Размножение и циклы развития водорослей (Ю. Е. Петров)
Воспроизведение себе подобных у водорослей совершается посредством вегетативного, бесполого и полового размножения.
Вегетативное размножение одноклеточных водорослей заключается в делении особей надвое. У многоклеточных водорослей оно происходит несколькими способами, в том числе при механическом разрушении слоевища на части (волнами, течением, в результате погрызов животными) или вследствие процессов, сопровождающихся распадением нитей на многоклеточные или одноклеточные части. Например, делению нитей сине-зеленых водорослей на части нередко предшествует отмирание отдельных клеток. Иногда для вегетативного размножения служат специальные образования. На слоевищах сфацелярии (из бурых водорослей) вырастают почки, которые опадают и прорастают в новые слоевища. Харовые водоросли образуют одноклеточные или многоклеточные клубеньки, перезимовывающие и дающие новые растения. У ряда нитчатых водорослей (например, у улотрикса из зеленых) отдельные клетки округляются, накапливают большое количество запасных питательных веществ и пигментов, одновременно происходит утолщение их оболочки. Такие клетки называют акинетами. Они способны переживать неблагоприятные условия, когда обычные вегетативные клетки погибают, что ведет к разрушению нити. Подобного типа акинеты есть у нитчатых сине-зеленых водорослей, но их подчас называют спорами. Некоторые красные, бурые, зеленые и харовые водоросли имеют стелющиеся побеги, на которых вырастают новые слоевища.
Размножение частями слоевищ не всегда приводит к возобновлению нормальных растений. Морские водоросли, растущие исключительно на твердых грунтах (камнях и скалах), нередко частично или полностью разрушаются под действием волн. Оторванные части или целые слоевища не способны снова закрепиться на твердых грунтах, так как этому мешает постоянное движение воды. Кроме того, органы прикрепления вновь не образуются. Течения относят такие слоевища в наиболее спокойные места, обычно с илистым или песчаным дном, где они продолжают расти, лежа на грунте. Со временем более старые части отмирают и отходящие от них ветви превращаются в самостоятельные слоевища. Таким способом происходит их постоянное вегетативное размножение. При этом вследствие произрастания в спокойных местах такие водоросли сильно видоизменяются: ветви их становятся тоньше, уже и слабее разветвляются. В подобных случаях говорят о неприкрепленных или свободноживущих формах соответствующих видов. Иногда они образуют крупные скопления, например неприкрепленные формы красных водорослей: филлофора в Черном море, фурцеллярия в Балтийском море, анфельция в дальневосточных морях.
Неприкрепленные формы донных водорослей никогда не образуют органов полового и бесполого размножения. Органы размножения можно наблюдать у них крайне редко - на тех обрывках или слоевищах, которые были оторваны после закладки указанных органов. В этих случаях их развитие и созревание завершаются нормально, но в последующем органы размножения уже не развиваются вновь.
Вегетативное размножение в сущности представляет собой форму бесполого размножения, осуществляемого вегетативными частями.
Бесполое размножение настоящее сопровождается, во-первых, делением протопласта клетки на части и, во-вторых, выходом продуктов деления из оболочки материнской клетки. При этом перед делением протопласта в нем происходят какие-то не вполне изученные процессы физиологической перестройки, ведущие к его омоложению. Выход продуктов деления из оболочки материнской клетки - наиболее существенное отличие настоящего бесполого размножения от вегетативного. Бывает, что в клетках образуется по одной споре, но, в отличие от акинет, они покидают оболочку материнской клетки.
Бесполое размножение водорослей происходит посредством спор или зооспор (спор со жгутиками). Они образуются или в клетках, не отличающихся по форме от других клеток, или в особых клетках, называемых спорангиями, которые нередко имеют другие размеры и форму, чем вегетативные. Главное отличие спорангиев от других клеток заключается в том, что они возникают как выросты обычных клеток и выполняют только функцию образования спор. Отличительная особенность спор и зооспор - упрощенная по сравнению с обычными клетками форма и мелкие размеры. Они бывают шаровидными, эллипсоидными или яйцевидными, покрытыми оболочкой или без нее.
