5.3. Приобретенный (индуцированный) иммунитет
Успехи, достигнутые в области медицинской иммунологии, послужили стимулом к поиску аналогичных явлений в фитоиммунологии. Исследовали природу индуцированной устойчивости у растений и возможности ее практического применения. Но когда в них не обнаружили антител и стало ясно глубокое отличие в природе иммунитета животных и растений, изучение этого вопроса затормозилось. Однако в последние 20 лет интерес к проблеме приобретенного (индуцированного) иммунитета вновь повысился. Это было связано с несколькими причинами. 1) Устойчивость выведенных селекционных сортов растений недолговечна и рано или поздно они начинают поражаться вновь появившимися расами паразитов. 2) Химическая защита растений довольно дорога, а грибы приспосабливаются к фунгицидам, которые становятся неэффективными. Кроме того, часто неконтролируемое и безграмотное использование фунгицидов в больших количествах загрязняет окружающую среду и вредно отзывается на здоровье животных и человека. 3) Есть возможность регулировать синтез защитных веществ у восприимчивых растений внешними воздействиями.
Устойчивость растений к заражению может повышаться как в результате перенесенного заболевания, так и в результате вакцинации.
О механизме повышения устойчивости растений после перенесенного вирусного заболевания мы уже говорили. Разберем теперь приемы вакцинации.
В настоящее время широко используется вакцинация ослабленными штаммами вирусов. Наибольшие успехи достигнуты при вакцинации томатов вирусом табачной мозаики для борьбы с мозаичным заболеванием томатов в условиях закрытого грунта. Опрыскивание растений ослабленной культурой ВТМ повышает урожай и не сказывается на качестве плодов. Такого рода вакцинация успешно осуществляется в Голландии, Швеции, Дании, Англии, Канаде, США, Японии, а также в нашей стране.
Имеются работы, указывающие на возможность вакцинации и других культур. Так, слабопатогенный штамм ВТМ S7 использовался для защиты томатов от столбура (микоплазмы), огурца - от вируса зеленой крапчатой мозаики. Показано, что картофель, вакцинированный вирусом X, не заражался фитофторозом, а также другими вирусами. Защитные свойства вакцинации сохранялись при этом в течение ряда лет.
Этот метод, однако, должен использоваться с осторожностью, поскольку никогда нельзя исключить появления в популяции вакцинного штамма агрессивных форм, которые поразят устойчивые сорта. Поэтому он используется пока лишь в условиях теплиц.
Вакцинацию растений можно осуществлять также патогенными грибами. Так, для иммунизации огурцов использовали ослабленные культуры (сохранявшиеся в чистой культуре 14 лет) листового и стеблевого штаммов возбудителя аскохитоза Ascochyta cucumeris. Эти штаммы задерживали проявление симптомов и развитие заболевания, в то время как непатогенные штаммы гриба не обладали вакцинирующим действием.
В других опытах первый настоящий лист растений огурца заражали патогеном Colletotrichnm lagenarium, вызывающим антракноз. Через 4 дня у организма развивалась системная устойчивость, т. е. все выше расположенные, а также не раскрывшиеся листья становились устойчивыми к заражению этим и другими паразитами. При этом обнаружена лигнификация тканей прямо под аппрессорием патогена. В другом случае использовали для иммунизации растений горшечного табака конидии Peronospora tabacina, которые помещались в почву вокруг стебля. В листьях развивалась системная устойчивость. Эти приемы вакцинации, как и в случае с вирусами, также должны использоваться с осторожностью, поскольку не исключена возможность заражения растений.
