Геотропизм
После того как мы рассмотрели многообразие влияний света на растения, обратимся теперь к тому источнику раздражений, который мы не можем ни увидеть, ни услышать, ни понюхать, ни попробовать на вкус, ни потрогать, но все-таки чувствуем. Сила его воздействия на Земле повсюду в общем одинакова, поэтому он, как правило, не особенно приметен. Но наряду со светом он представляет собой основной фактор, определяющий положение растений в пространстве. Конечно, вы уже догадались, что речь идет об ускорении силы тяжести (g=981,56 см/с2). Большое значение силы тяжести для организмов было выявлено особенно благодаря колоссальному прогрессу в области космических полетов и исследования космоса. Как раз поведение живых существ, не подвергающихся действию силы тяжести (невесомость), позволяет судить об эффективности этого фактора, проявление которого не может быть произвольно исключено на Земле.
Присущую всем растениям способность воспринимать земное притяжение и реагировать на него называют геотропизмом. Таким образом, геотропизм представляет собой ответную реакцию на ускорение силы тяжести. И если фототропизм известен уже с глубокой древности, то на искривления органов растений, обусловленные геотропизмом, внимание впервые было обращено только в XVIII веке. В 1837 г. французский ученый-биолог Дютроше (1776-1847) писал: "Глубокая тайна заключена в том, что стебли растут вверх, а корни - в противоположном направлении".
Особенно бросающийся в глаза пример геотропной ответной реакции - это направление роста деревьев, развивающихся на горном склоне, на которое вы, вероятно, уже обратили внимание во время пребывания в горах. Деревья расположены вовсе не перпендикулярно к тому конкретному участку поверхности Земли, на котором растут, а совершенно независимо от крутизны склона - в направлении, от центра Земли (рис. 24). При положительной геотропной реакции органов или их частей, например, главных корней высших растений, ризоидов водорослей, печеночных мхов и заростков папоротников, обнаруживается рост в направлении к центру Земли. А отрицательно геотропные органы или их части, напротив, растут в направлении от центра Земли. Таковы прежде всего оси главных побегов, но также ножки плодовых тел шляпочных грибов и спорангиеносцы видов Мисоr (наземных плесневых грибов) и другие объекты. Уже при прорастании семени, которое происходит в темноте, то есть вне влияния света, и независимо от положения самого семени, наблюдается изгиб зародышевого корешка вниз, а проросткового побега - вверх. Известны также случаи возвращения в вертикальное положение надломленных или полегших растений, обусловленного геотропизмом. Несомненно, вы уже замечали, что прибитые к земле бурей или дождем хлебные злаки могут снова подниматься.
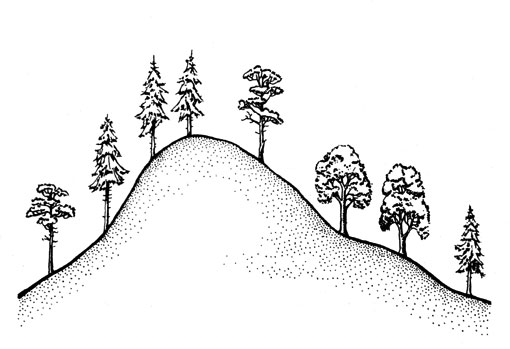
Рис. 24. Направление роста стволов деревьев на горных склонах, обусловленное отрицательным геотропизмом
Плагиотропизмом обладают прежде всего боковые корни 1-го порядка, боковые ветви и листья, тогда как корневища (подземные побеги) многих растений обнаруживают поперечный геотропизм. Если придать ненормальное положение дорзивентральным органам, таким, как листья или цветки, то они могут вернуться в изначальное положение благодаря изгибанию (геотропному повороту) черешков или цветоножек. А боковые корни 2-го порядка и боковые ветви плакучих форм деревьев вообще не обладают геотропизмом. Достаточно вспомнить вавилонскую иву (Salix babylonica).
