Фототропизм
Среди факторов, вызывающих проявление тропизмов, свет был, бесспорно, первым, на действие которого человек обратил внимание. Уже в древних литературных источниках были описаны изменения положений органов растений, названные гелиотропными. "Гелиос" означает по-гречески "солнце", а гелиотропный - "направленный к солнцу". Греческий философ Теофраст (371-286 гг. до н. э.), которого считают основоположником научной ботаники, упоминал, например, что почти все листья обращены своими верхними сторонами к свету. Был известен и гелиотропизм некоторых цветков. Так, римский ученый Варрон (116-27 гг. до н. э.) отмечал, что цветки, которые называют гелиотропными, с утра обращены в сторону восходящего солнца и в течение дня следуют за ним вплоть до его заката, будучи все время обращенными к нему. Особенно четко такое поведение выражено у видов рода гелиотроп (Heliotropium) из семейства бурачниковых. Вероятно, вам знаком Heliotropium arborescens - широко известное разводимое на балконах растение с крупными фиолетовыми пахнущими ванилью цветками. Но поскольку такого рода ответную реакцию растения можно вызвать и искусственным освещением, изначальное название "гелиотропизм" уступило место термину "фототропизм". Из органов растений, отклоняющихся в сторону источника света (положительный фототропизм), прежде всего назовем молодые побеги. Корни же афототропны, то есть свет не оказывает влияния на направление их роста. Отрицательно фототропны зародышевые корешки некоторых растений (рис. 16), усики с присосковидными дисками девичьего винограда (виды Рarthenocissus) и усики других цепляющихся растений, корни-присоски плюща (Hedera helix), гипокотиль (участок стебля между главным корнем и семядолями) развивающейся из семени омелы (Visсum), а также ризоиды печеночных мхов и заростков папоротников. Плагиотропным называют положение многих боковых ветвей семенных растений, а трансверзально, то есть поперечно-фототропным, - положение большинства листьев листостебельных растений и талломов печеночных мхов. Фототропные ответные реакции растений можно наблюдать как в природе, так и у комнатных растений. Как раз комнатные растения особенно часто реагируют на одностороннее освещение соответствующими ростовыми движениями - характерным положением стебля и листьев. Например, если вы перенесете колеус (Coleus blumei), о котором мы еще будем говорить, из теплицы, где свет падал со всех сторон, или из сада в комнату, имеющую лишь одно окно, то вскоре увидите изгиб в сторону источника света. Этот простой эксперимент легко продолжить, повернув примерно через неделю вазон с растением на 180°. Если в вашем распоряжении несколько экземпляров этого растения, то, кроме поворота вазона на 180°, вы можете и по-иному менять положение объектов, а также оставить один экземпляр в качестве контрольного.
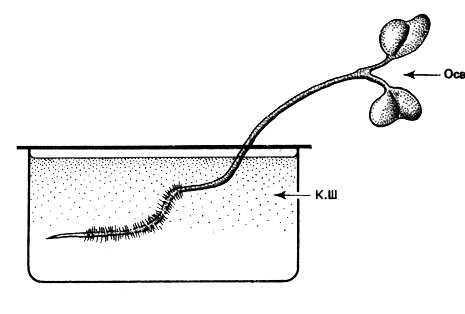
Рис. 16. Влияние одностороннего освещения (Осв) на проросток горчицы белой (Sinapis alba) в водной культуре. Ось побега положительно, а зародышевый корешок отрицательно фототропны (К.Ш - корневая шейка); семядоли расположены под прямым углом к лучам света (поперечный фототропизм)
Многие растения солнечных местообитаний могут располагать свои листовые пластинки в направлении с севера на юг. Поэтому их называют компасными растениями. По обочинам дорог (например, в Тюрингии и Саксонии) можно наблюдать теплолюбивый латук дикий, или компасный (Lactuca serriоlа) (В СССР это растение распространено в южной половине европейской части страны, на Кавказе, в Средней Азии, в южных областях Сибири). Его очередно расположенные на стебле, сидячие, почти вертикально направленные листья поворачиваются у своих оснований так, что их пластинки оказываются в плоскости, показывающей направление с севера на юг. Благодаря такому своеобразному положению листьев относительно слабые утренние и вечерние солнечные лучи попадают на всю поверхность листа, а яркие полуденные - на "ребро". Подобным же образом ведет себя и североамериканское растение Silphium lасinatum, которое считают типичнейшим компасным растением. Этот крупный, достигающий высоты от 1 до 4 м многолетник тоже относится к семейству сложноцветных и растет на просторах прерий. Охотники уже давно знают его как компасное растение, поскольку по положению его листьев они даже в пасмурную погоду могут определить страны света. Правда, кроме освещения, на компасные растения оказывают влияние также тепловое излучение и некоторые другие факторы.
