Зависимость свободных перемещений от внешних факторов
Как и все другие процессы жизнедеятельности, движения тоже зависят от действия внутренних и внешних факторов. К числу важнейших внутренних факторов относят, в частности, генетически обусловленные особенности, соответствующую стадию развития, а также ритмические изменения "физиологического состояния". Из внешних факторов, наиболее сильно влияющих на движения, назовем свет, температуру, химические соединения и земное притяжение. Например, способ движения свободно перемещающегося организма (с помощью жгутиков, амебоидно или скольжением) в значительной степени зависит от его наследственно обусловленных особенностей. Если механика движений, уже описанная в предыдущем разделе, проявляется без специфического воздействия какого-либо из внешних факторов, то говорят об автономных движениях и противопоставляют их индуцированным, то есть вызванным раздражениями, пришедшими извне. Хотя четкое разграничение часто затруднено, все же как раз исследование влияния внешних факторов привело к получению важных сведений о проявлении движений и управлении ими.
Вызванные внешними раздражениями упорядоченные свободные перемещения именуют таксисами. Если движение происходит в направлении источника раздражения, то перед нами положительный таксис. В противоположном же случае говорят об отрицательном таксисе. По характеру ответной реакции различают топотаксисы и фоботаксисы. Если топотаксис представляет собой целенаправленное изменение направления движения, то фоботаксис производит впечатление "реакции испуга" ("фобос" по-гречески значит "страх" или "ужас"), и при этом организм попадает в область оптимального действия раздражения, так сказать, окольными путями. Например, ответные реакции сперматозоидов мхов и папоротников - топотаксисы, а бактерий - фоботаксисы. Сине-зеленые водоросли и настоящие водоросли обнаруживают и те и другие изменения направлений движения.
Свободные перемещения, вызываемые действием света, встречаются у бактерий, сине-зеленых водорослей, настоящих водорослей и слизистых грибов. При этом речь прежде всего идет о фотоавтотрофных организмах, которые благодаря этому могут находить наиболее благоприятные места. Положительный фототопотаксис, как правило, можно наблюдать при слабой освещенности. Сильная освещенность, напротив, вызывает отрицательный фототопотаксис. Ответные реакции в виде фотофоботаксисов могут быть вызваны уменьшением и увеличением освещенности. Поскольку при этих процессах воспринимается не направление, откуда исходит раздражение, а изменение интенсивности раздражения во времени, бывшие до сих пор привычные термины "положительный и отрицательный фототопотаксисы" больше не применяют. Ведь вместо фототопотаксис можно просто сказать: фототаксис. Некоторые выбранные нами примеры, вероятно, помогут наглядно объяснить влияние света на движения.
У пурпурных бактерий, например, спектр проявления фотофобной ответной реакции во многом совпадает со спектром эффективности фотосинтеза. Следовательно, фоторецепторы обоих процессов (фотосинтеза и движения) должны быть идентичными. У этих организмов удивительно высока чувствительность к различиям в освещенности. Уже понижение ее на 1-3% от исходной может вызвать перемещения клеток. У Rhodospirillum латентный период составляет от 0,1 до 1,5 с. Один из известнейших примеров фотофобной ответной реакции дает Тhiоspirillum. Эта пурпурная бактерия на одном конце имеет пучок жгутиков; пространство, в котором осуществляется движение жгутиков, имеет форму конуса, основание которого всегда ориентировано против направления движения (рис. 12). На препарате под микроскопом, который с помощью специального устройства может проецировать на этот препарат слабо освещенное поле, можно легко наблюдать движение этой бактерии. Как только конец этой спириллы, несущий жгутики, попадает в темноту, она меняет движение на обратное, причем пучок жгутиков разворачивается в противоположную сторону. Поскольку эти бактерии собираются в освещенном поле, его называют световой ловушкой. Но при очень сильном освещении бактерии покидают поле.
