустируются) находящимся в среде раствором солей железа (у вольвоксовых), кальция (у харовых) или кремния (у диатомовых), которые укрепляют матрикс, создавая нередко структуру, похожую на панцирь. Соли, как известно, в отличие от целлюлозы, упорядочиваются не в линейные, а в трехмерные структуры, что делает оболочку необычайно красиво орнаментированной. Нередко оболочки пропитываются органическими соединениями, из которых самыми распространенными являются лигнин и кутин, секретируемые протопластом клетки. Причем кутин выполняет, помимо опорной, еще и защитную функцию, поскольку задерживает губительные ультрафиолетовые лучи и предохраняет клетку от излишней потери воды, тем самым улучшая условия транспирации.
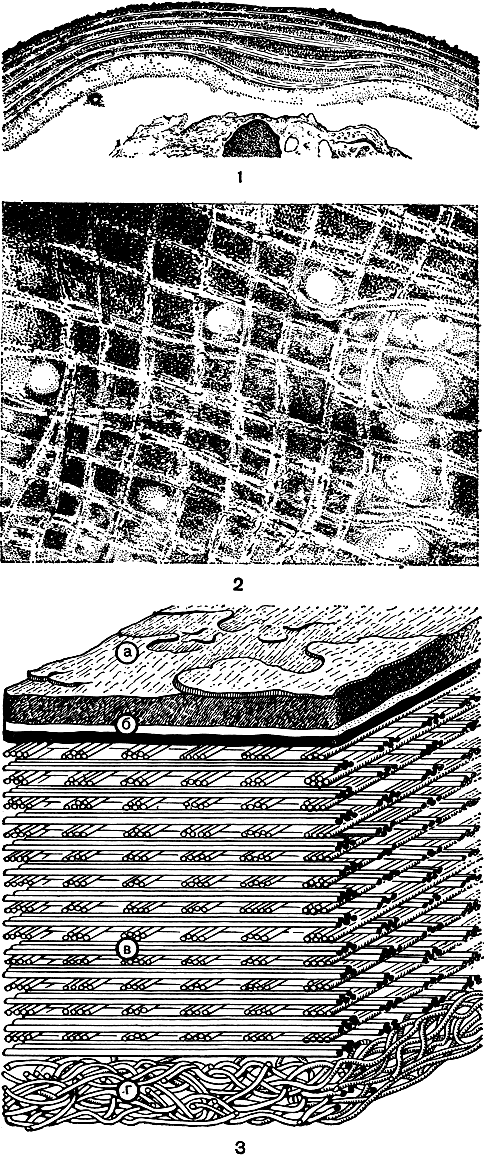
Рис. 4. Организация клеточной оболочки по данным электронной микроскопии: 1 - многослойная оболочка Glaucocystis nostochinearum; 2 - расположение целлюлозных фибрилл в оболочке Pyrocystis lunula; 3 - модель строения оболочки: а - внешняя часть наружного слоя, б, в, г - внутренний слой оболочки с разным характером упорядоченности целлюлозных фибрилл
От инкрустации следует отличать процесс адкрустации, под которым понимают скопление вещества на поверхности клетки. Так, кутин, выделяясь наружу, затвердевает и образует кутикулу. Подобного рода образование возникает на вегетативных клетках у порфиры, на оогониях у эдогониума, на эпидермальных клетках у многих бурых водорослей.
К адкрустирующим веществам относятся и всевозможные слизи. У некоторых сине-зеленых водорослей слизистый слой, называемый здесь влагалищем, бывает довольно массивным, порядка 0,5-1,0 мкм. Мощная слизистая капсула появляется у многих одноклеточных зеленых водорослей (пальмелла, диктиосфериум, факотус), десмидиевых и в других группах. Она, очевидно, способствует созданию вокруг клетки своеобразного микроклимата. Слизь представляет собой продукт жизнедеятельности оболочки и по своей природе может быть либо пектиновой, либо целлюлозной. От постоянных слизистых форм следует отличать организмы с временно ослизняющимися, как у хламидомонад, оболочками. О таких водорослях обычно говорят, что они находятся в пальмеллевидном состоянии, которое, по всей вероятности, помогает им переживать всевозможные неблагоприятные условия.
У красных водорослей на поверхности оболочки откладываются в большом количестве сульфированные полисахариды, образующие вокруг клетки мощную капсулу. Поскольку эти полисахариды водорастворимы, то их отложение идет непрерывно. Из подобного рода соединений получают ценный продукт агар-агар, который находит широкое применение, в частности, в кондитерской промышленности.
Выделения протопласта поступают на поверхность клетки через особые отверстия в оболочке - поры. Их диаметр у десмидиевых равен приблизительно 120-200 Å, у фукуса - 370 Å, у осциллатории - 300-600 Å, иногда и до 900 Å. В среднем на 1 мкм2 поверхности приходится до 20-30 пор. Они размещаются рав-номерно или группами. Если у одноклеточных водорослей благодаря порам устанавливается прямая связь протопласта с внешним миром, то у многоклеточных через поры на внутренних стенках - и с соседними клетками таллома. Иногда через эти поры беспрепятственно проходят цитоплазматические тяжи (плазмодесмы) (рис. 5, 1). Они, по сути, связывают протопласты всех клеток таллома в единое целое и обеспечивают однозначную реакцию многоклеточного организма на изменения в окружающей среде. Не случайно именно у тех водорослей, у которых существуют подобного рода межклеточные коммуникации, можно иногда наблюдать почти фантастическую картину, как, точно по мановению дирижерской палочки, все клетки нити одновременно приступают к образованию то акинет, то зооспор.
Сравнительное изучение особенностей заложения, развития и строения пор показало, что они различаются по происхождению: одни возникают в ходе клеточного деления, как результат неполного срастания перегородки, другие образуются энзиматически, путем разрушения оболочки. Впоследствии поры могут закупориваться пробкой, как это нередко наблюдается у сине-зеленых (в особых клетках - гетероцистах, о которых рассказано ниже) и красных водорослей (рис. 5, 2).

Рис. 5. Строение плазмодесм и гетероцист: 1 - плазмодесмы в оболочках зеленой водоросли Coleochaete (показаны стрелкой); 2 - продольный срез через гетероцисту сине-зеленой водоросли Anabaena azollae: а - пробка, б - полярное утолщение оболочки гетероцисты
Помимо пор, оболочки многих водорослей снабжаются различного рода выростами в виде щетинок, шипиков и чешуек. Их роль для клетки неоднозначна: в одних случаях они выполняют, очевидно, защитную функцию, а в других обеспечивают оптимальные условия жизни, например поддерживают организмы в состоянии парения.
В протопласте любой клетки различают два тесно связанных друг с другом компонента: ядро (карион, или нуклеус, что в переводе соответственно с греческого и латинского означает "ядро") и цитоплазму, причем от степени их развития зависит уровень организации клетки в целом. Водоросли - единственная группа растений, где представлены все известные в настоящее время типы клеточной организации: прокариотическая - у сине-зеленых водорослей, мезокариотическая - у панцирных жгутиконосцев из группы динофлагеллат, эукариотическая - у водорослей остальных отделов.