Сине-зеленые водоросли, являющиеся прокариотами, имеют два рода спор - эндоспоры и экзоспоры. Эндоспоры образуются по нескольку в клетках в результате дробления содержимого. Экзоспоры возникают как вырост протопласта на вершине клетки (только у одноклеточных представителей порядка хамесифоновых); по мере его роста в длину появляются перетяжки, отчленяющие шаровидные споры.
Образованию спор и зооспор у эукариотических водорослей как в спорангиях, так и в вегетативных клетках предшествует деление ядра. При этом в зависимости от особенностей цикла развития может происходить редукция числа хромосом (мейоз). Дочерние ядра равно-мерно распределяются в цитоплазме. Одновременно делятся хлоропласты и другие органеллы, после группировки их вокруг отдельных ядер происходит деление цитоплазмы и окончательное формирование спор или зооспор. У некоторых динофитовых зооспоры образуются почкованием на поверхности материнской клетки.
У большинства эукариотических водорослей бесполое размножение осуществляется посредством зооспор. В одной клетке или спорангии их может быть от одной (эдогониум из зеленых) до нескольких сотен (кладофора из зеленых). Зооспоры могут иметь различное строение, что в известной мере отражает различия в строении одноклеточных водорослей, бывших предками соответствующих групп. Зооспоры бывают с одним, двумя, четырьмя или множеством жгутиков; в последнем случае они располагаются венчиком на конце.
У водорослей можно встретить несколько типов спор. Многие хлорококковые из зеленых и желто-зеленых имеют споры, одевающиеся оболочкой внутри материнской клетки. Такие споры называют апланоспорами. При образовании особо утолщенной оболочки их именуют гипноспорами, так как они способны длительный срок находиться в состоянии покоя. Гипноспоры формируются по одной в клетке, но, в отличие от акинет, оболочка материнской клетки не участвует в образовании их оболочки. Иногда апланоспоры сразу в материнской клетке приобретают форму, подобную ей. В таких случаях говорят об автоспорах. Существуют еще споры, в названии которых отражается число их в спорангии: тетраспоры - образуются по 4 (многие красные и диктиотовые из бурых), биспоры - две споры в спорангии (некоторые кораллиновые из красных), моноспоры - одна спора в спорангии (некоторые красные).
Споры и зооспоры обычно выходят в воду через отверстие в стенке спорангия целой группой, окруженные слизистой оболочкой, которая вскоре расплывается. Зооспоры по выходе из спорангия, находясь еще в общей оболочке, начинают активно двигаться и после разрыва оболочки моментально расплываются в разные стороны. Подобным образом выходят и гаметы при половом размножении.
Половое размножение заключается в слиянии двух клеток (гамет), в результате чего образуется зигота, вырастающая в новую особь или дающая зооспоры. У водорослей половое размножение бывает нескольких типов. В простейшем виде оно представляет соединение содержимого двух вегетативных клеток. У одноклеточных жгутиковых водорослей (некоторые вольвоксовые) половой процесс сводится к слиянию двух особей и называют его гологамией. При слиянии содержимого двух безжгутиковых вегетативных клеток половой процесс называют конъюгацией (рис. 24). Это единственная форма полового размножения в классе конъюгат из зеленых водорослей. Гораздо чаще половое размножение у водорослей, в том числе у одноклеточных жгутиковых, связано с дроблением содержимого клеток и образованием внутри их специализированных половых клеток - гамет. У всех водорослей, кроме конъюгат и красных водорослей, по крайней мере мужские гаметы имеют жгутики, а у гамет противоположного пола они имеются не всегда. Образуются гаметы так же, как споры и зооспоры. Особые вместилища гамет называют гаметангиями. Число гамет в клетке или гаметангии может колебаться от одной до нескольких сотен. У примитивных водорослей гаметы формируются в вегетативных клетках.