Низшие растения тоже обладают фототропными реакциями. Из грибов, например, наиболее хорошо исследованы принадлежащие семейству Mucoraсеае виды родов Phycomyces и Pilobolus, у которых положительный фототропизм обнаруживают спорангиеносцы.
Во время развития растения фототропизм его органов может изменяться. Так ведут себя, например, цветоножки цимбалярии постенной (Суmbalaria muralis), проявляющие сначала положительный фототропизм, а после совершившегося в цветках оплодотворения, ко времени созревания плодов - отрицательный (рис. 17). Благодаря этому семена цимбалярии, растущей на каменистых обрывах и стенах, попадают в трещины скал и в щели каменных стен. Эти местообитания благоприятны для прорастания семян этого растения и представляют собой экологические ниши, в которых оно существует. Другими примерами изменения тропизмов могут быть цвето- и плодоножки солнцецвета монетолистного (Helianthemum nummularium) и настурции большой (Tropaeolum majus). Некоторые части проростков, например, такие, как колеоптили, гипокотили и эпикотили (участки стебля между семядолями и первыми надсемядольными листьями), ведут себя по-разному в зависимости от освещенности. При этом количество света от 10 до 10000 лк*с приводит к первой положительной ответной реакции, а свыше 10000 лк*с - ко второй. У колеоптилей овса количество света от 10000 до 100000 лк*с вызывает даже отрицательную реакцию. Если сила света исключительно велика, снова наступает уменьшение изгибания, то есть третья - положительная - реакция. Вторую положительную реакцию можно считать естественной фототропной реакцией, тогда как сила света, необходимая для проявления растением третьей положительной ответной реакции, вряд ли может быть достигнута в природных условиях.
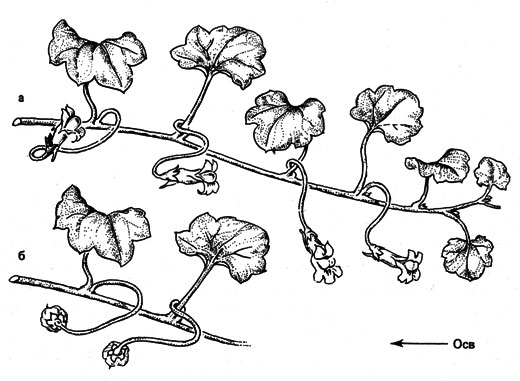
Рис. 17. Положительный фототропизм цветоножек (а) и отрицательный - плодоножек (б) цимбалярии постенной. Изменение происходит после оплодотворения. Осв - направление лучей света
Благодаря изгибаниям, обусловленным фототропизмом, органы растений занимают наиболее выгодное положение. Например, определенное положение листьев способствует оптимальному использованию света при фотосинтезе. Следовательно, способность к такого рода изменениям положений органов экологически оправдана и поэтому была наследственно закреплена в ходе эволюции.