У прокариот отсутствует морфологически оформленное ядро, а его функции выполняет состоящий из микрофибрилл ДНК нуклеоид, который, несмотря на примитивность своей организации, не только содержит, но и точно передает из поколения в поколение всю генетическую информацию о клетке. Более того, прокариоты лишены и многих типичных плазматических органелл (хлоропластов, митохондрий, диктиосом), но зато снабжены многочисленными и разнообразными как по форме, так и по составу включениями.
Мезокариоты уже обладают четко дифференцированным ядром, однако в его строении и поведении сохранились некоторые черты примитивности, присущие нуклеоиду. Подобная двойственность проявляется и в других чертах организации клетки, что позволяет говорить о промежуточном положении этих организмов между примитивными прокариотами и достигшими в целом наиболее высокого уровня развития эукариотами.
У большинства водорослей в клетке присутствует всего одно ядро, но известны случаи, когда их бывает два-три и больше. Клетки с несколькими десятками, как у кладофоры, или сотнями, как у водяной сеточки (гидродиктион), ядер называют ценоцитными. Примечательно, что эти водоросли возвращаются к одноядерному состоянию при образовании специализированных клеток бесполого (апланоспоры, зооспоры) и полового (гаметы) размножения.
Форма, размеры и местоположение ядра в клетке сильно варьируют у разных водорослей, а также в зависимости от фазы развития, на которой находится тот или иной организм.
Очень мелкие ядра (диаметром 1-3 мкм) характерны для подавляющего большинства одноклеточных водорослей, хотя и здесь встречаются такие "рекордсмены", как гематококк (10 мкм), гимнодиниум (12-16 мкм), вольвокс (13-17 мкм), эремосфера (25-30 мкм). У многоклеточных водорослей наряду с мелкими ядрами (1,5-2,5 мкм у улотрикса, 2,5-3,5 мкм у стигеоклониума, 2,0-4,0 у уронемы) попадаются крупные (35-57 мкм у некоторых видов спирогиры) и даже такие гигантские, как у ацетабулярии (1,7мм).
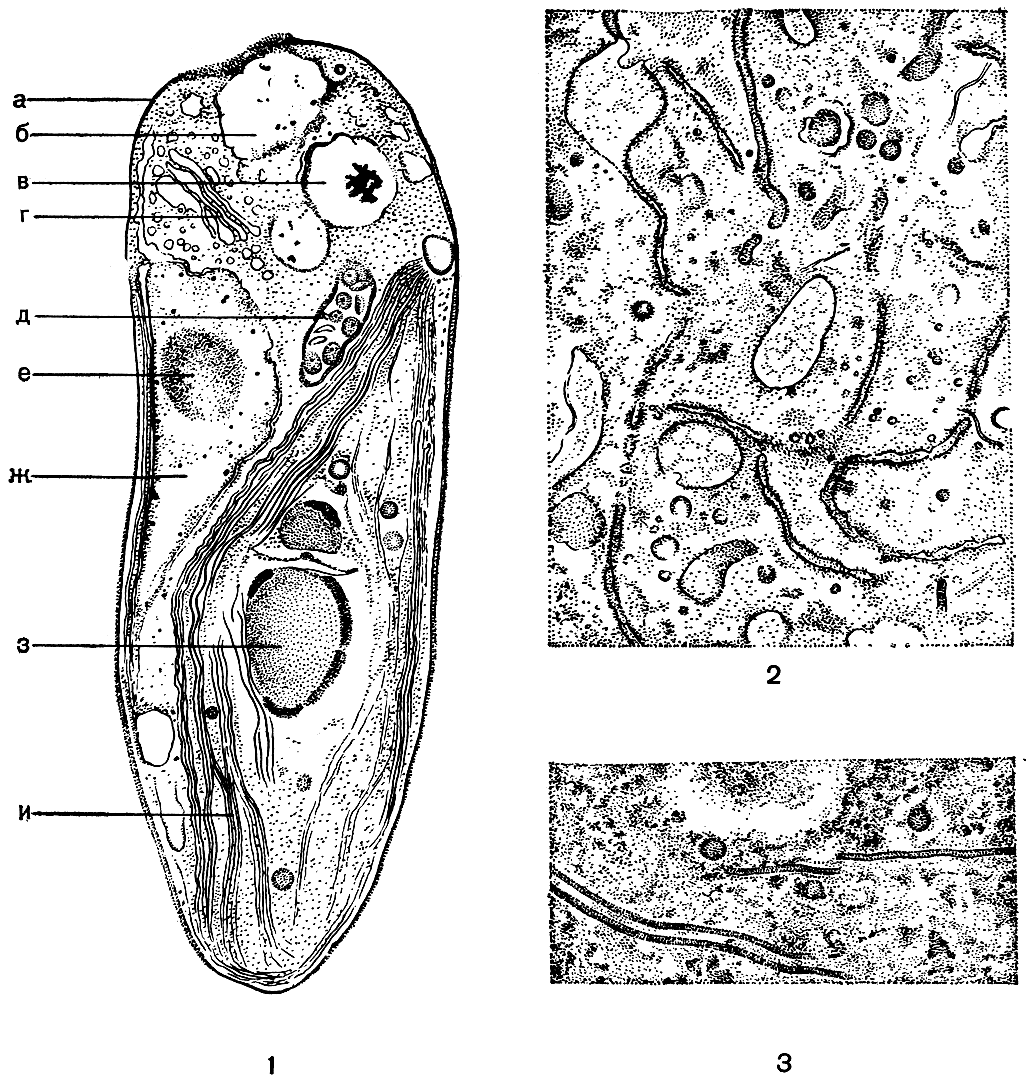
Рис. 6. Ультратонкое строение клетки: 1 - продольный срез через клетку одноклеточной зеленой водоросли Pedinomonas minor (электронный микроскоп): а - плазмалемма, б - пульсирующая вакуоля, в - статическая вакуоля, г - диктиосома, д - митохондрия, е - ядрышко, ж - окруженное двухмембранной оболочкой ядро с ядрышком, з - пиреноид с крахмальной обкладкой, и - хлоропласт с пучком ламелл; 2 - эндоплазматическая сеть в цитоплазме клетки спирогиры; 3 - микротрубочки в цитоплазме клетки спирогиры
Размеры ядер даже в пределах одного таллома могут существенно колебаться. Так, у одного вида бриопсиса (Bryopsis halymeniae) из сифоновых ядра спорангия имеют диаметр, равный 30-45 мкм, а ядра вегетативных клеток таллома - 3,0-4,5 мкм. У другого вида бриопсиса (Bryopsis hypnoides) ядра в проростке зиготы оказываются крупнее ядра зиготы почти в 1700 раз. У водяной сеточки в молодых клетках ядра не превышают в диаметре 2,8 мкм, а в старых иногда достигают 16 мкм. Интересно, что перед делением диаметр ядер обычно увеличивается более чем в 2 раза. В этот же период происходит и его перемещение, как правило, в центр (эдогониум) или в переднюю часть клетки (эвглена).
В ядре у водорослей выявлены те же самые структуры, что и в ядрах других эукариотов: оболочка, ядерный сок (нуклеоплазма, кариоплазма, кариолимфа), ядрышко (нуклеолус), включения хроматина.
