6.1. Ветроопыление (Анемофилия)
* (В экологии опыления используются такие суффиксы, как филия и гамия (фильный и гамный). Поскольку гаметы как таковые не включаются в процесс переноса пыльцы, следует избегать употребления суффикса гамия и пользоваться им только при описании процесса оплодотворения. Термины "автогамия", "ксеногамия" еще допустимы. Термины "зоогамия" или "зоидиогамия" относятся к оплодотворению подвижными гаметами, а не к переносу пыльцы животными.)
Ветроопыление - преобладающий тип абиотического опыления включает в себя, вероятно, 98 % всех известных случаев и широко распространено в таких семействах, как Gramineae, Cyperaceae, Juncaceae, в различных порядках Amentiferae и Urticales. Во многих семействах энтомофильных растений имеется небольшое число анемофильных представителей, к которым относятся виды Thalictrum, Ambrosia, Fraxinus и т. д. Анемофилия у этих растений носит явно вторичный характер, впрочем, как и в семействах однодольных, отмеченных выше, все их ближайшие филогенетические предки - энтомофильны. Как уже было сказано, мы придерживаемся той точки зрения, что у покрытосеменных энтомофилия является более примитивным типом опыления, поэтому если ближайшие предки покрытосеменных были насекомоопыляемыми и если покрытосеменные имеют монофилетическое происхождение, то самые древние представители этого класса тоже были насекомоопыляемыми.
О том, что анемофилия произошла от энтомофилии, свидетельствуют, возможно, наличие реликтовых нектарников в цветках многих анемофилов (Cannabis, Urtica и т. д.; см. Stager, 1902), а также специфические запахи.
Благовещенская (1970) высказала предположение, что первые цветки покрытосеменных не были дифференцированы на анемофильные и энтомофильные и эта дифференциация произошла позднее в процессе эволюции. Возможно, что все это произошло именно так, но тогда возникает вопрос: что было первичным - дифференциация или синдром покрытосемянности? Такой же вопрос возникает в отношении утверждения Мёзе (A. D. J. Meeuse, 1972) о том, что основные анемофилы представлены раздельнополыми покрытосеменными. Однако эту гипотезу мы здесь обсуждать не будем. Во всяком случае, в палеонтологической летописи как анемофильные, так и энтомофильные цветки встречаются очень рано; сережки из среднего эоцена, описанные Крепетом (Crepet, 1974), обладают морфологическими чертами, которые указывают на происхождение от предков с функционировавшими околоцветниками. В частности, старый вопрос о первичности анемофилии у Palmae после исследований в тропиках должен быть решен отрицательно (см. Silberbauer-Gottsberger, 1973; Essig, 1971, 1973).
По сравнению с биотическим опылением, ветроопыление характеризуется меньшей точностью, и для того чтобы оно было успешным, необходима чрезвычайно высокая концентрация пыльцы вблизи источника ее возникновения, поскольку с увеличением расстояния от родительского дерева на каждую единицу длины количество пыльцы уменьшается в 3 раза. У бифункционального ветроопыляемого цветка воспринимающая поверхность обычно сразу же перегружается своей собственной пыльцой, и только полная несовместимость или дихогамия, как правило, могут предотвратить самооплодотворение. Дихогамия обнаружена у Thalictrum, Potamogeton, Plantago и Rumex, у которых она, очевидно, недавнего происхождения. Первоначальная херкогамия малоэффективна, более результативно развитие раздельнополости. Добавим, что эти две функции цветка полностью независимы и, как уже отмечалось, могут объединяться; у ветроопыляемых растений развитие, по-видимому, шло от раздельнополости к двудомности, что и подтверждают полученные данные. Параллельное развитие двух синдромов: ветроопыления и однодомности - может быть прослежено, например, у Thalictrum (Kaplan, Mulcahy, 1971) или Acer (de Jong, 1976). Вопрос о том, что появилось первым - анемофилия или однополость, - не представляет большого интереса. Де Йонг утверждает, что у Acer вначале развилась однополость, и как следствие этого - анемофилия. Однополость может быть связана также с другими функциями растения, например с распространением диаспор, чему очень способствует диморфизм (экзоохория головок с семянками у Xanthium). Морфологически тычиночные и пестичные цветки у анемофилов могут быть совершенно разными.