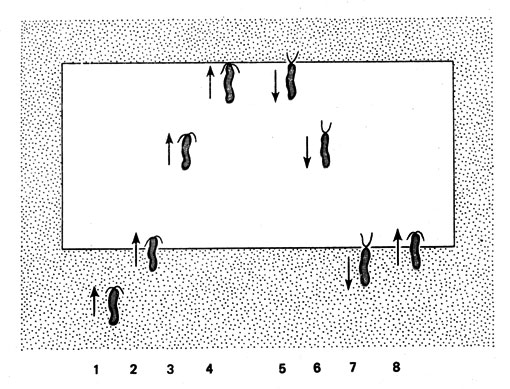
Рис. 12. Положительная фотофобная ответная реакция Тhiоspirillum jenense. Положение 2 - вхождение в светлое поле, положение 4 и 7 - обратная ответная реакция на границе между светлым и темным полями
В противоположность бактериям сине-зеленые водоросли способны к перемещениям, связанным как с фотофоботаксисами, так и с фототаксисами. Если фотофобная ответная реакция вызывает изменение направления движения, то перемещение, обусловленное фототаксисом, совершается в виде поочередных движений вперед и назад, о которых уже шла речь. Под влиянием одностороннего освещения движения в "правильном" направлении длятся дольше, чем в "неправильном". Вопросы, связанные с объяснением восприятия направления светового потока, еще не вполне выяснены.
Для проявления обусловленных фототаксисом движений у сине-зеленых водорослей, например у Рhоrmidium , достаточно коротковолновой области видимого света и определенной части ультрафиолетовой. Напротив, для проявления фотофобной ответной реакции требуется красный свет. Сопоставление соответствующих спектров поглощения со спектрами проявления ответных реакций показывает, что при фототаксисе в восприятии света участвуют каротиноиды и фикобилины, а при фотофоботаксисе - фикобилины и хлорофилл а. Следовательно, эти две ответные реакции проявляются в результате участия разных фоторецепторов. Таким образом, между фотофобными ответными реакциями и фотосинтезом имеется тесная взаимосвязь.
В качестве третьего примера возьмем эвглену - Euglena (рис. 13). Эта имеющая жгутик одноклеточная водоросль живет, например, в прудах, вращаясь в воде с помощью жгутика. При снижении освещенности ее жгутик меняет направление своего движения, а одностороннее освещение приводит к тому, что эти водоросли собираются у той стороны культурального сосуда, которая обращена к свету. Следовательно, эвглене свойствен как фотофобо-, так и фототаксис. У нее спектры ответных реакций располагаются тоже в синей и ультрафиолетовой областях видимого света, но по-иному, чем у сине-зеленых водорослей. А красный свет на нее не действует. Свет воспринимают, по-видимому, каротины, связанные с белками; при этом следует иметь в виду и связанные с белками флавины. О локализации соответствующих пигментов в клетке этой водоросли сейчас многое известно. Если раньше в качестве специфического фоторецептора рассматривали стигму ("глазок"), встречающийся у многих флагеллят, то теперь эту роль приписывают находящемуся в основании жгутика парабазальному телу, представляющему собой местное утолщение жгутика. Парабазальное тело имеется и у тех флагеллят, обнаруживающих фототаксис, у которых стигмы нет. Стигма же во время ротации, т. е. вращательных движений клетки, осуществляет ритмически повторяющиеся затенения истинного фоторецептора. Фототаксис у Euglena следует представить себе как сумму фотофобных ответных реакций, при которых регулярно повторяющиеся затемнения способствуют восприятию направленности светового потока. Это объясняет, почему ответным реакциям и в виде фотофоботаксисов, и в виде фототаксисов свойственны как одинаковые фоторецепторы, так и сходные спектры проявления.