Возможны также и изменения геотропизма. Известный тому пример - снотворный мак (Рараver somniferum); из подсохшего млечного сока, собираемого с его несозревших плодов-коробочек, получают опиум. Верхняя часть стебля, несущего бутон, бывает сначала положительно, а при распускании цветка становится отрицательно геотропной. Многие растения, как, например, относящийся к семейству бобовых земляной орех (Arachis hypogaea), который родом из Южной Америки, имеют отрицательно геотропные цветоножки, но, став плодоножками, последние становятся положительно геотропными. Противоположным образом ведет себя лилия-саранка (Lilium martagon). Если у хвойного дерева удалить отрицательно геотропный верхушечный побег, то плагиотропные боковые ветви отгибаются вверх и становятся отрицательно геотропными. Однако при этом, как правило, лишь одна из боковых ветвей замещает верхушечный побег и занимает его место, а остальные снова возвращаются в исходное положение. Изменения температуры и освещения тоже могут приводить к изменению геотропной ответной реакции. Так, отрицательно геотропные побеги некоторых сорных растений наших полей под влиянием холодов становятся поперечно-геотропными. Примерами могут служить полевая горчица (Sinapis arvensis) и красная яснотка (Lamium purpureum).
Вероятно, здесь речь идет об экологической приспособленности, поскольку благодаря изменению геотропной реакции эти сорняки нередко оказываются под защитой снежного покрова. Поперечно-геотропные корневища адоксы мускусной (Adоха moschatellina) и других растений, если окажутся освещенными, то становятся положительно геотропными. Напротив, затемнение поперечно-геотропных побегов малого барвинка (Vinсa minor), а также и других трав приводит к тому, что побеги обнаруживают отрицательный геотропизм. Плагиотропное положение боковых ветвей и листьев обусловлено более сложно, чем это кажется на первый взгляд, поскольку оно представляет собой равнодействующую двух противодействующих сил. Во-первых, здесь речь идет об отрицательном геотропизме, который стимулирует рост нижней стороны органа, а во-вторых, о тенденции к усилению роста верхней стороны, то есть о тенденции, которую называют эпинастией. С помощью вращения растения, о котором еще будет рассказано подробнее, действие тяжести может быть устранено. Листья тогда располагаются вдоль оси побега (см. рис. 26, в и г). В противоположность листьям, чья эпинастия автономна, у боковых ветвей она индуцируется длительным воздействием силы тяжести. Удаление верхушки главной оси побега может устранить эпинастию и привести к тому, что боковая ветвь станет расти вверх. Но бывают и такие боковые ветки, которые после удаления верхушки главной оси остаются плагиотропными, что можно наблюдать, например, у видов Araucaria - хвойных деревьев, растущих в Южном полушарии. Может быть, вам знакомо известное декоративное растение "норфолкская, или комнатная ель" (Araucaria excelsa).
Геотропный изгиб, как и фототропный, обычно представляет собой обусловленное ростом движение органа, которое проявляется в результате неодинакового роста его противоположных сторон. Однако иногда и уже развившиеся части растения бывают способны к геотропным изменениям положения в пространстве. К примеру, близ узлов стеблей злаков (соломин) рост уже завершился. Но если эти стебли оказались в горизонтальном положении, то рост возобновится, особенно на сторонах, ставших нижними. Благодаря этому и происходит уже упоминавшееся возвращение в вертикальное положение полегших зерновых культур. Очевидно, сила тяжести действует при этом и как ориентирующий, и как индуцирующий фактор. Деревья и ветви, закончившие рост в длину, также в состоянии осуществлять геотропные ответные реакции в результате вторичного роста в толщину, то есть образования специальных клеток. В процессе этого медленного роста возникает так называемая крень. У хвойных пород она обнаруживается в виде креневой древесины на нижней стороне, а у лиственных - на верхней в виде тяговой древесины. Образование креневой древесины в результате вспучивания, а тяговой древесины вследствие сокращения приводит к искривлению. При плагиотропных изменениях положения листьев с вздутыми сочленениями, имеющимися у некоторых растений (например, бобовых и мальвовых), осуществляются и тургорные движения.