У некоторых растений наряду с анемофилией наблюдается и энтомофилия, хотя, конечно, эти два способа опыления неравнозначны. Так, анемофильное растение Plantago media более или менее регулярно опыляется насекомыми, даже медоносными пчелами; менее заметный Р lanceolata также посещают насекомые (Clifford, 1962). В цветках Calluna образуется нектар, и эти цветки посещают и опыляют насекомые. Вместе с тем большое количество пыльцы Calluna разносится также ветром, поэтому дополнительное опыление с помощью ветра неизбежно. Большое количество пыльцы у анемофильных растений привлекает насекомых (сборщиков пыльцы), для части которых, например для определенных представителей семейства журчалок (Syrphidae), пыльца составляет существенную часть диеты. Однако следует отметить, что пыльцу некоторых анемофилов пчелы метаболизировать не могут и все же часто взятки пчел-сборщиц содержат или иногда полностью состоят из пыльцы анемофильных растений (см. Sharma, 1970). Поскольку строение цветка анемофильного растения приспособлено к другому способу опыления, роль насекомых в опылении этих растений в лучшем случае весьма сомнительна. Посещение насекомыми "однополых" цветков вообще не имеет смысла. Пойяр (Pojar, 1973), который не касался вопроса о роли шмелей в опылении анемофильных растений, отметил, что пригодность в пищу такой пыльцы может способствовать сохранению ценной популяции опылителей, которые, возможно, не смогли бы существовать на пище, предоставляемой только облигатными энтомофилами (ср. гл. 15). Образование огромного количества пыльцы у анемофильных растений (так, например, кукуруза дает 50 кг пыльцы с 1 га) может показаться своего рода расточительством. Однако эту величину всегда следует сравнивать с непроизводительными затратами на выделение нектара. Продукция пыльцы, исчисляемая от 6 до 80 кг на 1 га (Fiegri, Iversen, 1975), соответствует продукции 15-600 кг меда (Maurizio, Grafl, 1969).
Относительное значение анемофильного и энтомофильного способов опыления меняется не только в зависимости от видовой принадлежности, но также и в зависимости от принадлежности к различным надвидовым таксонам и также в зависимости от характера местообитания, главным образом от наличия в нем насекомых, поскольку ветер практически дует повсюду.
При условии случайного рассеивания пыльцевых зерен из общего количества произведенной пыльцы (N) только некоторое число пыльцевых зерен (n) оказывается "эффективным", т. е. осуществившим опыление; это число является функцией площади поверхностей рылец а и общей площади поверхности цветка А:
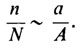
При условии случайного рассеивания пыльцевых зерен из общего количества произведенной пыльцы (N) только некоторое число пыльцевых зерен (n) оказывается 'эффективным', т. е. осуществившим опыление; это число является функцией площади поверхностей рылец а и общей площади поверхности цветка А
Анемофилия и синдром анемофилии могут быть представлены как серия компенсаторных механизмов. Простейший из них - увеличение количества продуцируемой пыльцы N. Хотя каждый может легко убедиться в огромном количестве продуцируемой пыльцы, встряхивая, например, цветущую ветвь Corylus, строгую количественную оценку в данном случае дать трудно, поскольку чрезвычайно сложно выбрать соответствующую единицу счета. Наиболее подходящей единицей может быть число образовавшихся пыльцевых зерен на один семязачаток; но для растений с однополыми цветками данные такого рода получить очень трудно. О трудностях, связанных с получением правильной оценки, свидетельствует табл. 1, заимствованная из работы Поля (Pohl, 1937b). По данным этого автора, число пыльцевых зерен у Corylus варьирует от 3,5 до 0,25 млн. Поль (Pohl, 1937b) определил число пыльцевых зерен на один семязачаток для нескольких видов; его список, за исключением культурных растений, у которых число семязачатков может изменяться под действием искусственного отбора, приводится в табл. 1. Так как пока мы располагаем очень небольшим количеством сравнимых данных, можно сделать только предварительный вывод о том, что у анемофилов (и тычиночных цветков) число пыльцевых зерен, рассчитанных на один семязачаток, в целом превышает число пыльцевых зерен, образующихся в нектарных цветках, но что их диапазоны в значительной мере перекрываются. Обычно у анемофилов число пыльников в одном цветке невелико, значительно больше их обнаружено у энтомофилов или хироптерофилов, у которых аттрактантом является пыльца.
| Вид | Способ опыления | Число пыльцевых зерен, тыс. |
|---|---|---|
| Corylus avellana | Анемофилия | 2549 |
| Fagus silvatica | Анемофилия | 637 |
| Aesculus hippocastanum* | Энтомофилия | 452 |
| Acer pseudoplatanus | Энтомофилия | 94 |
| Tilia cordata | Энтомофилия | 44 |
| Plantago lanceolata | Энтомофилия | 15 |
| Sanguisorba officinalis | Энтомофилия | 11 |
| Betula verrucosa | Анемофилия | 7 |
| Polygonum bistorta | Энтомофилия | 6 |
* (Aesculus по числу пыльцевых зерен (452) является исключением среди энтомофилов и должен быть рассмотрен в связи с синдромом распространения (очень крупные семена, производимые в очень незначительном количестве).)