Оболочка ядра состоит из двух мембран (внутренней и наружной) и имеет поры (рис. 6, 1 ж). Толщина оболочки, например, у хлореллы (Chlorella pyrenoidosa) равна 300-450 Å, у клостериума (Closterium ehrenbergii) - 200-300 Å, у ацетабулярии (Acetabularia mediterranea) - 200 Å. Диаметр пор у клостериума Эренберга составляет 700 Å, у ацетабулярии - 800 Å. Наличие пор обусловливает высокую проницаемость оболочки ядра и способствует свободному прохождению через нее крупных макромолекул. Однако оболочка обладает очень высокой избирательностью и пропускает через себя далеко не все из них. Поэтому не следует представлять ее в виде решета, которое чисто механически отсеивает все то, что не соответствует размеру его отверстий. В силу этого о проницаемости оболочки приходится говорить как о биологическом процессе, проходящем при непосредственном участии самой структуры.
Сильные преобразования претерпевает оболочка ядра во время ядерного деления, которое часто, хотя и не всегда, сопровождается сначала ее фрагментацией, а затем и полным распадением.
Все внутреннее пространство ядра заполнено (рис. 7, 1-6) ядерным соком, в котором, помимо ядрышка, удается различить хроматиновые структуры в виде скрученных нитей и глыбок; самые крупные глыбки иногда называют хромоцентрами, кариосомами или ложными ядрышками по чисто морфологическому сходству с последними. Теперь считают, что хроматиновые нити не что иное, как форма существования хромосом в покоящемся ядре, а хромоцентры - их более плотные участки. Обычно хроматин скапливается вдоль оболочки; остальная же часть ядра выглядит почти гомогенной, поскольку там лишь иногда удается обнаружить структуру в виде нежной сеточки с гранулами в ее узлах. Только у очень немногих водорослей (эвглена, кладофора) хроматиновые образования распределяются по всему ядру более или менее равномерно, маскируя его содержимое (рис. 7, 5, 6).
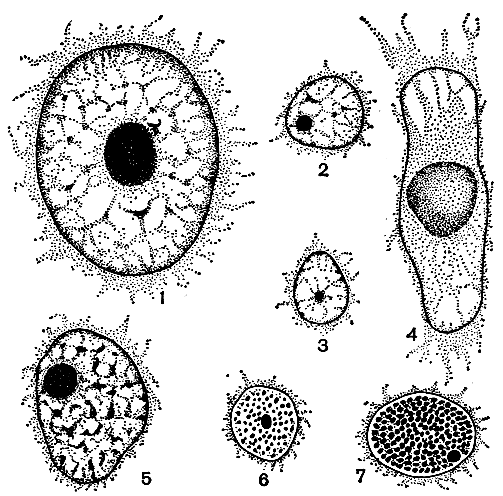
Рис. 7. Интерфазные ядра клеток у разных водорослей: 1 - Microspore; 2 - Chlamydomonas; 3 - Hydrodictyon; 4 - Spirogyra; 5 - Cladophora; 6 - Colacium; 7 - Glenodinium
Переход ядер к делению сопровождается усилением спирализации, укорачиванием, а затем и формированием морфологически четко выраженных хромосом. Исключение составляют мезокариотные водоросли: у них хромосомы находятся постоянно в конденсирован-ном состоянии и поэтому всегда хорошо видны (рис. 7, 7).
Хромосомы ядра в совокупности составляют хромосомный набор. Каждый вид организмов имеет свой набор строго определенных по форме, размерам и количеству хромосом (кариотип). Он служит своего рода паспортом, куда занесены в зашифрованном виде все основные исходные данные об организме. Их расшифровка сопряжена с большими трудностями, и здесь предстоит еще очень большая работа. У водорослей в подавляющем большинстве случаев хромосомы очень мелкие, без перетяжек, и имеют вид точек или палочек. В пределах одного набора они, как правило, выглядят все "на одно лицо", поэтому изучать их бывает очень трудно (рис. 8, 1-7).
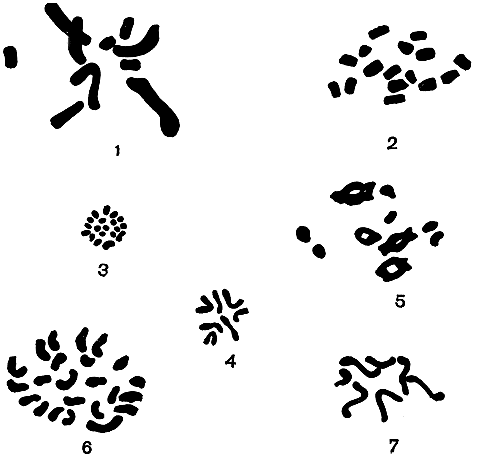
Рис. 8. Хромосомные наборы у разных водорослей: 1 - улотрикс (n = 10); 2 - микроспора ( n = 16); 3 - гидродиктион (n = 19); 4 - эвдорина (n = 7); 5 - ульва (n = 10); 6 - трентеполия (n = 24); 7 - вольвулина (n = 7)
Каждое ядро обычно содержит одно, но иногда и большее количество ядрышек. Обычно они бывают диаметром 1-2 мкм, хотя встречаются и более крупные. Например, у спирогиры (Spirogyra neglecta) они достигают 11-14 мкм. Ядрышки состоят из очень плотного вещества и лишены мембранного барьера. В них, в отличие от хроматина, скапливается не ДНК, а РНК. Во время ядерного деления ядрышки претерпевают циклические преобразования, связанные с разрыхлением, потерей красящей способности и диспергированием (рассеиванием) во время первой стадии ядерного деления (профазы) и конденсацией, восстановлением красящей способности с полной реконструкцией своего облика к концу этого процесса (в телофазе).
Однако среди водорослей встречаются отдельные группы, где имеются отклонения от подобной схемы, которая признается классической, поскольку присуща подавляющему большинству организмов не только растительного происхождения. Здесь в первую очередь следует назвать эвгленовые водоросли, где ядрышко делится без предварительного разрыхления путем гантелевидной перетяжки на два дочерних (рис. 199). Несколько иначе ведут себя ядрышки у спирогир: они в той или иной степени разрыхляются, слегка диспергируются, но не теряют при этом своей красящей способности. В результате ядро оказывается заполненным так называемым ядрышковым веществом, ко-торое очень затрудняет наблюдение за ходом ядерного деления, часто маскируя его детали.
Совокупность всех преобразований, связанных с делением ядра, составляет митотический цикл, или митоз. У водорослей, как и у других растений, реорганизация ядра следует в строгой последовательности в несколько этапов (фаз). Всего различают 5 фаз: интерфаза, или состояние покоя (самая продолжительная по времени), профаза, метафаза, анафаза и телофаза. Из телофазы ядро снова переходит в интерфазу.
Второй компонент клетки - цитоплазма - состоит из гомогенного основного вещества (стромы) и погруженных в него телец (органелл) различного размера и формы: митохондрий, диктиосом, эндоплазматической сети и т. д. Этот основной набор органелл присущ клеткам не только растений, но и животных.