В качестве вакцины в настоящее время используются также индукторы паразита. Например, из мицелия грибов Phytophthora infestans и Fusarium culmorum выделен липогликопротеидный комплекс (ЛГП) - биогенный индуктор, который в концентрации 0,0005% вызывает повышение устойчивости картофеля, томатов и сахарной свеклы к различным болезням. Он обладает системным действием: при обработке клубней картофеля и семян томатов и сахарной свеклы повышенную устойчивость к болезням приобретали растения, выросшие в полевых условиях. Их урожайность повышалась на 10 - 30%. Действие этого индуктора основано не на фунгитоксичности, а на повышении естественного свойства устойчивости. Предполагается, что одним из механизмов действия индуктора является вызываемая им системная сенсибилизация растений. Под влиянием сенсибилизирующих доз ЛГП клетки клубня картофеля и других растений приобретают отдельные черты ультраструктуры, характерные для специализированных терпеноидогенных клеток, с которыми связан биосинтез терпеноидов и фенолов. А фитоалексины картофеля и томатов, как мы уже отмечали выше, принадлежат к числу сесквитерненоидов. Сенсибилизированные клетки, таким образом, уже подготовлены для массового синтеза фитоалексинов, которые при встрече с патогеном быстро образуются и накапливаются.
Таким образом, сенсибилизация связана с функциональной перестройкой клеток клубня в сторону повышения их биосинтетических возможностей, и в первую очередь, с образованием специализированных защитных веществ: терненоидных фитоалексинов, суберина. Возрастают при этом и энергетические возможности сенсибилизированных клеток (увеличивается число митохондрий, исчезает крахмал из амилопластов, используемый в качестве энергетического субстрата).
Основным недостатком этого приема иммунизации растений является дороговизна получения индуктора. Поэтому перед исследователями стоит задача совершенствования технологии их производства.
Химическая иммунизация. Этот прием вакцинации подразумевает обработку растений химическими веществами, которые регулируют протекание защитных реакций. Технологически этот метод прост (растения опрыскивают растворами веществ или замачивают в них семена), перспективен, поскольку можно получить много соединений, влияющих на метаболизм и не обладающих токсическим действием на человека и окружающую среду.
В качестве препаратов-иммунизаторов используют стимуляторы роста, фенолы, макро- и микроэлементы и пр. Они обладают системным действием, и, не оказывая влияния на патоген, изменяют метаболизм растения-хозяина, повышая его устойчивость. Их называют иногда веществами внутри растительного действия.
Каковы механизмы повышения устойчивости растений под влиянием химических иммунизаторов? Например, при опрыскивании водной эмульсией полиакриловой кислоты на поверхности растения образуется пленка, которая препятствует проникновению ростков ржавчины; ауксины повышают устойчивость клеточной стенки к пектолитическим ферментам патогена; гербициды диаллат на пшенице и дихлорциклопропан на рисе способствуют активизации фенольного метаболизма и окислительных процессов, усиливают синтез лигнина и способствуют увеличению прочности клеточных стенок; такие соединения, как антибиотик хлорамфеникол, гербицид симазин, ретарданты индуцируют синтез фитоалексинов; гербициды, ретарданты обладают сенсибилизирующим действием, т. е. подготавливают растение к заражению.
В последние годы в связи с достижениями в области биотехнологии для повышения устойчивости растений к заболеваниям начали использовать методы клеточной инженерии. В частности, для получения растений, устойчивых к грибам или фитонатогенным бактериям, используется следующая последовательность приемов, представленная на схеме (рис. 20). Получают растения-регенеранты, проверяют их на устойчивость и выделяют линии, которые значительно превосходят исходный сорт по устойчивости к бактерии или грибу.
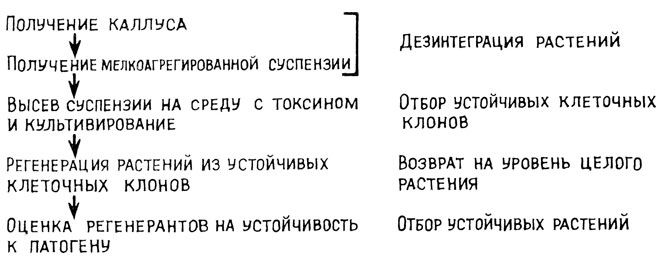
Рис. 20. Схема получения устойчивых к патогенам растений методом клеточной инженерии
Широко используется также метод верхушечной меристемы для оздоровления растений от вирусной инфекции. Для этого выделяют меристему, свободную от вируса, получают из нее каллусную ткань, а затем растение-регенерант, свободное от вируса, которое можно высаживать в почву.