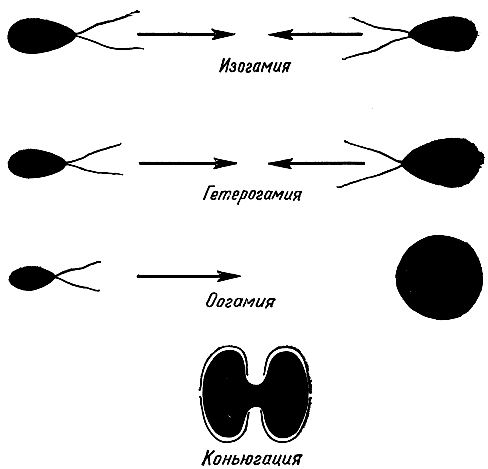
Рис. 24. Формы полового процесса у водорослей
В зависимости от относительных размеров гамет, участвующих в слиянии, различают следующие типы полового процесса (рис. 24):
1) изогамия - гаметы одинаковой величины и формы;
2) гетерогамия, или анизогамия,- одна гамета (женская) крупнее другой (мужской), но сходна с ней;
3) оогамия - женская гамета, называемая яйцеклеткой, лишена жгутиков, неподвижна и значительно крупнее мужской, которую называют сперматозоидом или антерозоидом, она может быть бесцветной; гаметангии с яйцеклетками именуются оогониями, а гаметангии с мужскими гаметами - сперматангиями или антеридиями;
4) автогамия - особый тип полового процесса, распространенный у части диатомовых. Заключается он в том, что ядро клетки предварительно делится с мейозом на 4 ядра, два из них разрушаются, и оставшиеся два ядра сливаются, образуя вновь диплоидное ядро. Автогамия не сопровождается увеличением числа особей, а лишь их омоложением.
При гетеро- и оогамии мужские и женские гаметы могут развиваться на одной особи или колонии (обоеполые, или однодомные, виды) или на разных (раздельнополые, или двудомные, виды). Среди водорослей, для которых характерна изогамия, существуют гомоталличные виды (у них сливаются гаметы из одного слоевища или колонии) и гетероталличные (слияние возможно только между гаметами из разных особей), которые ввиду отсутствия морфологических различий обозначаются знаками + и -, соответственно различают + гаметы и - гаметы.
В результате слияния гамет образуется шаровидная зигота, при этом жгутики отпадают и появляется оболочка. Зиготы некоторых водорослей какое-то время сохраняют жгутики, тогда получается планозигота, которая способна плавать от нескольких дней до трех недель. В зиготе происходит слияние двух ядер гамет, и она становится диплоидной. В дальнейшем зиготы разных водорослей ведут себя различно. Одни зиготы вырабатывают толстую оболочку (гипнозиготы) и впадают в период покоя, длящийся до нескольких месяцев. Другие зиготы прорастают без периода покоя. В одних случаях из зигот непосредственно вырастают новые слоевища. В других зиготы делятся с мейозом и образованием зооспор; такие зиготы предварительно растут, и из них в зависимости от размеров выходят 4-32 зооспоры.
Среди водорослей наблюдаются случаи партеногенетического (без оплодотворения) развития женских гамет. Внешне они подобны обычным зиготам, и их называют азиготами или партеноспорами.
У одного и того же вида водорослей в зависимости от времени года и внешних условий наблюдаются разные формы размножения (бесполое и половое), при этом происходит смена ядерных фаз (гаплоидной и диплоидной). Исключение составляют виды, лишенные полового процесса. Изменения, претерпеваемые особями вида между одноименными стадиями (моментами жизни), составляют его цикл развития.