Митохондрии (рис. 6, 1 д) обычно имеют овальную форму; они покрыты двух-мембранной оболочкой, причем внутренняя мембрана образует выросты - кристы. Митохондрии располагаются преимущественно в тех местах клетки, где проходят процессы, связанные с затратой энергии (около ядра, у основания жгутиков, вокруг стигмы), и выполняют роль дыхательных центров. Число, размеры митохондрий и степень развития крист взаимосвязаны друг с другом и с функциональным состоянием клетки.
Диктиосомы (рис. 6, 1 г) состоят из собранных в стопки уплощенных мешочков, или цистерн, число которых иногда достигает нескольких десятков. От мешочков по краям отпочковываются разного размера пузырьки. Диктиосомы на поперечных срезах имеют вид характерных коротких, компактных пучков. Число диктиосом в клетке очень варьирует, и всю их совокупность там называют аппаратом Гольджи, в честь ученого, впервые наблюдавшего эти структуры еще в прошлом веке. Диктиосомы осуществляют синтез полисахаридов, которые затем переходят в пузырьки и транспортируются в разные концы клетки.
Вся цитоплазма пронизана в той или иной степени развитой сетью каналов. Местами они образуют расширения - цистерны, иногда довольно значительные - пузырьки. Вся эта система получила название эндоплазматической сети, или эндоплазматического ретикулума (рис. 6,2). Эндоплазматическая сеть осуществляет синтез главным образом белков и транспортирует их преимущественно в те места, где развертываются интенсивные формообразовательные процессы. Отличительной особенностью клеток водорослей следует признать слабое развитие эндоплазматической сети. У желто-зеленых, золотистых, бурых, диатомовых и красных водорослей она представлена единственным каналом, огибающим хлоропласт и затем переходящим в наружную мембрану ядерной оболочки, за счет чего устанавливается тесная взаимосвязь между ядром и хлоропластом (рис. 9).
Совсем недавно в цитоплазме клеток различных организмов, в том числе и водорослей, обнаружены короткие (по сравнению с каналами эндоплазматической сети) образования с жесткими гладкими контурами, получившие название микротрубочек (рис. 6, 3). В сечении они имеют вид цилиндров с диаметром просвета 200-350 Å. Микротрубочки оказались крайне динамичными структурами: они могут то появляться, то исчезать, перемещаться из одного района клетки в другой, увеличиваться или уменьшаться в числе. Они сосредоточиваются преимущественно вдоль плазмалеммы (самый наружный слой цитоплазмы), а в период клеточного деления перемещаются в область формирования перегородки. Их скопления обнаруживаются также вокруг ядра, вдоль хлоропласта, около стигмы. Последующие исследования показали, что эти структуры присутствуют не только в цитоплазме, но и в ядре, хлоропласте, жгутиках.
Функции микротрубочек до конца не выяснены. Возможно, они участвуют в транспорте веществ по клетке, а характер локализации побуждает рассматривать их как цитоскелетные образования, призванные поддерживать форму как отдельных органелл, так и клетки в целом.
В клетках водорослей, как впрочем и других растительных организмов, присутствуют также лизосомы, где отмечается скопление большого набора ферментов из числа гидролаз, способных расщеплять важнейшие классы химических соединений, включая белки, углеводы, нуклеиновые кислоты и т. д. Имеются пероксисомы, в составе которых обнаружены каталаза и целый ряд других ферментов пероксидазного комплекса; сферосомы, содержащие гидролитические ферменты, липиды и ароматические аминокислоты. Из всего этого видно, что цитоплазма клеток водорослей достигает очень высокой степени структурной организации.
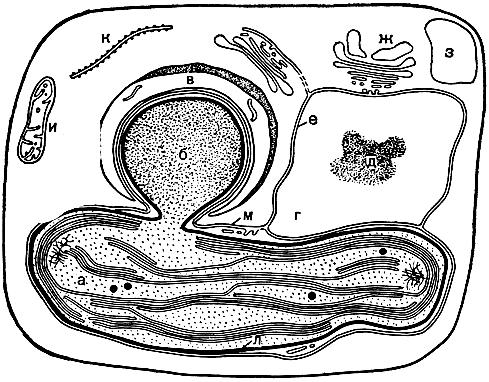
Рис. 9. Схема структурной связи между ядром, эндоплазматической сетью и хлоропластом у бурых водорослей: а - хлоропласт с ламеллами, собранными в пучки по 3; б - пиреноид; в - обкладка пиреноида; г - ядро; д - ядрышко; е - ядерная оболочка, состоящая из двух мембран; ж - диктиосома; з - вакуоля; и - митохондрия; к - фрагмент канала эндоплазматической сети; л - оболочка хлоропласта, образованная двумя мембранами; м - эндоплазматическая сеть, окружающая хлоропласт и связанная с наружной мембраной ядерной оболочки
В интенсивно растущих клетках, помимо перечисленных органелл, можно наблюдать хорошо развитую систему пузырьков, или вакуолей. Они обладают способностью сливаться друг с другом, что иногда приводит к возникновению одной большой вакуоли, располагающейся обычно в центре клетки. Ее появление компенсирует тот недостаток цитоплазмы, который возникает при резком увеличении объема клетки в период ее интенсивного роста.
В свете данных электронной микроскопии по-новому решается вопрос о природе вакуолей. Как теперь выясняется, в их образовании принимает участие не только аппарат Гольджи и эндоплазматическая сеть, но даже плазмалемма. Среди многочисленных функций, которые выполняет вакуолярная система в клетке, немаловажное место занимает функция осморегуляции.
К особому типу вакуолей в клетках некоторых водорослей следует отнести пульсирующие вакуоли (рис. 2, 1б; 6, 1б). Они, как правило, появляются в подвижных клетках, снабженных жгутиками, но иногда (у некоторых зеленых и диатомовых) образуются и в неподвижных клетках. Эти вакуоли обладают способностью ритмично пульсировать, т. е. попеременно сокращаться и расширяться. В простейших случаях (у многих золотистых и зеленых водорослей) имеется одна или несколько простых вакуолей, при своем сокращении выбрасывающих содержащуюся в них жидкость в цитоплазму или через специальные выводные канальцы за пределы клетки. В других (например, у эвгленовых) - налицо целая система вакуолей, состоящая из главной вакуоли, в которую открывается одна или несколько вакуолей второго порядка, часто, в свою очередь, окруженных вакуолями третьего порядка. При сокращении последних жидкость поступает в вакуоли второго порядка, которые, сокращаясь, наполняют главную вакуолю. Выведение жидкости из главной вакуоли осуществляется через выводной канал. У некоторых перидиней обнаружено еще более сложное устройство системы сократительных вакуолей.
Функция пульсирующих вакуолей - удаление из клетки излишка воды. Наряду с этим пульсирующие вакуоли, по-видимому, выполняют и экскреторную функцию, выбрасывая из клетки ненужные продукты обмена веществ. И все же следует признать, что пульсирующие вакуоли еще во многом остаются загадочными образованиями: они могут то появляться, то исчезать; непонятны и причины их сокращения.