Если процесс изгибания корней довольно прост из-за того, что зона их растяжения коротка, то у осей побегов это происходит значительно сложнее. Один из лучших объектов для демонстрации отрицательно геотропных изгибов стеблей - это колеус, или "декоративная крапива" (Coleus blumei), одно из излюбленных комнатных растений. Этот хорошо растущий и легко отвечающий на воздействия внешних факторов представитель семейства губоцветных представляет собой гибрид, возникший в результате скрещиваний тропических видов рода Соlеus. Это легко размножаемое и легко культивируемое растение уже много десятилетий используется для проведения опытов с учебными целями. Срезанные молодые растения могут сохраняться даже при выращивании их в воде, взятой из водопровода. Для приведения растений в горизонтальное положение особенно удобно применять четырехгранные вазоны. Проведенный нами опыт продолжался более 112 суток. Для постановки такого опыта наиболее пригодны помещения, где растение освещено со всех сторон, например теплицы. В помещениях же с односторонним освещением на геотропную ответную реакцию накладывается фототропная. У горизонтально расположенного растения уже через час или два можно обнаружить небольшой, но заметный изгиб. Он начинается с верхушки и распространяется в направлении к основанию. Примерно уже через сутки все листья изменяют свое положение. Характерно также прилегание нижних листьев к стеблю. А приблизительно через 30 суток после того, как растение было расположено горизонтально, вся олиственная часть стебля становится вертикальной. Дальше стебель не изгибается, поскольку здесь происходит одревеснение, ибо геотропные изгибы могут совершать, как правило, только ткани, способные расти.
На 50-й день опыта мы поставили вазон с растением донышком вниз, а днем позже олиственная часть побега снова оказалась изогнутой. При этом особенно хорошо было видно, что стебель сначала изгибался более чем на 90°, и лишь постепенно происходил обратный изгиб, приводивший его в строго вертикальное положение. Такие проявления колебательных движений некоторые исследователи называют автотропизмом. Через 50 или 100 суток после начала опыта мы повторно располагали растение горизонтально, и его стебель снова и таким же образом проявлял отрицательный геотропизм. Из-за увеличения собственного веса растения второй вертикальный отрезок его стебля довольно заметно отклонился от положения, в котором оказался первый отрезок. Во время опыта, продолжавшегося более 112 суток, произошли не только изгибания стебля вследствие усиления роста на его нижней стороне, но и другие процессы. Примечательны деление, растяжение и дифференцировка клеток, связанные с удлинением стебля и новообразованием листьев. Следует упомянуть также образовавшиеся в пазухах листьев боковые побеги, которые в нашем опыте частично были удалены. Наконец, следует отметить и опадание старых листьев.
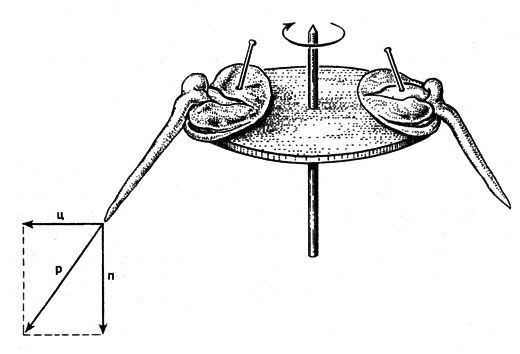
Рис. 25. Изгиб зародышевого корешка у Viсia sativa при центрифугировании. Направление, в котором растет корешок, совпадает с направлением равнодействующей (р) силы земного притяжения (п) и центробежной силы (ц)
В отличие от освещения, которое легко варьировать, а поэтому относительно просто исследовать последствия его влияния, сила тяжести повсюду на поверхности Земли проявляется непрерывно и почти одинаково. Поэтому исследования воздействий этого фактора более трудны. Эти воздействия можно обнаружить, особенно при использовании двух специфических методов: во-первых, с помощью влияния центробежной силы и, во-вторых, при применении клиностатов. В 1806 г. англичанину Найту (1759-1838) удалось продемонстрировать наличие влияния силы тяжести благодаря тому, что он закрепил семена фасоли по периферии вертикально расположенного вращающегося мельничного колеса, приводившегося в движение водой. При этом под действием центробежной силы корни росли наружу, а побеги - внутрь. Если центробежная сила (ц) и сила земного притяжения (п) примерно одинаковы, то проявляется закон равнодействующих сил (рис. 25). Это доказывает, что геотропизм представляет собой ответную реакцию на ускорение движения массы. Варьируя скорость вращения центри-фуги, можно судить об интенсивности раздражения. С помощью клиностата тоже можно наглядно продемонстрировать влияние силы тяжести (рис. 26).