А теперь, после ознакомления с характером ответных реакций, рассмотрим, как осуществляются фототропные движения. При положительном фототропизме изгибание органа происходит прежде всего в результате задержки роста освещенной стороны органа и усиления роста на затененной его стороне. Особое значение здесь имеет ауксин. Поскольку те части побегов, с которыми связан рост в длину, довольно велики, участки возможных их искривлений тоже длинны, а радиусы искривлений также значительны. Напротив, корни, у которых зоны роста относительно коротки, имеют короткие участки, способные изгибаться, и небольшие радиусы искривлений. То же относится и к спорангиеносцам Pilobolus и Phycomyces, у которых менее освещенная сторона также сильнее растет. Место восприятия раздражения светом находится обычно ближе к вершине органа, чем зона его изгибания, и между этими участками осуществляется проведение раздражения. Если изгибание, начавшееся у вершины органа, распространяется к его основанию, то обнаруживается апикальная (верхушечная) ответная реакция. Если же изгибание начинается у основания (базиса), то речь идет о базальной ответной реакции. Например, колеоптили, постоянно используемые для проведения экспериментов, при первой положительной реакции воспринимают свет только самыми верхними участками (длиной около 250 мкм). Это можно доказать, закрыв верхушку алюминиевой фольгой, что задержит изгибание. При второй положительной реакции апикальная область восприятия света оказывается несколько большей (около 500 мкм), но и базальные участки колеоптилей обнаруживают небольшую восприимчивость. Время, проходящее после начала действия раздражителя до наступления ответной реакции, при первом изгибании колеоптилей овса в сторону источника света составляет от 25 до 60 мин, а продолжительность осуществления реакции (от начала до завершения изгибания) - 24 ч. Спорангиеносцы же Рilоbоlus изгибаются значительно быстрее, поскольку продолжительность их ответной реакции не достигает и 2 ч.
Изгибание органов и одноклеточных структур происходит, как уже упоминалось, в ту сторону, на которой рост менее значителен (рис. 18, в). Однако при строго верхушечном росте фототропно реагирующие клетки ведут себя по-иному. Например, верхушечный рост несозревших спорангиеносцев Рilоbolus и пыльцевых трубок при одностороннем раздражении приостанавливается. Но в это же время подвергшаяся раздражению сторона столь сильно растет, что возникает новая верхушка, обращенная к свету (рис. 18, а). Таким образом, здесь положительно фототропное изгибание происходит в результате более интенсивного роста той стороны, которая обращена к свету. При движении листьев, обладающих поперечным фототропизмом, воспринимать световые раздражения могут, во-первых, либо черешки, либо листовые пластинки и, во-вторых, обе эти части листьев. Поперечно-фототропное положение листьев, в общем, обусловлено изгибанием как черешков, так и листовых пластинок. К этому могут быть причастны и скручивания черешков. Наконец, следует также упомянуть, что поперечно-фототропное положение листьев может быть обусловлено и изменениями тургорного давления. Известные примеры тому - листья бобовых и мальвовых, обладающие сочленениями.
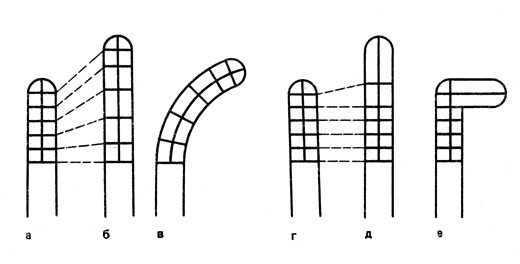
Рис. 18. Обусловленное фототропизмом изгибание отдельных клеток при интеркалярном (а-в) и строго верхушечном (г-е) росте: а и г - исходные состояния, б и д - рост без раздражения светом, вне - рост при одностороннем освещении
В связанных с фототропизмом ответных реакциях растений проявляются некоторые закономерности, о которых речь шла при рассмотрении основ физиологии раздражений. Так, действие закона количества раздражения особенно хорошо обнаруживается при восприятии раздражений. Например, у колеоптилей овса, порог раздражения которых находится между 3 и 25 лк*с, при увеличении количества света примерно до 100 лк*с сохраняется некоторое соответствие между количеством раздражения и величиной ответной реакции. Если же освещать колеоптили с двух строго противоположных сторон, то для того чтобы вызвать изгиб в какую-либо сторону, интенсивность освещения с этой стороны должна превышать другую минимум на 3%. Но у спорангиеносцев Рilоbоlus это происходит лишь при разнице не менее чем 20%, поскольку здесь способность к восприятию различий менее выражена. Таким образом, как колеоптили, так и спорангиеносцы демонстрируют проявление закона Вебера - Фехнера. Если растения, обнаруживающие фототропизм, освещать с разных сторон, но так, чтобы направления лучей от двух источников не составляли 180°, то изгиб произойдет в сторону равнодействующей, которая возникает в параллелограмме сил обоих раздражений. Следовательно, ответная реакция протекает в соответствии с законом результантов (равнодействующих). В качестве примеров можно привести колеоптили злаков и. проростки двудольных растений.