Своеобразную группу вакуолей составляют так называемые газовые вакуоли, присутствующие в клетках сине-зеленых водорослей. Они имеют вид полостей, ограниченных тонкой мембраной и заполненных газом. Наличие газовых вакуолей, несомненно, уменьшает удельный вес организма, и он поднимается в верхние слои воды. Именно в этом многие усматривают биологическое значение газовых вакуолей. Такое объяснение было бы полностью приемлемо, если бы не случаи обнаружения подобных образований и у глубоководных водорослей. В связи с этим было высказано предположение, что в условиях глубинной жизни при дефиците кислорода начинается внутриклеточное брожение, приводящее к образованию газовых вакуолей. С этой точки зрения "парение" представляется явлением вторичным.
Газовые вакуоли начинают усиленно формироваться при повышении интенсивности света. Очевидно, в этом случае они выполняют защитную функцию, отражая часть света и тем самым предохраняя клетку от его избытка.
В отличие от животных (гетеротрофных) организмов растения принадлежат к автотрофам, т. е. обладают способностью образовывать органические вещества из неорганических в процессе фотосинтеза. Правда, некоторые водоросли из числа эвгленовых, одноклеточных зеленых, желто-зеленых и пирофитовых водорослей утрачивают эту способность и переходят к гетеротрофному способу питания, но это свойство у них уже вторичного происхождения.
У водорослей фотосинтетический аппарат представлен разными уровнями организации: от морфологически неоформленного (у сине-зеленых) до типичных хлоропластов (у зеленых водорослей).
Хлоропласты, первоначально названные хроматофорами (и сейчас еще этот термин сохраняется в научной литературе), представляют собой специфические органеллы растительной клетки, несущие зеленый пигмент хлорофилл, существующий в нескольких формах (а, b, с, d, е). Помимо него, в хлоропластах у красных, желто-зеленых, бурых и других водорослей присутствуют и такие пигменты, как фикоцианин, фикоэритрин, фикоксантин, β - каротин, ксантофилл и т. д., которые часто маскируют основную зеленую окраску красной, желто-зеленой, золотистой, бурой и т. д.
В противоположность высшим растениям хлоропласты водорослей отличаются большим разнообразием по форме, числу, местоположению в клетке и, как теперь выясняется, даже по внутренней организации. Хлоропласты у водорослей могут быть чашевидными, лентовидными, спиралевидными, пластинчатыми, звездчатыми и т. д. Как правило, в подвижных клетках у зеленых водорослей присутствует всего один хлоропласт; у водорослей из других отделов их бывает два и больше; у эвгленовых и динофлагеллат в молодых клетках - от 50 до 80, а в старых 200-300. Хлоропласты занимают в клетке либо центральное, либо постенное положение.
Хлоропласты у водорослей, как и у других растений, окружены оболочкой и состоят из основного вещества (стромы) и погруженных в него пластинчатых, или ламеллярных, структур, а также различного рода включений, отличающихся по размерам, форме и составу своего содержимого (рис. 6, 1и).
Оболочка хлоропласта толщиной порядка 100-150 Å образована двумя одиночными мембранами. Иногда у бурых, золотистых, желто-зеленых, диатомовых водорослей этот барьер усложняется за счет примыкающего к хлоропласту канала эндоплазматической сети, в результате чего вокруг него возникает четырехмембранная система, а у панцирных жгутиконосцев из группы динофлагеллат и эвгленовых водорослей мембрана канала эндоплазматической сети настолько тесно прилегает к наружной мембране оболочки хлоропласта, что граница между ними становится неразличимой, создавая полное впечатление существования здесь трехмембранной системы.
Расположенные в строме пластинчатые структуры (ламеллы) по виду напоминают миниатюрные уплощенные мешочки, которые называют тилакоидами (греч. "тилакоидес" - мешковидный) или дисками. Они придают хлоропласту характерную слоистость, что очень хорошо видно на поперечных срезах. Ширина дисков у водорослей разных видов колеблется от 70 до 600 Å, а иногда и более.
В отличие от высших растений в хлоропластах у водорослей выявлено большое разнообразие в расположении дисков (рис. 10). Они могут быть одиночными, как у красных водорослей, и опоясывать весь хлоропласт, ориентируясь параллельно его оболочке (поэтому на поперечных срезах выявляется система в виде длинных обособленных пластинок). У остальных водорослей, за исключением зеленых, диски собираются в пачки по 2-4 и могут лежать там порознь или тесно прилегать друг к другу. У харовых и зеленых водорослей число дисков в пачке достигает нескольких десятков (у нителлы - от 4 до 20, у хлорококка - от 2 до 40). Причем эти колебания в значительной степени зависят от возраста органеллы (по мере ее старения число дисков в пачке увеличивается), интенсивности освещения (при низкой освещенности число дисков в пачке больше, чем при высокой) и даже от качества света.
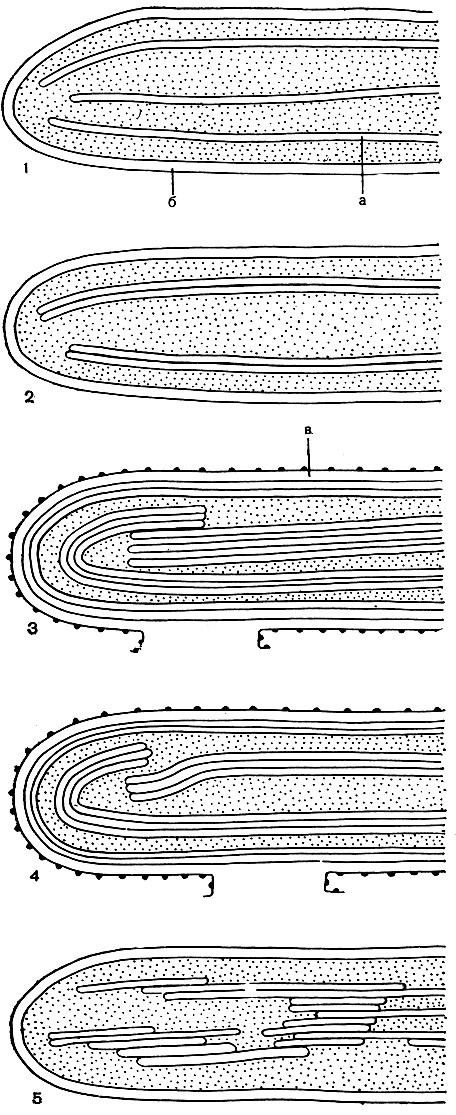
Рис. 10. Схемы расположения дисков в хлоропластах различных водорослей: 1 - хлоропласты красных водорослей; 2 - хлоропласты криптомонад; 3 - хлоропласты бурых водорослей; 4 - хлоропласты золотистых водорослей; 5 - хлоропласты зеленых водорослей; а - диски, б - оболочка хлоропласта, в - канал эндоплазматической сети, огибающий хлоропласт
Увеличение числа дисков в пачке, как правило, сопровождается их укорочением. В хлоропластах некоторых зеленых водорослей можно наблюдать, как короткие многодисковые пачки, чередуясь с единичными длинными дисками, упорядочиваются в штабеля или стопки - структуры, получившие название граноподобных образований, так как по внешнему виду они очень напоминают компоновку дисков в гранах, свойственных хлоропластам высших растений.