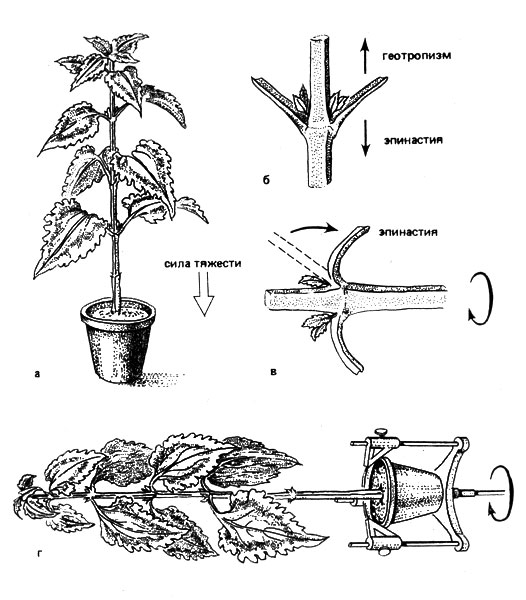
Рис. 26. Геотропизм и эпинастия у Coleus blumei. При нормальном положении растения геотропизм и эпинастия действуют в противоположных направлениях (а и б). У горизонтально расположенного растения, медленно вращающегося в клиностате вдоль своей продольной оси, изгиба не происходит (в); эпинастия листьев больше не уравновешивается отрицательным геотропизмом (г)
Идея создания такого прибора, позволяющего исключить одностороннее воздействие силы тяжести, принадлежит английскому ботанику Хантеру (1728-1793) и немецкому ботанику Саксу (1832-1897). Если клиностат включен, то горизонтально расположенное растение поворачивается вокруг своей продольной оси, и все стороны стебля и главного корня примерно одинаково и симметрично подвергаются влиянию земного притяжения. Вследствие этого растение находится в "выровненном силовом поле" и не происходит изгибания ни стебля, ни главного корня. Эпинастия тоже может быть показана с помощью клиностата. После того как одностороннее действие силы тяжести оказывается устраненным, заметно усиливается рост верхней стороны листа, и листья располагаются вдоль стебля. Скорость вращения клиностата обычно составляет несколько оборотов в минуту. При истинной невесомости в космических кораблях растения ведут себя так же, как на клиностате. Это подтверждает, что такого рода приборами можно пользоваться для исследования влияния силы тяжести. Как время действия раздражителя, так и время проявления ответной реакции может быть относительно коротким. Так, первое составляет, например, у пастушьей сумки (Capsella bursa-pastoris) лишь 2 мин, а у подсолнечника (Неlinthus) только 3 мин. Время проявления ответной реакции у колеоптилей овса равно 14, а корней обыкновенных бобов (Viсiо fabа) - примерно 85 мин. Если раздражение продолжительно, порог раздражения проявляется при ускорении около 10-2 г, то есть при одном проценте ускорения земного притяжения. Некоторые растения могут воспринимать отклонение от вертикали величиной около 1°. Поскольку растения способны суммировать подпороговые раздражения, ими воспринимаются и очень кратковременные изменения положения в пространстве. Однако заметные ответные реакции проявляются прежде всего при длительном действии силы тяжести. Этим объясняется медлительность проявления ответных реакций, избавляющая растения от непрерывного образования изгибов.
Как при фототропизме, так в определенных пределах и при геотропизме безразлично, действует ли сильный раздражитель в течение короткого времени или слабый, но соответственно дольше. Решающее значение имеет количество раздражения. Эта пропорциональная взаимозависимость количества раздражения и величины ответной реакции свидетельствует о применимости закона количества раздражения к процессам, обусловленным геотропизмом.