Таким образом, в основе фитоиммунитета лежит множество защитных реакций. Но все они отражают уже завершающую фазу взаимоотношений двух партнеров, конечный неспецифический ответ клетки на контакт с несовместимым патогеном. Между тем, будет ли растение устойчиво или восприимчиво, т. е. индуцирование защитных реакций должно осуществляться на начальных этапах взаимодействия с патогенами, когда происходит распознавание партнеров. Именно распознавание "своего" от "чужого" является первичным процессом, тогда как ответные защитные реакции развиваются спустя какое-то время, необходимое для реализации процесса узнавания.
Различают 2 фазы в процессах, развивающихся после встречи растения и патогена: детерминантная - партнеры распознают друг друга, в результате чего определяется, будут ли они совместимыми или нет; экспрессивная - выражает события, продиктованные детерминантной фазой, - быстрое или медленное включение защитных реакций.
Какие продукты распознает растение? Это метаболиты паразита, называемые индукторами или элициторами. Они находятся на поверхности патогена или в составе его выделений. Очевидно, растение в процессе эволюции "научилось" распознавать такие метаболиты паразита и использовать их как триггер для ответного включения всего комплекса защитных реакций. Большинство индукторов представляет собой высокомолекулярные соединения, в составе которых имеются углеводные группы. В очень низких концентрациях (10-5 M) они вызывают некротическую реакцию (реакцию сверхчувствительности) и накопление фитоалексинов.
Хорошо изученным индуктором является липогликонротеидный комплекс, выделенный из клеточного содержимого гриба возбудителя фитофтороза картофеля Phytophthora infestans. О химической природе других индукторов мы уже говорили в разделе о фитоалексинах.
Механизм распознавания до настоящего времени экспериментально еще не изучен. Согласно гипотезе Альберсхейма и Андерсон-Проути [11], индукторы патогена представляют собой как бы антигенную детерминанту для их распознавания рецепторными белками (лектинами) в составе мембран растения-хозяина. Продуктом гена авирулентности патогена должен быть фермент, ответственный за синтез молекулы индуктора, тогда как продуктом гена устойчивости является система, участвующая в образовании рецепторного белка па мембране растения-хозяина. Поскольку ген устойчивости растения и ген авирулентности паразита комплементарны, то происходит взаимодействие их продуктов, что и приводит к включению реакции сверхчувствительности и отторжению паразита.
Считается, что в процессе распознавания вполне вероятен межклеточный контакт партнеров посредством лектин-углеводного взаимодействия. Имеются наблюдения, подтверждающие возможность такого механизма взаимодействия растения-хозяина и патогена.
Так, обнаружено, что гликопептиды клеточной поверхности гриба Phytophthora infestans, содержащие глюкозамин - специфический лиганд для лектина картофеля, - связываются с плазмалеммой, выделенной из паренхимных клеток клубней картофеля. При этом в устойчивой комбинации это связывание сильнее, чем в неустойчивой. Факт взаимодействия плазмалеммы с гликоконъюгатами гриба подтверждается также подавлением гемагглютинирующей активности плазмалеммы гликопептидами патогена.
В плазменной мембране сои обнаружены сайты со способностью активно связывать глюканы клеточной стенки патогенного гриба Phytophthora megasperma. Эти сайты могут играть роль в элицитор-обусловленном защитном ответе. Белок мембран, связывающий глюкан, сохраняет свою связывающую активность после солюбилизации. Гликопротеины из клеточной стенки уредоспор Puccinia graminis tritici селективно связывались с лектино-подобными белками, выделенными из листьев пшеницы.
О действии индукторов (элициторов) на уровне поверхности клетки свидетельствуют также наблюдения, в которых показано, что элициторы ряда патогенов обладают способностью деполяризовать плазмалемму клеток растений. Этот эффект был быстрым, стабильным и обратимым. Обратимость действия элицитора свидетельствует о том, что он не повреждает растительную мембрану.
Большинство известных в настоящее время индукторов являются неспецифическими, т. е. они вызывают защитные реакции у всех сортов растений (устойчивых и неустойчивых к патогену, из которого они выделены) и присутствуют у всех рас гриба. Именно неспецифические индукторы распознаются растением и, очевидно, такой механизм лежит в основе видового иммунитета.