Диски являются теми структурами, к которым прикрепляются пигменты и в которых осуществляется фотосинтез. Не исключено, что тот или иной способ группировки дисков зависит в значительной степени от особенностей качественного состава пигментов, который, как уже говорилось, у водорослей отличается большим разнообразием.
В самое последнее время в хлоропластах были обнаружены своеобразные трубчатые элементы - микротрубочки сечением около 260 Å, которые обычно располагаются вдоль оболочки органеллы группами по 5-6 штук в каждой и, очевидно, выполняют роль каркаса.
В строме хлоропласта, помимо пластинчатых структур, выявлены более плотные зоны, представляющие скопления ДНК, многочисленные, рассеянные, мелкие (в среднем порядка 140 Å) частицы - рибосомы, глобулы различного размера, формы и состава, а также зерна крахмала, которые образуются в ходе фотосинтеза.
Обладая всем необходимым для осуществления белкового синтеза, хлоропласты относятся к числу самовоспроизводящихся органелл. Они размножаются путем перетяжки надвое и, в очень редких случаях, почкованием. Эти процессы приурочены к моменту клеточного деления и идут столь же упорядоченно, как и деление ядра, т. е. события следуют здесь в строгой последовательности одно за другим: стадия роста сменяется периодом дифференциации, за которым наступает состояние зрелости, или готовности к делению.
Хлоропласты являются единственным местом отложения крахмала в клетке, причем у водорослей часть его, и весьма значительная, концентрируется вокруг специфических образований, получивших название пиреноидов (греч. "пирен" - косточка и "эйдос" - вид, т. е. имеющий вид косточки). Пиреноиды самым тесным образом, как структурно, так и функционально, связаны с хлоропластом (рис. 9).
Среди водорослей только сине-зеленые целиком состоят из беспиреноидных форм. Пиреноиды, как правило, отсутствуют и в клетках высших растений. Исключение составляет лишь небольшая группа мхов из порядка антоцероталес.
Пиреноид представляет собой плотное образование белковой природы, окруженное снаружи обкладкой в виде сплошного кольца или отдельных пластинок в числе от двух и более, обычно крахмальной природы. Однако у бурых водорослей она образована водорастворимым полисахаридом - ламинарином; у красных - особой формой крахмала, так называемым багрянковым крахмалом; у эвгленовых - парамилоном; у хризомонад - липидами. Изредка, как у желто-зеленых, пиреноиды лишены обкладки, и поэтому их называют голыми. Обкладка с ее высоким коэффициентом преломления света делает пиреноиды очень заметной структурой клетки, и они первыми бросаются в глаза при просмотре под микроскопом живого материала.
Световой микроскоп не давал возможности проникнуть в детали строения пиреноида, и лишь благодаря электронному микроскопу удалось внести ясность в этот вопрос. Оказалось, строма пиреноида плотнее стромы хлоропласта; она пронизана либо одиночными, либо собранными в пачки дисками, которые представляют собой продолжение дисков хлоропласта, и не отделена мембранным барьером (рис. 2, 3; 9). По сути, границей между хлоропластом и пиреноидом условно служит его обкладка.
В голых же пиреноидах этот вопрос трудно решить даже на ультраструктурном уровне, поскольку единственными критериями остаются слабоуловимые различия в плотности стромы и поведении ламеллярной системы, которая при переходе в пиреноид претерпевает некоторую редукцию в числе дисков, сопровождающуюся увеличением их ширины.
Обычно пиреноиды располагаются в хлоропласте (погруженные пиреноиды). Реже они лежат вне его (выступающие пиреноиды), что особенно характерно для красных водорослей, некоторых родов эвгленовых, пирофитовых, желто-зеленых и бурых водорослей.
Чаще всего хлоропласт содержит только один пиреноид, хотя их может быть два и более, а у отдельных водорослей (спирогира, кладофора) их число даже доходит до нескольких десятков.
Форма пиреноидов отличается крайним разнообразием: от круглой и многоугольной до палочковидной.
Размеры пиреноидов колеблются в широких пределах: мелкие не превышают в диаметре 3 мкм, а крупные достигают 10-15 мкм, причем этот показатель варьирует от вида к виду, у особей внутри одного вида в зависимости от химического состава среды, интенсивности света, температуры и т. д. и даже в пределах одной клетки (наряду с одним крупным может существовать несколько мелких), на что, в частности, влияет возраст пиреноидов (молодые пиреноиды всегда мельче закончивших свой рост старых).
Новые пиреноиды могут возникать тремя различными способами: делением надвое, фрагментацией крупного родительского пиреноида на большое число мелких дочерних и в результате новообразования. Причем в первом и во втором случаях пиреноид не всегда распадается на равновеликие части, что является еще одной причиной наличия в клетке пиреноидов разного размера.
Часто в пределах одного и того же организма можно наблюдать одновременно все способы образования новых пиреноидов.
Сейчас накопилось уже много фактов, свидетельствующих о том, что пиреноид является не просто местом скопления крахмала или иных резервных веществ, а той зоной хлоропласта, в которой или при участии которой наиболее активно осуществляется их синтез. Возникнув первоначально как центр образования запасных веществ и в первую очередь как центр крахмалообразования в хлоропласте, пиреноид как бы остановился на полпути в своем поступательном движении, не успев получить того структурного воплощения, которое присуще типичным органеллам.
В дальнейшем развитие клетки пошло не по линии совершенствования этого образования, а в направлении создания качественно иной, менее громоздкой структуры с аналогичными функциями, о чем свидетельствует отсутствие пиреноидов в хлоропластах растений, занимающих более высокую ступень развития по сравнению с водорослями.
У подвижных или утративших подвижность клеток на переднем конце тела часто можно наблюдать небольшое, но очень заметное, благодаря интенсивно-красной или ярко-оранжевой окраске, пятно - стигму, или глазок (рис. 2, 1в). Такая окраска объясняется скоплением здесь большого количества каротина.
В одних случаях (у зеленых, золотистых, бурых, некоторой части желто-зеленых водорослей) стигма располагается в хлоропласте (рис. 11, 7, 2), а в других (у эвгленовых, панцирных жгутиконосцев) - за его пределами, в непосредственной близости от двигательного аппарата клетки (рис. 11, 3, 4, 5).
Вот, собственно, и все сведения, которые удалось собрать о стигме за долгую историю ее изучения с помощью светового микроскопа.
Электронный микроскоп намного расширил границы наших знаний в этой области. С его помощью не только были раскрыты тайны строения, но и обнаружены такие особенности тонкой организации, которые убедительно свидетельствуют о существовании стигм разной степени сложности.
Основу стигмы составляют плотные, разного диаметра (от 750 до 6600 Å) пигментонесущие глобулы в числе от 5-6 до нескольких десятков, которые располагаются плотными рядами (их иногда насчитывается до 9). Такой тип строения стигмы, хотя он наиболее распространен, считается самым примитивным (рис. 11, 1).
Явным признаком усложнения стигмы следует признать группировку глобул по 3-5 в комплексы, отграниченные друг от друга тонкой мембраной, что наблюдается у эвгленовых водорослей (рис. 11, 3).