У одних видов органы бесполого и полового размножения развиваются на разных особях; тогда растения, образующие споры, называют спорофитами, а растения, производящие гаметы,- гаметофитами. У других водорослей споры и гаметы образуются на одних и тех же растениях; одновременно у таких видов могут существовать и особи, дающие только споры, т. е. спорофиты (порфира). Сейчас растения, способные производить и споры (зооспоры) и гаметы, как правило, называют гаметофитами. Однако во избежание путаницы с настоящими гаметофитами, производящими только гаметы, их лучше называть гаметоспорофитами.
Развитие органов размножения того или иного типа у гаметоспорофитов определяется температурой. Например, пластинчатые слоевища одного из видов порфиры (Porphyra tenera) при температуре ниже +15, +17 °С производят органы полового размножения, а при более высокой температуре - органы бесполого размножения. И у других водорослей гаметы обычно появляются при более низкой температуре, чем споры. При промежуточной температуре развитие тех или иных органов размножения на гаметоспорофитах определяется другими факторами - интенсивностью света, длиной дня, сезонными изменениями химического состава воды или солености (для морских водорослей). Гаметоспорофиты существуют у улотриксовых, ульвовых и кладофоровых из зеленых водорослей, у эктокарповых, хордариевых, сфацеляриевых и пунктариевых из бурых водорослей, бангиевых и некоторых немалиевых из красных.
Спорофиты и гаметофиты (гаметоспорофиты) бывают одинакового строения или разного, и соответственно существуют понятия изоморфной (сходной) и гетероморфной (разной) смены форм развития (чередования поколений). В отношении большинства водорослей неправильно говорить о чередовании поколений спорофитов и гаметофитов (гаметоспорофитов), так как они нередко существуют одновременно. Иногда они могут расти в несколько различных экологических условиях. Например, спорофит порфиры имеет вид ветвящихся нитей из одного ряда клеток, которые внедряются в известковый субстрат (раковины моллюсков, известковые скалы) и предпочитают слабое освещение, проникая на большую глубину. Гаметоспорофит порфиры пластинчатый и растет вблизи уреза воды, в том числе в приливно-отливной зоне.
Разница в строении спорофитов и гаметофитов (гаметоспорофитов) при гетероморфной смене форм развития может быть очень значительной. Гаметоспорофит или гаметофит может быть многоклеточным, высотой в несколько сантиметров, а спорофит - микроскопическим, одноклеточным (акросифония из зеленых). Возможна и обратная картина, когда гаметофит микроскопический и даже одноклеточный, а спорофит достигает в длину 12 м (ламинария японская из бурых). Гаметофиты и спорофиты подавляющего большинства водорослей - самостоятельные растения. У ряда водорослей спорофиты растут на гаметофитах (филлофора Броди из красных) или гаметофиты развиваются внутри слоевищ спорофитов (циклоспоровые из бурых).
Поскольку при половом процессе в результате слияния гамет и их ядер происходит удвоение набора хромосом в ядре, то в последующем в какой-то момент цикла развития наступает редукционное деление ядра (мейоз), в результате которого дочерние ядра получают одинарный набор хромосом. Спорофиты многих водорослей диплоидные, и мейоз в цикле их развития совпадает с моментом образования спор, из которых развиваются гаплоидные гаметоспорофиты или гаметофиты. Такой мейоз называют спорической редукцией (рис. 25,1).
В спорофитах примитивных водорослей (кладофора, эктокарпус и многие другие) наряду с гаплоидными спорами могут образовываться диплоидные споры, которые снова развиваются в спорофиты. Споры, появляющиеся на гаметоспорофитах, служат для воспроизведения материнских растений. Спорофиты и гаметофиты водорослей, стоящих на верхних ступенях эволюции, строго чередуются без самовозобновления (ламинариевые из бурых, многие флоридеи из красных).
Ряд водорослей имеют мейоз в зиготе, т. е. зиготическую редукцию (рис. 25, 2). Она характерна для конъюгат из зеленых водорослей.