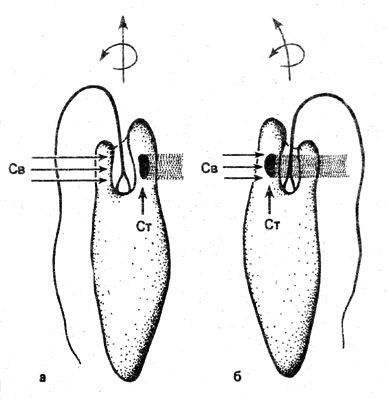
Рис. 13. Клетки эвглены (Еuglenа) при боковом освещении (Св). При поворотах клетки около ее продольной оси фоторецептор, находящийся у основания жгутика, периодически затемняется (б) стигмой (Ст). Это приводит к повороту влево (см. стрелку)
Как показывают эти немногочисленные примеры, можно выявить разные типы ответных реакции. Это касается не только фотофобных ответных реакций бактерий и сине-зеленых водорослей, с одной стороны, и Euglena - с другой, но еще больше - перемещений в пространстве синезеленых водорослей и Euglena, связанных с фототаксисом. Как при реакциях типа фотофоботаксиса, так и фототаксиса свет действует, во-первых, как раздражитель и, во-вторых, как регулирующий фактор. При оценке уровня проявления разных типов ответных реакций у одноклеточных (главным образом) или других мелких организмов, само собою разумеется, нельзя исходить из тех же критериев, какие привлекаются, например, для суждения о деятельности органов чувств позвоночных животных или насекомых. В сравнении с относительной простотой строения этих мельчайших организмов их физиологическая деятельность оказывается удивительной.
Кроме перемещений, вызываемых действием света, к числу важнейших свободных передвижений растительных организмов в пространстве относятся и ответные реакции, обусловленные хемотаксисом. Благодаря им становятся возможными соответствующие способы обнаружения источников питания и слияния гамет. Положительный хемотаксис проявляется в тех случаях, когда движение происходит в том направлении, где концентрация вещества-раздражителя высока. Если же движение происходит в противоположном направлении, то говорят об отрицательном хемотаксисе. Кроме того, различают хемотопотаксис и хемофоботаксис. Вместе с тем в противоположность перемещениям, связанным с фотофоботаксисами, перемещения, вызванные хемофоботаксисами, упорядоченны. Поясним это примером. Представьте себе, что организм, проявляющий реакцию в виде хемофоботаксиса, изменяет направление движения в результате падения концентрации вещества-раздражителя. Хотя такого рода единичная хемофобная ответная реакция не зависит от направления, этот процесс все же будет повторяться до тех пор, пока организм не перестанет передвигаться в сторону снизившейся концентрации. Но при этом в соответствии с принципом "проб и ошибок" он все же переместится туда, где концентрация выше или оказывается оптимальной. В настоящее время известно множество веществ, вызывающих раздражения. Давайте теперь поговорим о связанных с хемотаксисом перемещениях вегетирующих растительных организмов, а затем о хемотаксисе гамет.
Из газообразных веществ, способных вызывать свободные перемещения, в первую очередь следует назвать кислород, сероводород и углекислый газ. В обмене веществ многих организмов эти газы играют важную роль. Например, если вы поместите в освещенное поле зрения микроскопа несколько клеток спирогиры (Spirogyra) - нитчатой зеленой водоросли класса сцеплянок, встречающейся в наших водоемах, - то нередко увидите, что около ее хлоропластов образуется скопление аэробных бактерий. Между прочим, это влияние на бактерии выделяющегося при фотосинтезе кислорода (положительный хемофоботаксис) впервые наблюдал еще в 1881 г. ботаник Энгельманн (1843-1909).
Отрицательный хемофоботаксис обнаруживается у некоторых других бактерий (например, у видов Clostridiurn), Кислород они использовать не могут, и их называют анаэробами.
Имеется много свидетельств и о вызывающих раздражения действиях растворимых веществ. Например, на бактерии брожения и гниения привлекающе действуют не только органические вещества, такие, как глюкоза, лактоза, маннит, декстрин, аспарагин, креатин, мочевина, пептон и мясной экстракт, но также все нейтральные соли, а равно и другие химические соединения. Эти сапрофитно живущие бактерии способны использовать, разлагая, остатки умерших существ. В природе обычно совместно действуют целые группы разных организмов, причем вещества, образующиеся в результате жизнедеятельности одних, могут служить источником питания для других. Возникшие в ходе эволюции бактерии брожения и гниения обладают специфическим обменом веществ и поэтому оказываются не только важными звеньями в цепи биологического круговорота веществ, но и хозяйственно полезными для человека.
Много обнаруживающих хемотаксис видов имеется среди флагеллят. Особенно обстоятельно исследовано поведение одноклеточной, не имеющей хлорофилла водоросли Polytoma uvellа, живущей в разлагающемся иле, для обеспечения существования которой необходимы ненасыщенные жирные кислоты. Восприимчивость этой водоросли к действию раздражителя очень высока. Например, пороговая величина раздражения олеиновой кислотой, вызывающего хемотаксис, соответствует концентрации кислоты около 10-8 мол/л, то есть всего 0,00000282 г/л!