Еще более сложной считается организация стигмы у некоторых панцирных жгутиконосцев, поскольку, помимо комплексов глобул, у них имеется необычного вида пластинчатое тело. Оно состоит из целой серии параллельно расположенных, сообщающихся между собой, уплощенных мешочков, которых иногда бывает до 50, причем самые крайние из них переходят в находящийся поблизости канал эндоплазматической сети. Да и каждый комплекс глобул в отдельности несколько сложнее по своей организации, чем у эвгленовых, так как состоит не из одного, а из двух рядов глобул, промежуток между которыми заполняется зернистым веществом, и вместо одиночной мембраны окружается оболочкой, образованной из двух мембран (рис. 11, 4).
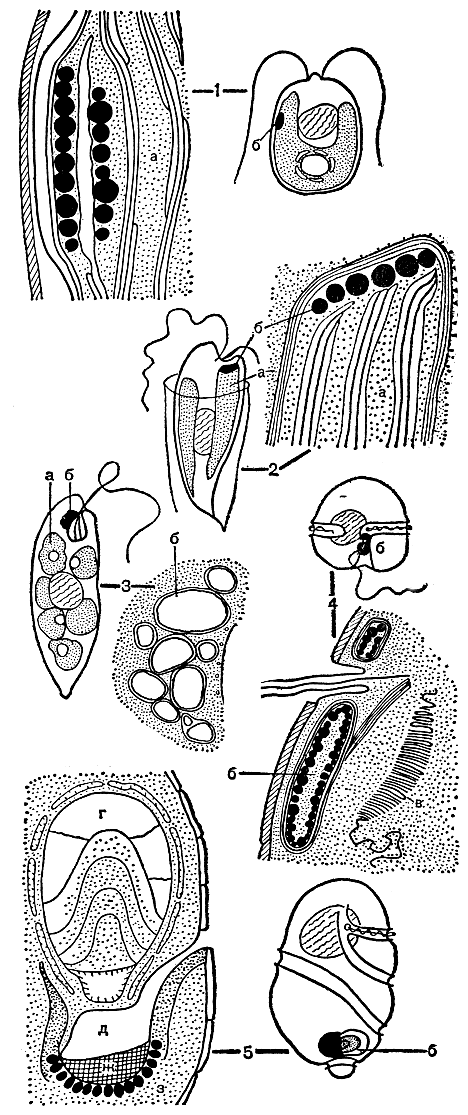
Рис. 11. Различные типы стигм, обнаруженные в клетках водорослей: 1 - Chlamydomonas; 2 - Dinobryon; 3 - Euglena; 4 - Glenodinium; 5 - Nematodium: a - хлоропласт, б - стигма, в - пластинчатое тело, г - линзовидное тело, д - полость, соединенная каналом с цитоплазмой клетки, ж - ретиноид, з - пигментные тела
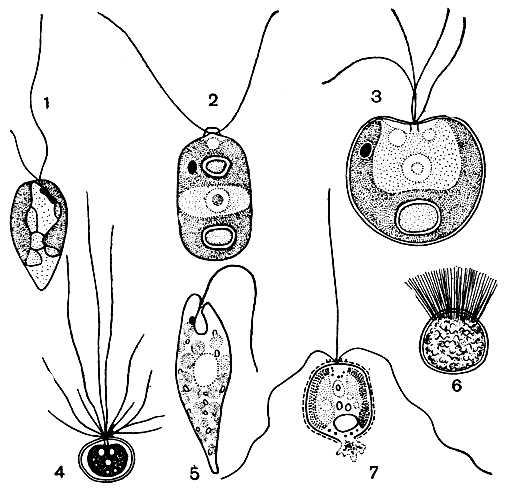
Рис. 12. Жгутиковые клетки: 1 - Ochromonas ludibunda; 2 - Chlamydomonas pertusa; 3 - Platymonas cordiformis; 4 - Schizochlamys gelatinosa; 5 - Euglena proxima; 6 - зооспора Derbesia neglecta; 7 - Chrysochromulina kappa с гаптонемой
Среди панцирных жгутиконосцев встречаются организмы и с очень сложно устроенной стигмой, представляющей многокомпонентную систему, куда входят линзовидное тело, ретиноид - тело, заполненное определенным образом ориентированнымй фибриллами, плотные глобулы. Таким образом, свет, попадая на линзу, фокусируется и затем концентрированным пучком посылается через ретиноид на глобулу, т. е. приходит туда в преобразованном виде (рис. 11, 5).
Основной функцией стигмы считается улавливание и до некоторой степени, очевидно, преобразование света, необходимые для определенной ориентации тела водоросли в пространстве. Стигму по ее роли в клетке можно уподобить локаторному устройству, принцип работы которого в данном случае, к сожалению, пока не выяснен. Поэтому сейчас еще преждевременно обсуждать вопрос о значении того или иного способа организации этой системы, ибо хорошо известно, что сложность и громоздкость конструкции не всегда являются показателем ее совершенства, рациональности и надежности в работе, тем более что выявлено много водорослей, которые лишены стигмы, но между тем прекрасно реагируют на свет.
Почти все водоросли (исключение составляют сине-зеленые и красные) могут образовывать клетки, подвижные в течение всей своей жизни или только на определенном этапе жизненного цикла (от нескольких часов до нескольких минут).
Подвижные клетки отличаются от неподвижных целым рядом особенностей организации, прежде всего появлением особого двигательного аппарата.
Перемещение в жидкой среде осуществляется при участии особых структур, представляющих собой либо временные (ризоподии), либо постоянные (реснички, псевдоцилии, жгутики) выросты клетки (рис. 12).
Ресничками называют многочисленные (от нескольких десятков до нескольких сотен) короткие (порядка 5-10 мкм) образования, совершающие энергичные колебательные движения. Псевдоцилии, или ложные реснички, имеют вид длинных, очень тонких, неподвижных волосков, которые обнаруживаются чаще всего только после подкраски. Под жгутиками понимают длинные (в несколько десятков микрометров) малочисленные образования с меньшей, чем у ресничек, частотой биений и с волнообразным характером движения, почему их иногда еще называют ундулиподиями (лат. "унда" - волна, "подиум" - возвышение).
Перечисленные образования - единственные клеточные структуры, расположенные за пределами самой клетки.
В дальнейшем речь пойдет исключительно о жгутиках, потому что они наиболее характерны для водорослей и принципиально ничем не отличаются от ресничек и псевдоцилий.
Среди водорослей чаще всего встречаются формы дву- или одножгутиковые, реже четырех-, восьмижгутиковые и совсем единичны с большим числом жгутиков, как у дербезии (рис. 12). У некоторых золотистых водорослей между двумя подвижными жгутиками располагается третий - неподвижный жгутик с расширением на конце, или гаптонема. С ее помощью клетка прикрепляется к субстрату (рис. 12, 7).