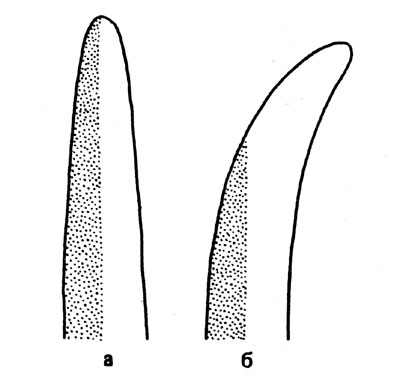
Рис. 19. Освещение половины (светлая краска) той стороны колеоптиля, которая обращена к читателю, светом, падающим перпендикулярно плоскости бумаги (а), и изгибание колеоптиля вдоль плоскости бумаги (б)
Для того чтобы начали осуществляться изменения положений органов, обусловленные фототропизмом, необходимо, как и при ориентировочных движениях хлоропластов и при проявлении фототаксиса, восприятие раздражения светом, При этом воспринимается не направленность световых лучей, а различие в освещенности обращенной к источнику света и затененной сторон. Это обнаруживается как при освещении лишь половины одной из сторон органа, так и при одностороннем освещении пустых колеоптилей изнутри. Колеоптиль изгибается вовсе не навстречу свету, а в направлении, определяемом различием в освещенности (рис. 19). Часто различие в освещенности возникает в результате поглощения света и его рассеивания внутри освещенного органа. У прозрачных объектов различие в освещенности может быть вызвано наличием затеняющих пигментов или изменением хода лучей (как в линзе). Затеняющие пигменты мы находим, в частности, у спорангиеносцев Рilоbоlus. A спорангиеносцы Phycomyces действуют как линзы. При нормальных условиях такая клетка, коэффициент светопреломления которой можно считать близким к 1,37, функционирует как собирающая линза (рис. 20). Свет фокусируется на более удаленной от его источника стороне. А поскольку у этого гриба одностороннее усиление освещенности стимулирует рост более освещенной стороны, споран-гиеносец изгибается в сторону источника света. Если же спорангиеносец поместить в вазелиновое масло, коэффициент светопреломления которого 1,47, и осветить с одной стороны, то клетка действует как рассеивающая линза. Направление изгиба в этом случае меняется на противоположное, и обнаруживается отрицательно-фототропная ответная реакция.
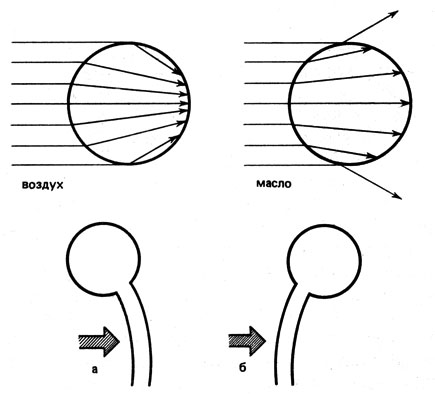
Рис. 20. Зависимость проявления положительно (а) или отрицательно фототропного (б) изгиба спорангиеносца Рhусоmусеs от среды, окружающей объект, и от хода лучей света (см. стрелки)
На движения, обусловленные фототропизмом, особенно влияет сине-зеленый и сине-фиолетовый свет. Для спектров проявления ответных фототропных реакций характерны одна вершина в области ультрафиолетовых и три - в области синих волн (рис. 21). Если три вершины в синей области видимого света хорошо согласуются с кривой поглощения света 8-каротином, то вершина в фиолетовой области указывает на флавины (например, рибофлавин). До сих пор еще не удалось выяснить, что воспринимает свет - каротин или флавин. Может быть, каротиноид служит фоторецептором, а флавин - затеняющим пигментом, способствующим созданию достаточного различия в освещенности. Другие гипотезы здесь мы рассматривать не будем.