Грибы тоже реагируют на присутствие определенных веществ, по-разному отвечая на раздражения разной интенсивности. Хозяйственное значение могут иметь свободные перемещения грибов-паразитов, находящих своих хозяев благодаря хемотаксису.
Большое значение для сохранения видов имеет хемотаксис, обнаруживаемый гаметами; при этом специфические привлекающие вещества, выделяемые половыми клетками, позволяют им встретиться. Поэтому такого рода вещества называют гамонами, или веществами, способствующими оплодотворению. Как правило, их выделяют женские половые клетки, чтобы вызвать перемещение мужских гамет или активизировать их. Привлечение сперматозоидов Fucus (род бурых водорослей) веществом, выделяемым яйцеклеткой, наблюдали еще в середине прошлого столетия. Но лишь несколько лет назад удалось выяснить химическую структуру этого соединения. Речь идет о серратене, который, как и эктокарпен, образуемый женскими гаметами бурых водорослей рода Ectocarpus, очень летуч и плохо растворим в воде.
К числу наиболее известных привлекающих веществ, способствующих осуществлению полового процесса, относится сиренин гриба Аllоmусеs. Это почти не летучее, растворимое в воде соединение, которое воспринимается мужскими гаметами уже в концентрации 10-10 мол/л. Но следует также отметить, что такая почти фантастическая способность воспринимать раздражение исключительна и свойственна, насколько сейчас известно, только этому хитридиевому грибу.
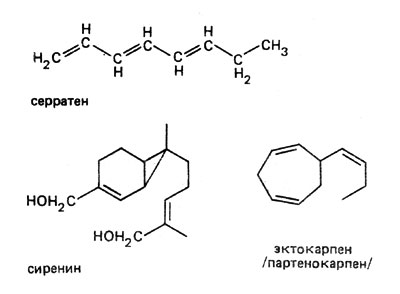
Формулы серратена, сиренина, эктокарпена
О химической природе веществ, привлекающих мужские гаметы мхов и папоротников, мы уже давно осведомлены. По-видимому, эти вещества выделяются клетками архегония, содержащего яйцеклетку, вместе со слизью и служат для привлечения сперматозоидов. У некоторых лиственных мхов таким веществом оказывается сахароза, тогда как у папоротников это широко распространенные и служащие субстратом цикла трикарбоновых кислот (цикла Кребса) яблочная и лимонная кислоты или их соли. Например, у сперматозоидов щитовника мужского (мужского папоротника) хемотаксис вызывает бималат кальция - кальциевая соль яблочной кислоты. Способность воспринимать незначительные различия в концентрации здесь относительно невелика. Иногда хемотаксис обнаруживается лишь при 30-кратном увеличении концентрации яблочной кислоты.
Как показывают исследования хемотаксиса, особенно хемотаксиса гамет, порог раздражения иногда может быть очень низким. Чувствительность систем восприятия у многих растений различна, поскольку относительно невысокой ее специфичности при обнаружении источников питания можно противопоставить исключительно высокую при привлечении гамет. В обоих случаях специфичность чувствительности целесообразна. К восприятию различий в концентрации может быть приложим закон Вебера-Фехнера. Ориентировка осуществляется благодаря механизмам движения. Повышение или снижение концентрации вещества-раздражителя вызывает, например, изменение характера движения жгутиков. Химические соединения могут, подобно свету, вызывать движения и управлять ими. Но для выявления отдельных ступеней процессов, которые при хемотаксисе ведут от раздражения к проявлению ответной реакции, еще требуются серьезные исследования.
О некоторых других таксисах упомянем лишь вкратце. Кроме света и определенных веществ, в числе возможных внешних раздражителей следует назвать температуру, электрический ток, силу тяжести, воду, прикосновения и ранения. В этих случаях говорят о термо-, гальвано-, гео-, гидро-, тигмо- и травматотаксисах. Но более детальное рассмотрение свободных перемещений, обусловленных этими таксисами, выходит за рамки этой книги.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'