Эффективная поверхность рылец (а) входит в формулу для вычисления эффективности анемофильного опыления, и, как было обнаружено, у многих анемофилов рыльца сильно увеличены, как, например, у Plantago, Corylus, представителей семейств Juncaсеае и Gramineae. Перистые рыльца, например рыльца у злаков, представляют собой особый случай, поскольку эффективность такого рыльца в улавливании разносимой ветром пыльцы гораздо больше, чем, например, эффективность листа такой же площади (Gregory, 1957); следовательно, такие рыльца обеспечивают большую эффективность в улавливании пыльцы. Чрезвычайно благоприятной в аэродинамическом отношении является также щетковидная форма соцветий Турка. Для удержания пыльцевых зерен рыльца анемофилов должны быть, так сказать, "действенными": чаще всего они бывают липкими.
Общая площадь поверхности цветка А входит как обратный коэффициент при вычислении эффективности анемофильного опыления; эта площадь имеет тенденцию к уменьшению различными способами, чтобы обеспечить свободный доступ к рыльцу. Процесс редукции в большей степени затрагивает околоцветник, который у анемофилов редуцирован всегда, а часто практически отсутствует или опадает. Цвет и запах не играют существенной роли в комплексе признаков, характерных для анемофильного типа опыления: они часто совсем отсутствуют. Прицветники и околоцветники часто бывают зелеными или от темно-коричневых до красноватых. Полагают, что темно-красный цвет играет важную роль в создании определенных температурных условий в цветке, особенно в почке. Некоторые анемофилы все же обладают очень красочными околоцветниками, например Plantago media (щетковидный тип цветка), или имеют определенный запах. Приведенные факты свидетельствуют о том, что у этих растений анемофилия развилась из энтомофилии сравнительно недавно.
Дальнейшее уменьшение неэффективной поверхности достигается выносом цветков за пределы листовой массы, например у злаков или Plantago, и, кроме того, редукцией листовой поверхности (Junci genuini sensu Buchenau); у анемофильных деревьев цветение происходит до появления первых листьев.
Эффективность рылец в улавливании пыльцы увеличивается благодаря выносу их наружу и выше окружающих прицветников, околоцветников и т. д. Однако весьма существенно, что и пыльники выносятся подобным же образом, обеспечивая возможность переноса пыльцы воздушными потоками. Анемофильные цветки часто характеризуются длинными тычиночными нитями, которые выносят пыльники над околоцветником и т. д., как у злаков, представителей Cyperaceae, Thalictrum, Plantago и др. встречаются даже "взрывающиеся" пыльники, например у Ricinus. У некоторых представителей Urticaceae в почках тычиночные нити находятся под сильным давлением и мгновенно высвобождаются при раскрывании цветка и пыльника (см. Mosebach, 1932). В результате этого пыльца чрезвычайно активно выбрасывается в воздух, попадая в более сильные воздушные потоки на некотором расстоянии от растения. Такой способ рассеивания пыльцы можно сравнить со способом рассеивания спор у многих криптогамных растений, особенно у высших мхов.
У ветроопыляемых растений встречается и другой - противоположный - механизм, а именно механизм для удержания пыльцевых зерен, хотя некоторые из подобных примеров, как оказалось, могут быть неверно истолкованы. Например, пыльцевые зерна у Betula или Corylus вначале активно задерживаются в сережках (соцветиях) между близко расположенными чешуйками. При раскачивании сережек ветром между чешуйками образуется отверстие, через которое высыпается пыльца. Это легко увидеть, если принести домой молодые сережки и дать им возможность созреть. Пыльца практически не высыпается из сережек до тех пор, пока они не начнут хотя бы слабо двигаться. Более примитивный механизм удержания пыльцы можно наблюдать у Pinus или У Triglochin, у которых пыльцевые зерна удерживаются между редкими чешуеподобными выступами жесткого прямостоячего мужского стробила до тех пор, пока влажность не уменьшится, а ветер не усилится до того, что сможет выдуть (и унести) пыльцу. Прямостоячие, сережкоподобные цветки встречаются также и у энтомофильных растений (Salix, тропических видов Castanea и Quercus); это, возможно, свидетельствует (поскольку энтомофилия является более примитивным типом опыления) о более низкой стадии развития по сравнению со стадией, на которой находятся родственные анемофильные виды растений с висячими сережками (Populus, виды Quercus умеренной зоны).