Существенное значение для интенсивности действия силы тяжести на какой-либо орган имеет его положение в данный момент. Уже Саксу было известно, что главный корень, обращенный вниз, то есть занимающий нормальное положение, не испытывает раздражения, но находясь в горизонтальном положении, он подвергается максимальному раздражению. Как показали обстоятельные исследования, интенсивность воздействия зависит от положения органа или от синуса угла его отклонения от нормального положения. Если разложить g на направленную параллельно оси органа продольную силу l и перпендикулярно направленную к ней компоненту к (рис. 27), то получится следующее уравнение: k = g*sin α.
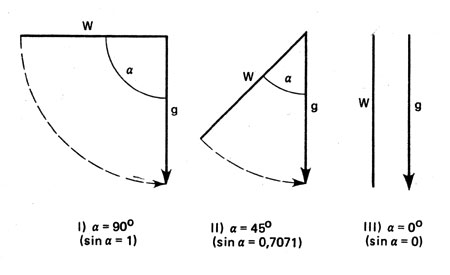
Рис. 27. Схематическое изображение проявления закона синуса: влияние силы тяжести (g) на корни (W), находящиеся в разных положениях. Действующая компонента силы тяжести примерно равна g*sin α
Как свидетельствует этот закон - закон синуса, с горизонтальным положением связано наиболее сильное проявление геотропной реакции, поскольку sin 90° =1 и k = g. Напротив, у вертикально расположенного корня угол отклонения от отвесной линии равен 0°, a sin 0°=0. Отсюда следует: к = 0.
Для восприятия света растения имеют пигменты. А как происходит восприятие раздражения, обусловливающего геотропизм? Сначала следует отметить, что восприятие раздражения ограничено определенными участками органов растения. У корней это калиптра (корневой чехлик). Подопытными объектами часто служат злаки, например ячмень (Hordeum) или кукуруза (Zеа), у которых корневой чехлик легко отделяется. В результате его удаления рост корня в длину едва замедляется или даже усиливается, но корень утрачивает способность осуществлять геотропные ответные реакции. Только после регенерации корневого чехлика возможность их осуществления восстанавливается. У колеоптилей область проявления ответных реакций ограничена самой верхушкой (участок длиной около 3 мм). Наконец, следует еще упомянуть о стеблях, у которых к геотропным реакциям способны, вероятно, лишь удлиняющиеся участки всех продолжающих рост растений.
Восприятие силы тяжести может происходить только в результате перемещения имеющих вес частиц. Согласно теории статолитов, восприятие возбуждения осуществляется при помощи статолитов, которые, следовательно, выполняют у растений функцию, подобную их функции у животных. Такие представления были высказаны еще в 1892 г. Ноллем (1858-1908). Чешский ботаник Немец (1873-1966) и немецкий физиолог Габерландт (1854-1945) независимо друг от друга в 1900 г. указали на лейкопласты или на образующиеся в них крахмальные зерна как на структуры, воспринимающие силу тяжести. Такие крахмальные зерна встречаются в клетках корневого чехлика (рис. 28) и в эндодерме - сохраняющем крахмал внутреннем слое клеток первичной коры стеблей многих растений. Если из лейкопластов устранить крахмал, подвергнув растение затемнению или охлаждению, то способность к геотропным ответным реакциям утрачивается. Следовательно, крахмал лейкопластов может играть решающую роль при выполнении ими функции статолитов. Необходимой предпосылкой для понимания осуществляемого статолитами восприятия силы тяжести служит представление о микроскопическом и субмикроскопическом строении растительной клетки, на которое в связи с этим мы здесь снова обратим внимание читателя.
