Механика движений
Благодаря все более и более совершенствовавшимся методам микроскопических исследований смогли стать понятными разные механизмы движения. Кроме известного вам, вероятно, движения с помощью жгутиков мы рассмотрим в дальнейшем также амебоидное движение и движение посредством скольжения.
К свободному передвижению с помощью жгутиков способно множество растительных и животных организмов. Этот тип движения мы обнаруживаем у бактерий, водорослей (например, у флагеллят, у зооспор и гамет зеленых и бурых водорослей), у некоторых грибов (зооспоры и гаметы), а также у сперматозоидов мхов, папоротников и голосеменных растений (Gymnospermae). Таким образом, во всех отделах мира растений, за исключением сине-зеленых водорослей, встречаются либо имеющие жгутики представители, либо одноклеточные, тоже имеющие жгутики стадии развития (Все семенные растения (как голо-, так и покрытосеменные) автор относит к одному отделу, что выглядит некоторым анахронизмом. Поэтому отметим здесь, что мужские гаметы покрытосеменных растений (спермин), как и многих голосеменных (например, хвойных), жгутиков не имеют. Нет жгутиков и у мужских гамет (спермациев) красных водорослей).
Жгутики, называемые также флагеллами или ресничками, представляют собой похожие на бичи выросты на поверхности клетки. Как показали современные исследования, проведенные с применением электронных микроскопов, жгутики всех эукариот обладают далеко идущим сходством в строении. Если принять во внимание огромные морфологические и физиологические различия между водорослями, грибами, мхами, папоротниками и голосеменными растениями, а также животными и человеком, то это, без сомнения, поразительно. Наряду с универсальностью генетического кода единообразие в строении этих органелл, осуществляющих движения, служит еще одним доказательством общего происхождения жизни, существующей ныне на нашей планете.
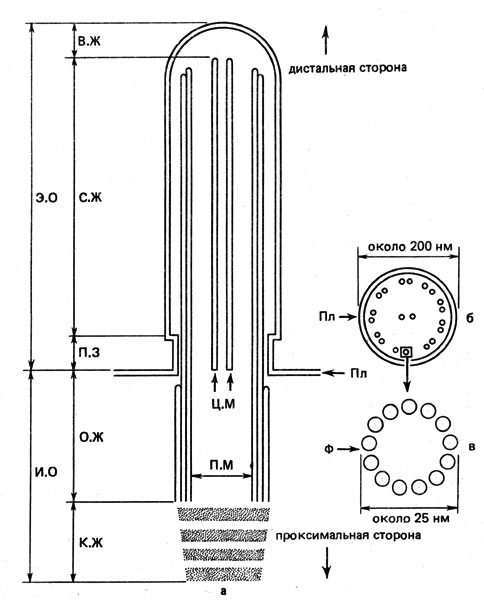
Рис. 10. Схема строения жгутика в продольном (а) и поперечном (б) сечениях, а также строения микротрубочки (в): В.Ж - вершина жгутика, И.О - интрацеллюлярный (находящийся внутри клетки) участок жгутика, К.Ж - корень жгутика, О.Ж - основание жгутика; П.З - переходная зона, Пл - плазмалемма, П.М - периферические микротрубочки, С.Ж - стержень жгутика, Ф - фибриллы (диаметром около 4 нм), Ц.М - центральные микротрубочки, Э.О - экстрацеллюлярный (находящийся вне клетки) участок жгутика
Главные части жгутика - это покрывающая его мембрана и аксонема, или комплекс осевых нитей. Мембрана представляет собой выдающийся наружу вырост плазмалеммы. Комплекс осевых нитей состоит из 20 микротрубочек, причем 9 пар (дублетов) микротрубочек расположены вокруг двух микротрубочек, находящихся в центре (рис. 10, б). Микротрубочки - это трубчатые образования, имеющие диаметры от 20 до 30 нм, состоящие каждое из 13 фибриллоподобных нитей (рис. 10, в). Эти нити сложены глобулярными молекулами белка тубулина и винтообразно закручены вокруг оси. Их длина может достигать многих микрон. В самой вершине жгутика, как правило, присутствуют лишь обе центральные микротрубочки, в переходной зоне - только периферические, а в основании - 9 триплетов, то есть групп из трех микротрубочек каждая (см. рис. 10, а). Длина жгутиков обычно бывает от 10 до 70 мкм, но иногда и более 100 мкм, тогда как поперечник, как правило, от 100 до 300 нм. Наружный отрезок жгутика, то есть собственно жгутик, состоит из кончика (вершины), стержня, а также в большинстве случаев и из переходной зоны. К внутреннему отрезку жгута относят его корень, основание (блефаропласты), а иногда и переходную зону. В корне жгутика тоже находятся фибриллярные элементы. Между мембраной и микротрубочками до сих пор не обнаружено никаких структур. Верхушку жгутика называют дистальным его участком, а корень - проксимальным. Жгутики, расположенные на переднем конце клетки, называют акроконтными, находящиеся на задней ее стороне, - опистоконтными, а при размещении их по всей поверхности клетки говорят о перитрихальном их расположении. Часто жгутики покрыты мерцательными волосками.