До наступления благоприятной (теплой и сухой) погоды пыльники обычно не раскрываются. Теоретически для анемофильных растений влажный климат очень неблагоприятен. В дождливую погоду пыльца быстро намокает. Пыльники лопаются, как правило, с наступлением теплой погоды независимо от того, связано ли это потепление с суточными колебаниями температуры или с более существенными сезонными изменениями погоды. У злаков пыльца высвобождается в результате взрывания пыльников (Пономарев, 1966). У различных видов вылет пыльцы происходит в разное время суток, что ограничивает возможности скрещивания. По данным Панди и др. (Pande et al., 1972), в Индии автогамные цветочки ячменя бывают открытыми примерно в течение часа, при этом пыльники лопаются в течение первой половины этого часа, а рыльца остаются восприимчивыми в течение 6 ч. Очень способствуют распространению пыльцы вихревые воздушные потоки, устанавливающиеся в сухую солнечную погоду, и особенно весной, когда земля еще почти голая.
На жизнеспособность пыльцы влияют внешние факторы, в том числе ультрафиолетовое излучение. Указанные в литературе периоды жизнеспособности пыльцевых зерен некоторых растений столь кратки, что делать какие-либо выводы из этих данных следует очень осторожно. Во всяком случае, чем короче период жизнеспособности пыльцы, тем меньше возможностей, для гибридизации (см. Тикменев, 1974; Пономарев, Прокундин, 1975). Следовательно, короткие периоды жизнеспособности пыльцы, как и короткие почти взрывные периоды цветения, можно рассматривать в качестве своего рода компенсаторного механизма.
У анемофильных растений очень важным свойством пыльцевых зерен является их летучесть, которая увеличивается с уменьшением их размеров. Пыльцевые зерна анемофилов принадлежат к классу пыльцы с "типичным" диаметром 20-30 (60) мкм; такие же мелкие или даже еще мельче пыльцевые зерна у многих энтомофилов (общий диапазон размеров пыльцы покрытосеменных примерно 10-300 мкм). Предполагают, что в силу физических законов самые мелкие пыльцевые зерна могут иметь сильную тенденцию к склеиванию, поэтому, возможно, у анемофильных растений и не бывает совсем мелких зерен. Однако существование гораздо более мелких анемохорных микроспор у других групп растений делает это предположение менее вероятным.
Проблема летучести некоторых относительно больших пыльцевых зерен у хвойных решается путем образования в пыльцевом зерне одного или нескольких воздушных мешков, создающих больший объем и большую поверхность без заметного увеличения веса. В результате крупные (50-150 мкм) и тяжелые (30-300*10-6мг) пыльцевые зерна Pinus принадлежат к наиболее эффективным в транспортном отношении. Большие количества пыльцы сосны были найдены за сотни километров от ближайших лесов. По-видимому, очень большие пыльцевые зерна анемофильных растений (без воздушных мешков) имеют сравнительно низкую плотность (Pohl, 1937а), что также обеспечивает им большую летучесть. Следует отметить, что вес пыльцевых зерен (см. Pohl, 1937а; Dyakowska, Zurzycki, 1959) зависит также от влажности воздуха, параметра весьма непостоянного. Во всяком случае, при подсыхании некоторые сферические пыльцевые зерна приобретают сплюснутую форму, имеющую наилучшие аэродинамические свойства.
Средняя скорость падения пыльцы в спокойном воздухе равна нескольким сантиметрам в секунду (Rempe, 1937).