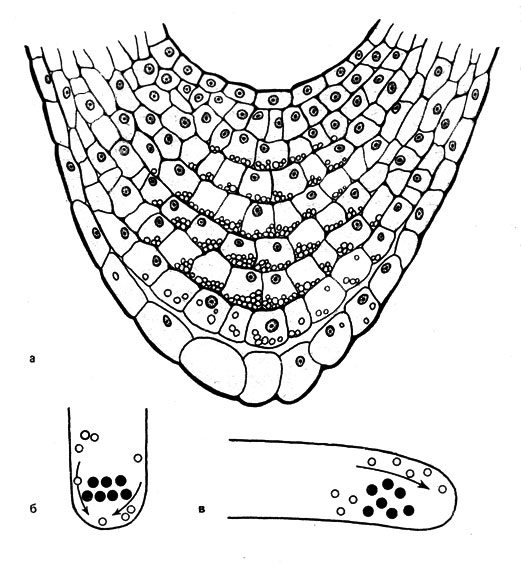
Рис. 28. Действие крахмальных зерен (а) и блестящих телец (б и в) в качестве статолитов, а - размещение статолитного крахмала в клетках корневого чехлика Rorippa amphibia (нормальное положение). И в тех положениях кончика корня, при которых действует раздражение, приводящее к проявлению геотропизма, крахмальные зерна находятся у той стороны каждой из клеток, которая оказывается в это время физически нижней, б и в - схематическое изображение кончиков ризоидов Charа, находящихся в вертикальном (б) и горизонтальном (в) положениях. Статолиты обозначены черными кружками, а направления движений пузырьков Гольджи (мелкие окружности), причастных к образованию клеточной оболочки, показаны стрелками
В настоящее время обсуждаются три возможных варианта действия статолитов. Во-первых, они могут оказывать воздействие вследствие ассиметричного расположения и, во-вторых, в результате продольного скольжения во время перемещений. В первом случае исследователи говорят о топографическом, а во втором - о кинетическом эффекте. Третий возможный вариант предполагает осуществление давления на плазматические структуры, которые и вызывают возбуждение. Этот эффект деформации, по-видимому, должен быть решающим прежде всего у высших растений. Очевидно, при этом в тех местах, где статолиты были до начала возбуждения, должно происходить прекращение давления.
С помощью электронно-микроскопических исследований можно показать, что клетки, находящиеся в области корневого чехлика, имеют весьма примечательную многослойную эндоплазматическую сеть (ЭС). Как правило, эта система мембран ориентирована здесь в основном параллельно клеточной оболочке нижней стенки клетки, в то время как близ других клеточных стенок ЭС развита значительно слабее. Отклонения корня от нормального положения приводят в этом случае к некоторому неравновесию внутри клеток корневого чехлика. Статолиты покидают определенные участки ЭС, что вызывает появление изгибов. Гольджиосомы в клетках тоже чаще обнаруживаются близ физически нижней стороны, чем близ верхней, и здесь они активнее. До сих пор, однако, еще не выяснено, можно ли считать ЭС и аппарат Гольджи искомыми структурами, чувствующими земное притяжение. В вопросе о том, какие последствия вызывает перемещение статолитов, тоже нет ясности. Может быть, из-за давления, оказываемого статолитами, изменяется активность связанных с мембранами ферментов (биокатализаторов). Но статолиты благодаря своему положению могут также влиять на активность обмена веществ в органоидах клетки. Самый известный пример этому - зеленая водоросль Сharа, с которой мы познакомимся ближе.
Кроме крахмальных зерен, статолитами, по-видимому, могут служить блестящие тельца, кристаллы белка, хлоропласты, митохондрии и другие структуры. Блестящие тельца обнаруживаются в кончиках одноклеточных ризоидов Сharа. Эти клетки прозрачны и совершенно не восприимчивы к свету, поэтому геотропные ответные реакции легко наблюдать, используя микроскоп. Блестящие тельца имеют высокий удельный вес и состоят, вероятно, из сульфата бария. При вертикальном положении ризоида, то есть при нормальных условиях, они лежат в центре ризоида несколько выше его кончика, и пузырьки Гольджи могут беспрепятственно доходить до растущего кончика (рис. 28, б). Предполагают, что цитоплазма, находящаяся близ кончика, имеет иную консистенцию, чем в других частях ризоида. Если же расположить ризоид горизонтально, то статолиты перекрывают пузырькам Гольджи путь к физически нижней стороне. На физически верхней стороне, у которой при этом пузырьки Гольджи как раз и собираются, происходит искривление (рис. 28, в). Таким образом, действие блестящих телец основано на топографическом эффекте. Если переместить статолиты из кончика ризоида в сторону его основания, то исчезнет способность к геотропной ответной реакции. Лишь после восстановления положения блестящих телец ризоиды Сharа снова станут чувствительными к действию силы тяжести. Следовательно, у этого объекта мы имеем дело с почти полной моделью цепи раздражений, обусловливающих проявление геотропизма.