Зиготы части пресноводных зеленых водорослей, таких, как вольвоксовые, улотриксовые и др., представляют собой одноклеточные спорофиты. Они производят до 32 зооспор, что по массе во много раз больше, чем пара родительских гамет. Таким образом, у этих водорослей, по существу, наблюдается спорическая редукция.
Некоторые группы водорослей имеют гаметическую редукцию, которая характерна для царства животных. Мейоз происходит при образовании гамет, остальные клетки всегда диплоидные (рис. 25, 3). Такая смена ядерных фаз присуща диатомовым и циклоспоровым водорослям, а также одному из видов кладофоры (Cladophora glomerata). Интересно отметить, что диатомовые преобладают по количеству видов над другими водорослями и встречаются во всех местообитаниях, где только способны расти водоросли. В свою очередь, циклоспоровые относятся к наиболее массовым морским водорослям. Видимо, цикл развития с гаметической редукцией дает этим водорослям какие-то преимущества.
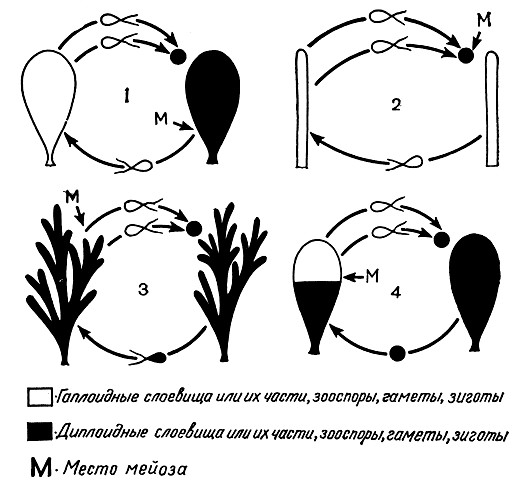
Рис. 25. Смена ядерных фаз у водорослей: 1 - спорическая редукция (ульва); 2 - зиготическая редукция (сфероплея, лишние ядра редуцируются); 3 - гаметическая редукция (кладофора собранная); 4 - соматическая редукция (празиола стебельчатая)
У зеленой водоросли празиолы (Prasiola stipitata) обнаружена соматическая редукция - мейоз совершается в вегетативных клетках верхней части диплоидного гаметофита, при этом появляются участки гаплоидных клеток, в которых вслед за тем образуются гаплоидные гаметы (рис. 25, 4).
В цикле развития водорослей, лишенных полового размножения (сине-зеленые, криптофитовые и эвгленовые) или имеющих его в редких случаях (золотистые, желто-зеленые и динофитовые), наблюдаются только изменения в строении тела. Поэтому применительно к таким водорослям принято говорить о цикломорфозе. Он может охватывать несколько поколений или ограничиваться периодом роста и развития одной особи. В наиболее резкой форме цикломорфоз выражен у гиеллы дернистой (Hyella caespitosa) из сине-зеленых водорослей и у гленодиниума Борге (Glenodinium borgei) из динофитовых.
Как циклы развития, так и цикломорфозы у водорослей отличаются большой пластичностью. Их прохождение во многом определяется экологическими условиями. Поэтому они далеко не всегда сопровождаются строго последовательным проявлением всех стадий. В зависимости от условий произрастания отдельные стадии и формы развития могут выпадать полностью (например, спорофит или гаметоспорофит и гаметофит) или, наоборот, существовать на протяжении нескольких поколений, с тем чтобы на период жизни одного поколения уступить место другой форме развития. Строго упорядоченные циклы развития существуют у водорослей, стоящих на верхних ступенях эволюции (рис. 26).