Эффективность переноса пыльцы анемофилов усиливается тем, что их зерна, гладкие и сухие, не слипаются друг с другом, а распространяются либо поодиночке, либо очень маленькими группами (Rempe, 1937). Поверхность пыльцевых зерен энтомофильных растений скульптурированная и липкая, поэтому зерна склеиваются вместе. Типичные анемофильные зерна обнаружены у представителей семейств Betulaceae и Gramineae. На примере нескольких семейств, в том числе Compositae, показано, что параллельно с эволюцией анемофилии шло развитие гладкой и сухой поверхности и тонкой экзины у пыльцевых зерен (Wodehouse, 1935). Обычно пыльцевые зерна у растений из семейства Compositae сильно скульптурированы и липкие; пыльцевые зерна растений из анемофильных родов Ambrosia или Artemisia - гладкие и сухие. Последний случай представляет собой реверсию, так как пыльцевые зерна примитивных энтомофильных покрытосеменных были с довольно гладкой поверхностью. Сильно скульптурированные оболочки пыльцы высоко развитых в эволюционном отношении энтомофилов являются вторичным признаком.
Наличие небольших количеств масел на поверхности пыльцы многих анемофильных растений [например, у Plantago spp. (Knoll, 1930b); у злаков (Pohl, 1929)] интерпретировали как доказательство их происхождения от энтомофильных предков. Однако проведенные в последнее время исследования роли оболочки пыльцевого зерна в процессах узнавания между пыльцевым зерном и поверхностью рыльца и в процессе прорастания пыльцы не позволяют рассматривать эти масла как инертное клееподобное вещество. Возможно, нет ни одного вида пыльцы покрытосеменных, у которого экзина была бы полностью лишена масел (Knoll, цит., Pohl, 1929), хотя у некоторых растений количество его очень незначительно, например у Corylus или Betula. Кроме того, оно может переходить с наружной поверхности оболочки во внутреннее пространство (ср. Hesse, 1979 и ранее цит.). У некоторых видов при переходе от энтомофилии к анемофилии количество масел меняется [например, у Atisma plantago (Daumann, 1966)]. Хвойные, как первичные анемофилы, не имеют масел на поверхности пыльцевых зерен.
Свободное и равномерное рассеивание пыльцевых зерен анемофилов легко продемонстрировать, оставив несколько веток с сережками Betulaceae или метелок злаков в вазе в комнате без сквозняка; через некоторое время пыльца ровным слоем покроет стол (стол должен быть темным). Кнолль использовал для наблюдений за переносом пыльцы по воздуху специальный прибор (Knoll, 1930b), состоявший из широкой стеклянной трубки, в которой пыльца могла свободно падать, и приспособления для подачи пыльцы в трубку, но так, чтобы при этом не создавались вихревые токи воздуха. На предметных стеклах, располагавшихся на дне трубки, получались "картины падения пыльцы", а о количестве масел на поверхности пыльцевых зерен можно было судить по следам, оставленным пыльцевыми зернами на стеклах, на которых они собирались (Daumann, 1966).
Необходимым условием для эффективного ветроопыления является ветер. Это было установлено как для разреженной растительности открытых мест, так и для верхнего яруса сомкнутых, многоярусных типов растительности. В густых лесах ветер бывает редко, а если и бывает, то он так слаб, что анемофилия трудноосуществима. В этих условиях даже довольно мелкие пыльцевые зерна оказываются слишком тяжелыми и падают слишком быстро, не успевая распространиться на большие расстояния. Семириков и Глотов (1971) установили, что максимальное расстояние, на которое может рассеяться пыльца Quercus petraea в лесу, - 80 м, а Коски (Koski, 1970) вычислил, что в Финляндии "возможное расстояние для опыления" в сосновом лесу составляет 53 м. Сходные данные получены в различных палинологических исследованиях (см. Fægri, Iversen, 1975).
Корреляция между спектром опыления (процентное соотношение различных агентов опыления, активных в данной местности) и более или менее ветреным климатом была выявлена в старой работе Кнута. В Германии 21% растений - анемофилы, в северном прибрежном районе 27 %; на очень ветреных островах Северного моря 36%; на плоских песчаных отмелях 47%. Спектры опыления могут оказаться для различных районов одинаковыми (такие примеры имеются), что свидетельствует о пригодности и эффективности различных агентов опыления в условиях разнообразного климата и фауны. Однако истинная ценность таких данных довольно ограниченна, так как помимо климатических воздействий имеют значение и другие факторы (Kugler, 1975).
Другим условием для эффективного осуществления анемофилии является достаточная скученность рассматриваемых видов. Небольшие популяции всегда находятся в невыгодном положении для опыления, но особенно это касается анемофилии, эффективность которой зависит от определенной плотности распространяющейся пыльцы. Вместе с тем даже у изолированных экземпляров двудомных анемофильных растений небольшое количество семян все же завязывается (если двудомность абсолютна).