Хотя жгутики и обладают единообразным строением, формы их движений очень различны. Наряду с гребными движениями (рис. 11) можно наблюдать и волнообразное или подобное движению бича, при котором весь жгутик, изгибаясь от основания до вершины, колеблется подобно волне. Кроме того, жгутики способны к винтообразным движениям, при которых клетка начинает вращаться. Удивительна мощность такого рода "мотора". В сравнении со скоростью современных транспортных средств скорости движения от 10 до 100 мкм/с можно считать скромными. Но требующаяся для этого частота движений жгутика (от 40 до 50 в течение секунды) представляет собой достойный внимания физиологический феномен, тем более что жгутики двигаются в жидкой среде, сопротивление которой значительно превышает сопротивление воздуха.
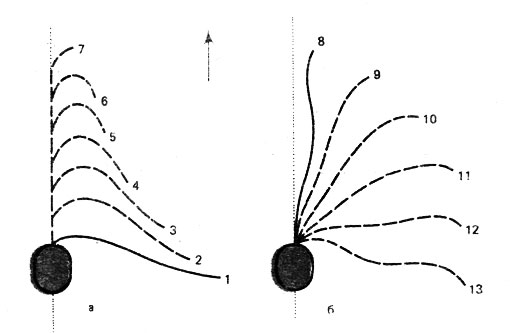
Рис. 11. Гребное движение жгутика одноклеточного жгутиконосца Monas sp. (стрелка указывает направление движения клетки в воде): а - возвратное движение жгутика, б - активный удар
Для объяснения внутренней механики движения жгутиков, т. е. волнообразно распространяющихся по ним укорочений и удлинений, предложены две взаимоисключающие гипотезы. Согласно первой, сами микротрубочки должны укорачиваться и удлиняться. В соответствии со второй микротрубочки скользят одна вдоль другой. Это второе объяснение более вероятно. Энергия, необходимая для движения жгутиков, предоставляется клетками в форме АТФ.
Жгутики относящихся к прокариотам бактерий тоже единообразны, но устроены они значительно проще. У них нет микротрубочек. На поперечном срезе обнаруживаются "лишь две фибриллы, а диаметр достигает 10-30 нм. Нет и элементарной мембраны. Особенно интересно состоящее из нескольких дисков и сильно дифференцированное базальное тело, закрепляющее жгутик в плазмалемме и в клеточной оболочке. По новейшим представлениям, жгутик бактерии должен не менять свою форму, а вращаться в своем "гнезде". Возможно, что речь идет о единственном в мире организмов случае, в котором осуществился принцип колеса, имеющего возможность свободно вращаться.