Если водоросли, мхи, папоротники и семенные растения имеют пластиды, то у грибов этих органелл нет. Поэтому здесь в качестве статолитов должны функционировать какие-то другие подвижные частицы. Но что это такое - кристаллы, митохондрии или субмикроскопические структуры, - еще не известно. Например, у обладающих геотропизмом спорангиеносцев Phycomyces до сих пор не удалось обнаружить перемещения каких-либо видимых компонентов клетки.
В результате восприятия растением земного притяжения возникает возбуждение, которое, как и при фототропизме, характеризуется поперечной поляризацией. У высших растений она проявляется в первую очередь в неравномерном распределении ауксина. Сначала рассмотрим это на примере побега, в котором при горизонтальном его положении происходит поперечное перемещение ауксина. Поэтому идущее от вершины к основанию продольное передвижение ауксина под действием силы тяжести на физически верхней стороне замедляется, а на нижней - убыстряется. Как показали опыты, проведенные на колеоптилях, оба эти процесса происходят независимо один от другого. Из-за обогащения ауксином клеток нижней стороны усиливается рост на этой стороне органа. Она становится выпуклой, а стебель или колеоптиль изгибается вверх. Если, например, отрезать верхушку колеоптиля и положить ее горизонтально, то это вызовет неравномерное распределение ауксина в ней. Если снова поместить эту верхушку на то место колеоптиля, где она была раньше, то колеоптиль изогнется (рис. 29, г-а). Следовательно, уже упомянутое нами правило "без ауксина нет роста" приложимо и к вызванным геотропизмом изменениям положения стеблей в пространстве. Согласно современным представлениям, ауксин выводится из клеток связанными с плазмалеммой пермеазами (особыми белками, транспортирующими вещества через биологические мембраны). По-видимому, в приведении их в состояние активности какую-то роль играет давление, оказываемое статолитами. Предстоит также выяснить, почему именно там, где находятся статолиты, увеличивается выделение ауксина. Следует также отметить взаимосвязь проведения возбуждения и активного перемещения веществ.
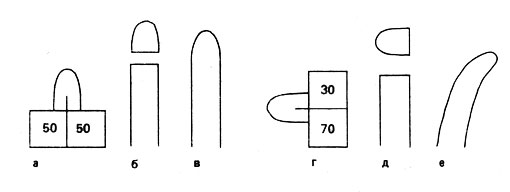
Рис. 29. Вызванное геотропизмом перемещение ауксина в колеоптилях (числа показывают относительное содержание ауксина в агаровых блоках). Горизонтальное положение колеоптилей приводит к асимметричному распределению ауксина (г-е)
У корней эти явления труднее объяснимы, чем у стеблей. В принципе геотропные ответные реакции и здесь можно было бы объяснить поперечным передвижением ауксина под влиянием силы тяжести. Поскольку корни в сравнении со стеблями значительно чувствительнее к ауксину, та сторона органа, где ауксина меньше, растет сильнее. Поэтому горизонтально лежащий корень изгибается в отличие от стебля вниз. Но, к сожалению, эта простая гипотеза экспериментально совсем не подтверждена. Поперечное передвижение ауксина обнаруживается только около кончика корня. Оно пропадает, если удалить корневой чехлик. А зона изгиба корня находится несколько выше его кончика. В проведении возбуждения ауксин тоже не участвует, так как оно распространяется от кончика корня к зоне растяжения, в то время как ауксин движется в противоположном направлении, к кончику. Как показали последние исследования, восприятие земного притяжения вызывает поперечное перемещение к нижней стороне органа какого-то задерживающего рост вещества. Продольное передвижение такого вещества, осуществляющееся преимущественно от верхушки к основанию, тоже еще должно быть доказано.
В заключение попытаемся интерпретировать цепь геотропных ответных реакций как кибернетическую, то есть саморегулирующуюся систему. При этом примем вертикальное положение органа за исходное, нормальное. Статолиты можно считать рецепторами отклонения от нормального положения, а зону роста - местом, реагирующим на информацию, поступающую от рецепторов. О приемлемости такой модели свидетельствуют, между прочим, и "колебательные движения", о которых мы уже говорили при обсуждении геотропных изгибов побегов Соlеus.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'