Вопрос об образовании и рассеивании пыльцы анемофильных растений представляет собой фундаментальный вопрос пыльцевого анализа и подробно изложен в соответствующих руководствах (например, Fægri, Ivensen, цит.). Обычно скорость вихревых токов воздуха больше скорости падения пыльцевых зерен, и постепенное снижение содержания пыльцы в воздухе обусловлено скорее ее диффузией, чем падением.
Многие исследователи пытались определить расстояние, на которое может уноситься пыльца с помощью ветра. Можно считать доказанным, что даже в нескольких десятках километров от леса в воздухе рассеяно достаточно пыльцы, чтобы произошло по крайней мере незначительное число оплодотворений. Так, Хессельман (Hesselmann, 1919), проводя исследования на побережье Ботнического залива, обнаружил, например, 700 пыльцевых зерен на 1 см2 как Betula (хорошая летучесть), так и Picea (плохая летучесть); при этом скорость падения зерен была соответственно 1,5 и 7,9 см/с. Другой интересный пример - перенос больших количеств пыльцы на значительное расстояние в сильном воздушном потоке (см. Rempe, 1937). Такой перенос пыльцы на большое расстояние очень важен для опыления и завязывания семян у изолированных растений, а также для переноса на значительные расстояния различных генов и получения локальных мутаций. Рекордное расстояние, на которое может переноситься пыльца некоторых анемофилов, варьирует от 150 до 1300 км (оба примера касаются хвойных), но подобные данные не имеют большого значения. Так например, пыльцевое зерно, поднявшись довольно высоко в воздух, может упасть куда угодно, но при этом оказаться нежизнеспособным. Большая часть пыльцевых зерен падает ночью, из чего можно сделать заключение, что среднее максимальное расстояние, на которое переносится пыльца, составляет примерно 50 км.
И наконец, для эффективного опыления большое значение имеет площадь массового оседания пыльцы. Она очень сильно изменяется в зависимости от изменения таких условий, как ветер, высота растительности, продолжительность периода высвобождения пыльцы и т. д., но в среднем эта величина варьирует от нескольких метров до нескольких сотен метров, а при исключительных обстоятельствах - до километра.
У анемофилов число семязачатков в одном цветке обычно очень незначительно по сравнению с большим количеством пыльцы; часто семязачаток вообще бывает один, поэтому в каждом цветке образуется только одно семя. Это часто резко отличается от того, что мы наблюдаем у родственных энтомофильных таксонов, например у рода Bocconia из семейства Papaveraceae (ср. Pohl, 1929). Возможно, такой контраст связан с характером пыльцы; когда одно липкое пыльцевое зерно типичного энтомофильного растения попадает на рыльце, то там в это же время уже оказываются прилипшими обычно несколько пыльцевых зерен, поэтому имеется возможность для оплодотворения нескольких семязачатков. Такой возможности нет у анемофилов в случае равномерного рассеивания гладких пыльцевых зерен, попадающих на рыльце поодиночке. Таким образом, редукцию числа семязачатков следует рассматривать в связи с синдромом рассеивания пыльцы.
Так как очень малое число семян на один пестик рассматривается обычно как производный признак, то это может указывать на то, что и анемофилия также носит производный характер.
Синдром анемофилии. Цветки однополые, распускаются до появления первых листьев или же располагаются над массой листьев, околоцветник невзрачный, слабо выражен или совсем отсутствует; аттрактанты отсутствуют; пыльники и рыльца заметно вынесены над остальными частями цветка; пыльцевые зерна мелкие, гладкие, сухие, образуются в больших количествах; часто развивается механизм для улавливания пыльцы; число семязачатков сильно редуцировано.
Концепция ветроопыления обычно предполагает, что ветер, воздушные потоки, переносят пыльцевые зерна на некоторое расстояние, по крайней мере от одного цветка к другому. Однако колебля цветки, ветер легко может вызвать автогамию в гомогамных гермафродитных цветках. Такая автогамия играет также важную роль при выращивании некоторых сельскохозяйственных культур, например капока (Ceiba pentandra).
Кроме того, ветер может играть косвенную роль в энтомофильном опылении путем привлечения внимания опылителей-разведчиков к цветкам в случаях, подобных микромелиттофилии или Oncidium которые рассматриваются в разд. 8.6.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'