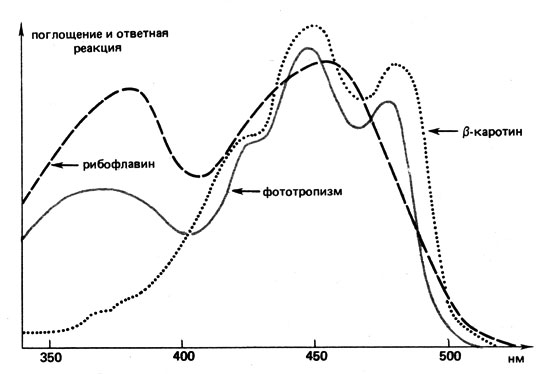
Рис. 21. Спектры поглощения света β-каротином и рибофлавином, а также спектр проявления ответной реакции при фототропизме (первый изгиб колеоптилей овса в сторону источника света)
На развивающиеся из спор протонемы мхов и заростки папоротников фотохимически действует в первую очередь длинноволновый свет. Поэтому ответная реакция этих объектов, обусловленная фототропизмом, связана с системой фитохромов. Наряду с этим, вероятно, свет поглощается и каким-то флавопротеидом. А поперечное по отношению к световым лучам положение листьев определяется влиянием и красного света. Очевидно, при этом рецепторами оказываются хлорофилл и фитохром.
Как уже упоминалось при рассмотрении движений хлоропластов, находящийся в эктоплазме фитохром расположен очень упорядоченно: молекулы неактивного фитохрома 660 лежат параллельно поверхности объекта, а молекулы активного фитохрома 730 - перпендикулярно ей. О локализации и расположении в клетке каротиноидов и флавинов известно значительно меньше. Для Рilоbоlus, например, характерно наличие содержащего каротиноиды утолщения в основании несущего спорангий пузыревидного расширения спорангиеносца (рис. 22). Как показывает этот рисунок, лучи света, попадающие на черный спорангий, дальше не проходят. Если направление светового потока меняется, то спорангиеносец изгибается до тех пор, пока содержащее каротиноиды утолщение снова не станет равномерно освещенным. Хотя каротиноиды мы обнаруживаем прежде всего в хлоро- и хромопластах, они встречаются и у грибов, не имеющих пластид. Распределение находящихся в основной протоплазме каротиноидов и флавинов, соответственно каротиноид-протеидов и флавин-протеидов, еще не выяснено. Однако принято считать, что эти пигменты расположены не столь упорядоченно, как молекулы фитохрома.
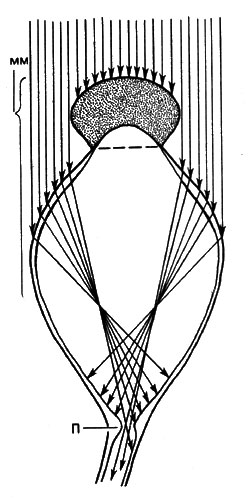
Рис. 22. Схематическое изображение хода лучей в симметрично освещаемом сверху спорангиеносце Рilоbоlus. Вследствие того что пузыревидное расширение спорангиеносца ведет себя как линза, лучи света попадают на содержащее пигмент утолщение (П), находящееся в основании расширения
Следующая за восприятием раздражения, то есть за поглощением света, фаза возбуждения характеризуется поперечной поляризацией. Особенно интенсивно исследовалась роль ауксина при изгибании колеоптилей овса. Вследствие асимметричного проведения ауксина возникает неодинаковое его распределение. Это проведение требует затрат энергии и присутствия в атмосфере кислорода. Физиологическая асимметрия возникает у верхушки. Уже до того как изгиб станет заметным, концентрация ауксина выше в клетках той стороны, которая станет выпуклой, то есть будет быстрее расти. Для возникновения движения ауксина поперек колеоптиля овса достаточно тех количеств света, которые вызывают первую и вторую ответные реакции. Экспериментально можно показать, что поток ауксина от вершины к основанию несколько сдвинут в одну сторону (рис. 23). Если с помощью тонкой стеклянной пластинки воспрепятствовать поперечному движению ауксина, то колеоптиль не изогнется (рис. 23, г). Такие же результаты получают, когда с помощью кусочков агара собирают ауксин из отрезанных верхушек колеоптилей (рис. 23, е-з). При беспрепятственном поперечном движении ауксина асимметричное его распределение возникает, но не возникает, если этого движения не допустить (рис. 23, ж и з). Механизм проведения раздражения идентичен асимметричному проведению ауксина. Одностороннее освещение приводит не только к поперечной поляризации, но и к торможению потока ауксина от верхушки к основанию. Это торможение происходит под влиянием тех же областей спектра, что и изгиб, обусловленный фототропизмом.