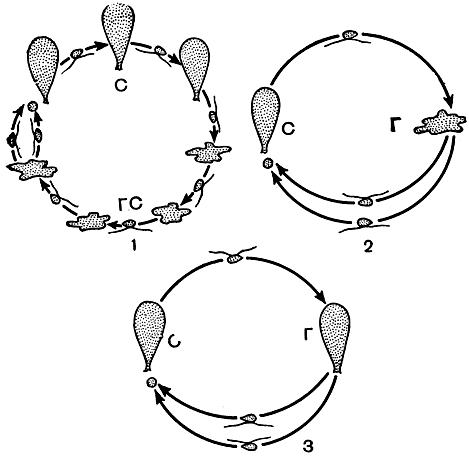
Рис. 26. Циклы развития водорослей: 1 - гетероморфный цикл с нерегулярной сменой форм развития; 2 - гетероморфный цикл с регулярной сменой форм развития; 3 - изоморфный цикл с регулярной сменой форм развития. С - спорофиты, Г - гаметофиты, ГС - гаметоспорофиты
Зачатки водорослей в виде спор, гамет и зигот не совсем стихийно разносятся водой. Они обладают различного рода таксисами, определяющими направление их движения в зависимости от внешних раздражителей: света (фототаксис), температуры (термотаксис), химических веществ, содержащихся в воде (хемотаксис). Способностью двигаться обладают не только зооспоры, но и споры, лишенные жгутиков. У них наблюдается амебоидное движение, при котором сначала образуется выступ, а затем в него перемещается содержимое всей споры.
Каждый род таксиса может быть положительным или отрицательным. При положительном таксисе зачатки водорослей двигаются в сторону усиления действующего фактора; при отрицательном таксисе - в обратном направлении. Характер таксиса определяется интенсивностью фактора и физиологическим состоянием двигающихся клеток. Слишком сильное освещение вызывает изменение положительного фототаксиса на отрицательный. Фототаксис зооспор бентосных (донных) водорослей, первоначально положительный, со временем сменяется на отрицательный, что обеспечивает оседание их на дно. Отрицательный фототаксис наблюдается также у зигот донных водорослей. Мужские гаметы обладают хемотаксисом, позволяющим им двигаться в сторону неоплодотворенных женских гамет, выделяющих особые химические вещества. Обнаружено, что споры некоторых бентосных морских водорослей, видимо, путем изменения объема тела и, таким образом, удельного веса концентрируются в слоях воды с определенной температурой и соленостью. В зависимости от направления течения в этих слоях споры выносятся к определенным участкам побережья, где и происходит развитие слоевищ.
Течения служат основным средством переноса зачатков на большие расстояния. Зооспоры сохраняют жизнеспособность в течение нескольких дней. Более длительное перемещение зачатков водорослей происходит плодоносящими слоевищами или их частями, которые сохраняются в живом состоянии до конца периода вегетации.
Наличие жгутиков у зооспор и некоторых гамет обеспечивает их перемещение в пределах лишь нескольких метров или десятков метров. Скорость движения зооспор и гамет можно сравнить со скоростью одноклеточных организмов со жгутиками - она не превышает 250 мкм/сек, или 0,9 м/ч. Такая небольшая скорость имеет значение для выбора наиболее подходящих слоев воды и места для непосредственного прикрепления на дне. На закрепление спор бентосных водорослей оказывает влияние присутствие других организмов и особей или спор данного вида и их число на единицу площади.
Для прорастания спор и зигот водорослей требуется комплекс условий, включающий определенные значения температуры, освещенности, содержания биогенных и биологически активных веществ. В противном случае они не прорастают. При этом зиготы некоторых водорослей, например фукусовых, не принадлежащие к гипнозиготам, сохраняют жизнеспособность в течение трех-четырех месяцев. Воспроизведению и сохранению некоторых водорослей в неблагоприятных условиях способствует образование цист. Они известны у золотистых, желто-зеленых, диатомовых и динофитовых водорослей. В каждой клетке формируется одна циста. Содержимое клетки округляется, и вокруг него вырабатывается твердая оболочка, содержащая кремнезем. При прорастании цист образуется одна особь, реже несколько.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'