По соотношению длины жгутиков у клетки различаются водоросли равножгутиковые и разножгутиковые, или гетероконты (рис. 12, 1, 2). Иногда, например у эвглен, второй жгутик настолько редуцируется, что удается обнаружить лишь его короткий пенек. Жгутики могут быть направлены вперед по ходу движения клетки, что свойственно большинству равножгутиковых форм. У разножгутиковых нередко наблюдается разница в частоте биений (гетеродинамизм) и в положении по отношению к клетке: обычно длинный (двигательный) жгутик направлен вперед и работает энергичнее, чем короткий (рулевой), который обращен в сторону или назад по ходу движения ор-ганизма. Считается, что двигательный жгутик обеспечивает поступательное движение клетки, а рулевой - корректирует его направление.
Как правило, вся жизнь жгутиконосцев проходит в непрерывном движении. Некоторые из них развивают скорость до 500 мкм/сек и преодолевают по 5-10 м в сутки - расстояния для столь крошечных существ поистине марафонские.
Жгутики бывают гладкими или опушенными. Последние несут на своей поверхности выросты в виде тонких, диаметром не более 50 Å, волосков (мастигонем) и неминерализованных, разных по форме и размерам чешуек (рис. 13, 1, 2). Опушенным обычно оказывается двигательный жгутик. Причем у некоторых водорослей (празинокладус, колеохете) жгутики одновременно несут и мастигонемы и чешуйки, но чаще имеется какой-то один тип образований. В отличие от чешуек мастигонемы располагаются очень упорядочение, параллельными рядами. И те, и другие закладываются в цитоплазме, в пузырьках с аморфным содержимым и в них переносятся на поверхность жгутика.
Несмотря на простоту внешнего облика, строение жгутика оказалось довольно сложным, что особенно хорошо видно при изучении его в электронном микроскопе. Сопоставление картин продольных и поперечных срезов позволило воссоздать в основных чертах принципы внутреннего устройства жгутика (рис. 14). Цилиндрическое, несколько сужающееся к вершине тело жгутика окружено снаружи тонкой, порядка 90 Å, мембраной, представляющей продолжение плазма леммы, и заполнено веществом, которое по виду напоминает цитоплазму и в которое погружена система правильно ориентированных микротрубочек. Они располагаются по схеме 9 + 2, что расшифровывается следующим образом: по периферии цилиндра параллельно его поверхности размещается 9 пар тесно прилегающих друг к другу микротрубочек, а в центре его лежат две одиночные микротрубочки. Причем те и другие связаны между собой целой серией переходных структур, в расположении которых усматриваются свои закономерности, что, надо думать, имеет прямое отношение к специфической функции жгутика - осуществлять движение по определенной траектории. Выявленная в жгутиках у водорослей система расположения микротрубочек присуща подавляющему большинству организмов не только растительного, но и животного происхождения. Отклонения в сторону упрощения или усложнения отмечаются крайне редко и обычно связаны, соответственно, с ослаблением или повышением частоты биения жгутика и с особенностями его движения.
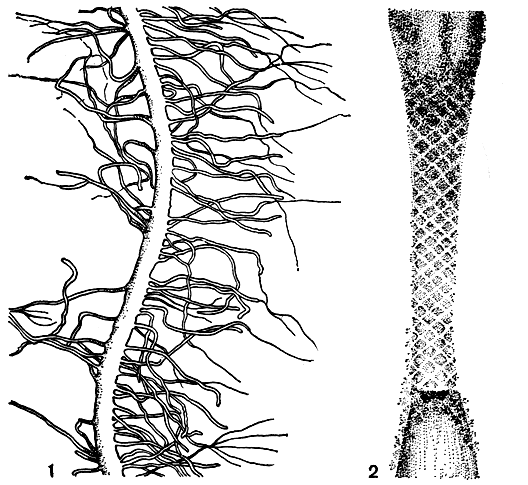
Рис. 13. Поверхностные структуры жгутиков: 1 - мастигонемы на жгутике сперматозоидов бурой водоросли Himanthalia lorea; 2 - чешуйки на жгутиках сперматозоида харовой водоросли Chara corallina
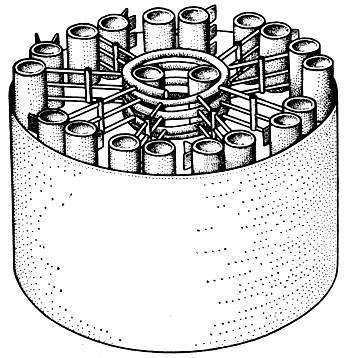
Рис. 14. Схема организации фибриллярных ультраструктур стержня реснички
При основании каждого жгутика лежит базальное тело (рис. 2, 2з). Оно имеет вид короткого, длиной около 2 мкм цилиндра, в периферической части которого располагается 9 триплетов микротрубочек, представляющих единую систему с периферическими парами микротрубочек жгутика. Обычно базальное тело одного жгутика лежит под углом 90 или 180° к базальному телу другого. Иногда базальные тела соединяются перемычкой, что обеспечивает максимальную согласованность в их действиях.
От базального тела внутрь цитоплазмы довольно часто, но не всегда отходит один или несколько так называемых жгутиковых корешков или ризопластов (рис. 2, 2k). Ризопласт представляет собой сложную систему, состоящую из двух компонентов: из одиночных или собранных в пучок (по 2-9) микротрубочек, которые направляются к плазмалемме, выполняя там роль цитоскелетных образований, и из поперечно-исчерченной, фибриллярной по своей природе структуры, направляющейся к ядру и, как у хризомонад, там закрепляющейся. Последнее указывает на связь жгутиков с этой клеточной органеллой. Таким образом, двигательный аппарат клетки представляет сложную систему, состоящую из жгутиков, базальных тел и ризопластов.
У подвижных или потерявших подвижность водорослей нередко обнаруживаются структуры, по своей внутренней организации очень напоминающие базальные тела. Они обычно располагаются около ядра (рис. 2, 2 и), а во время его деления расходятся в противоположные стороны, образуя своеобразные полюсы, или центры, деления, за что и получили название центриолей. Центриоли - непременные структуры клеток животных организмов, а среди растений встречаются только у водорослей. Как показывают ультраструктурные исследования, центриоли, с одной стороны, связаны с ядром, а с другой - с двигательным аппаратом клетки, объединяя их в своеобразный комплекс.
Интересно, что если между двигательным аппаратом клетки и ядром в отдельных случаях все же можно наблюдать отчетливую структурную связь, то между двигательным аппаратом и стигмой, вопреки ожиданиям, таковой не обнаружено, хотя взаимодействие, пусть и опосредованное, безусловно должно существовать.
В процессе филогенетического развития водорослей жгутиковый аппарат, как это случилось и с пиреноидом, постепенно редуцировался. Это сопровождалось сначала частичной, а затем и полной утратой подвижности, распадением жгутиково-ядерного комплекса и исчезновением центриолей в клетке.
В настоящее время не вызывает сомнения, что водоросли дали начало наземным растениям, проложив им дорогу на сушу. Этот акт подготавливался всем ходом развития водорослей, в котором решающая роль, по-видимому, принадлежала клетке. Большое разнообразие строения, состава и свойств отдельных клеточных компонентов свидетельствует о том, что здесь шел интенсивный формообразовательный процесс. На его основе методом проб и ошибок отбирались наиболее перспективные структуры и создавался такой тип клеточной организации, который позволил растениям перейти к наземному образу жизни.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'