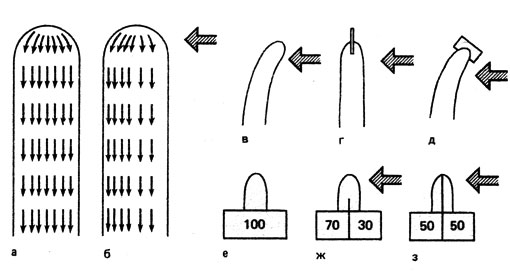
Рис. 23. Схема движения ауксина при освещении колеоптиля со всех сторон (а и е), а также поперечной поляризации потока ауксина при одностороннем освещении в направлении, указанном стрелками (б, в, д и ж). Когда поперечный поток ауксина прерван, неравномерного распределения ауксина не возникает, а, следовательно, не происходит и изгиб (г и з). Числами обозначено относительное содержание ауксина
Долгое время в научной литературе в качестве альтернативы положению об асимметричном проведении ауксина обсуждалась гипотеза индуцированного светом окисления ауксина как причины неодинакового его распределения. К такому мнению приходили в связи с тем, что во время опытов, проводимых в пробирках, ауксин легко окислялся. Однако в клетках обращенной к свету стороны органа не происходит сколько-нибудь заметного окисления ауксина.
В эффективности действия ауксина можно убедиться, если смешать 0,005%-ный раствор этого стимулирующего рост вещества с хлопковым маслом и нанести немного этой пасты на междоузлие относительно молодого растения. Уже через несколько часов нанесенный ауксин вызовет в этом месте рост в длину, что приведет к искривлению стебля.
Неравномерное распределение ауксина оказывается очень важным, но не единственным следствием поперечной поляризации. Возникают также физико-химические, связанные с физиологией обмена веществ различия между освещенной и затененной сторонами, в частности, снижение содержания сахара и концентрации ионов водорода в клетках освещенной стороны. Даже у тех организмов, у которых в осуществлении ответных реакций, обусловленных фототропизмом, ауксин не участвует, при одностороннем освещении возникает перепад в содержании некоторых веществ в тканях освещенной и затененной стороны. Примерами могут служить спорангиеносцы Phycomyces и Pilobolus. Правда, химическая природа асимметрично распределяющихся в них веществ пока еще не известна.
Кроме рассмотренных нами до сих пор движений, обусловленных фототропизмом, имеются и ответные реакции, тоже возникающие под влиянием действия света, но проявляющиеся по-иному, чем фототропные. Здесь мы ограничимся кратким рассмотрением ростовой реакции в ответ на освещение, а также скототропизма.
Ростовой реакцией в ответ на освещение называют изменение скорости роста, наступающее вследствие изменения интенсивности светового потока. Его можно наблюдать как после затемнения, так и после увеличения освещенности. Во избежание появления фототропных изгибов необходимо, чтобы свет падал на растение со всех сторон и симметрично. Положительная ответная ростовая реакция проявляется, если скорость роста возрастает (в частности, у спорангиеносцев Phycomyces). Напротив, если усиление освещенности вызывает задержку роста, то говорят об отрицательной ростовой реакции, возникающей в ответ на освещение (например, у колеоптилей овса). В противоположность движениям, связанным с фототропизмом, ростовая реакция, возникающая в ответ на освещение, - это только временное явление, и наступающие при этом задержки и ускорения роста вскоре снова компенсируются противоположными ответными реакциями. Фототропные же изгибы, напротив, продолжаются, как правило, в течение нескольких часов. Несмотря на эти различия, зависимость тех и других ответных реакций от спектрального состава света одинакова. Попытки причислить фототропизм к ростовым ответным реакциям приносили лишь частичный успех.
Под скототропизмом понимают рост в сторону затенения. Это явление можно наблюдать у некоторых тропических лиан. Например, проростки представителя семейства ароидных Monstera gigantea дорастают до дерева, которое эти растения будут затем использовать как опору, в результате связанных с ростом изгибов в сторону наиболее темного сектора горизонта. К тому же ствол этого дерева должен как бы "закрывать" отрезок горизонта величиной в несколько угловых градусов. И это не проявление отрицательного фототропизма: проростки нарастают на дерево со всех сторон. По достижении цели проявлявший скототропизм проросток становится растением, способным к движениям, обусловленным положительным фототропизмом, и растущим навстречу пробивающимся лучам света, т. е. в сторону кроны дерева.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'