После того как мы относительно подробно рассмотрели движение с помощью жгутиков, обратимся к амебоидному типу движения. Он встречается, в частности, у некоторых флагеллят, миксомицетов (слизистых грибов, или слизевиков), а также у немногих других грибов. Эта форма движения осуществляется путем образования псевдоподиев. Псевдоподии - это весьма разнообразной формы и величины цитоплазматические отростки, которые могут спонтанно возникать в любом месте. Их образование зависит в первую очередь от вязкости цитоплазмы и происходит в связи с движением самой цитоплазмы. Особенно хорошо исследованные в этом отношении объекты представляют собой плазмодии слизистых грибов, которые в виде тонкого слоя цитоплазмы могут ползти по субстрату, постоянно менять свою форму и достигать в поперечнике 30 см. Мы находим их, в частности, на разрушающейся древесине, на опавших листьях и на лесной почве. Скорость амебоидного движения относительно невелика и в среднем определяется как 0,5-3 мкм/с. Это соответствует отрезку пути примерно от 1,8 до 10,8 мм, пройденному за час. Если у вас появится возможность, например, прогуливаясь по лесу, понаблюдать за медленно переползающими плазмодиями, то запаситесь временем и терпением.
Считают, что движение плазмодия обусловлено присутствием в нем белковых фибрилл типа актомиозина, которые взаимодействуют с АТФ. Вероятно, эти белки играют определенную роль как в превращении золя в гель, так и геля в золь. При этом, возможно, находящаяся внутри плазмодия эндоплазма, пребывающая в состоянии золя, выдавливается желеподобной (имеющей консистенцию геля) эктоплазмой наружу, где она выступает в виде псевдоподия и становится более плотной, превращаясь в эктоплазму.
Еще одна форма движения растений - это скольжение, которое встречается как у бактерий и сине-зеленых водорослей, так и у настоящих водорослей (эвгленовых, диатомовых и дисмидиевых). Хотя причины, его вызывающие, весьма различны, картины его проявления очень сходны, поскольку мы не обнаруживаем ни внешне различимых изменений формы организмов, ни образования выступов, обусловливающих перемещение. Для скольжения особенно характерна тесная связь между выделением веществ и обеспечением поступательного движения, В большинстве случаев перемещение связано с образованием слизи. Выделение слизи, во-первых, можно считать причиной движения, а во-вторых, она служит смазочным материалом. Химически слизи представляют собой макромолекулярные сложные полисахариды, возникающие из веществ, образующихся при фотосинтезе.
Объектами многократных исследований были сине-зеленые водоросли, чьи гормогонии (участки нитей, состоящих из клеток) способны к относительно быстрым и хорошо заметным движениям посредством скольжения. Например, одетые слизистым чехлом многоклеточные нити анабены (Anabaena vаriabilis) могут, во-первых, активно двигаться вперед и назад по полоске слизи, а во-вторых, скользить в одном и том же направлении. Эти полоски слизи становятся хорошо различимы при использовании фазово-контрастного микроскопа. Скорость перемещения Anabaena variabilis составляет примерно 20 мкм/мин. Таким образом, скольжение происходит, в общем, медленнее, чем движение с помощью жгутиков или амебоидное. Применение же киносъемки позволяет еще лучше продемонстрировать скольжение Anabaena variabilis. Благодаря замедленной киносъемке при увеличениях, получаемых с помощью микроскопа, плотно переплетенные нити сине-зеленой водоросли являют собой при демонстрации фильма в высшей степени динамичную картину. Использование кинокамеры и здесь оказывается одинаково ценным как для проведения исследований, так и для учебных целей. У представителей некоторых родов сине-зеленых водорослей во время перемещения нитей происходит и их вращение. Привлекающее к себе внимание перемещение посредством скольжения свойственно диатомеям (кремневым водорослям), но только тем из них, которые имеют так называемый шов (щель, расположенную в плоскости симметрии кремневого панциря). Поэтому этот шов рассматривается некоторыми исследователями именно как органелла движения. Вдоль шва выделяется студнеподобное вещество, или "субстанция движения", которое набухает и приклеивается к субстрату. Возникновение необходимых для движения усилий связано, вероятно, с особым слоем цитоплазмы, который находится вне шва. Возможно, какую-то роль при этом играют и фибриллы. Скольжение десмидиевых водорослей, может быть, представляет собой движение посредством отталкивания: выделяемая ими слизь набухает и в результате этого отодвигает клетку.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'