А какова же природа специфической, сортовой устойчивости? Каким образом специфические паразиты препятствуют включению защитных реакций?
По мнению большинства исследователей, специфические патогены научились подавлять неспецифический защитный ответ растений. Так, из мицелия гриба Phytophthora infestans были выделены вещества, которые подавляли реакцию сверхчувствительности у картофеля, вызываемую инфицированием несовместимой расой. Эти вещества, получившие название супрессоров (фактор совместимости), представляют собой низкомолекулярные глюканы со связями β-1,3- и β-1,6- и содержатся лишь в совместимых расах. Эти соединения облегчают распространение несовместимого гриба но тканям растений, индуцируя в них восприимчивость. К числу супрессоров, кроме токсических глюканов, относятся также токсины паразитов и импедины (нетоксичный бактериальный фактор подавляющий защитный механизм растения-хозяина).
Как же действуют супрессоры? Предполагается, что в совместимой комбинации супрессоры, выделяемые патогеном, конкурируют с индуктором за рецепторный участок на плазмалемме растения-хозяина, имея к нему большее сродство, и препятствуют таким путем включению защитных реакций.
Считают, что супрессор имеет 2 активных центра: неспецифическую группировку, которая конкурирует с индуктором за рецепторный участок у хозяина, и специфическую группу, которая изменчива. Поскольку растение в процессе эволюции приобретает новые гены сортовой устойчивости (R), предполагается, что продукты этих новых генов устойчивости связывают супрессор паразита по его вариабельной группировке и как бы "уводят" его с поверхности взаимодействия паразита и хозяина. Индуктор остается вне конкуренции и вновь взаимодействует с рецепторами, индуцируя защитные реакции. Такое растение приобретает сортовую устойчивость к патогену.
В результате мутаций гриба вариабельная часть супрессора (которая связывалась продуктом R-гена) изменяется так, что теряет комплементарность к этому продукту и перестает им связываться. Супрессор вновь оказывается на свободе и начинает вытеснять индуктор в борьбе за рецептор. Так возникает ген вирулентности паразита, способный преодолевать ген устойчивости. Далее процесс повторяется.
Встает вопрос: как же реализуется (или передается) сигнал, ведущий от распознавания к включению защитных реакций, т. е. от детерминантной к экспрессивной фазе? Ответ на этот вопрос в настоящее время отсутствует. В соответствии с гипотезой индуктор - рецептор вслед за связыванием индукторов несовместимой расы патогена рецептором растения через плазмодесмы между плазматической мембраной и ядрами контактирующих клеток и соседних клеток хозяина проходят вторичные мессенджеры, аналогичные циклическим нуклеотидам других организмов. Такие мессенджеры запускают процессы обмена и, в конечном счете, приводят к синтезу и накоплению фитоалексинов и запуску других защитных реакций.
В качестве вторичных мессенджеров могут функционировать эндогенные индукторы. Образование этих индукторов происходит в результате деятельности синтезируемой фитопатогенными грибами эндонолигалактуроназы, которая вызывает распад клеточных стенок растения-хозяина с образованием пектиновых фрагментов (олигосахаридов), действующих как эндогенные индукторы фитоалексинов. В настоящее время эти соединения привлекают внимание исследователей как возможные посредники ответа растения на стресс. Они индуцируют такие естественные защитные механизмы растений, как синтез фитоалексинов, лигнина, ингибиторов протеиназ, богатых гидроксипролином гликопротеинов. Способность олигосахаридов индуцировать защитные реакции растений является одним из аспектов общего свойства этих молекул регулировать генную экспрессию.
Роль возможного вторичного мессенджера могут выполнять также ионы Ca2+. В связи с ролью Ca2+ в преобразовании внешних воздействий в клеточный ответ с помощью кальмодулина - рецептора, образующего интегральную часть Ca2+-мессенджер системы, изучали роль Ca2+ в ускорении синтеза фитоалексинов (ришитина и любимина). Обнаружено, что при обработке тканей клубня картофеля арахидоновой кислотой - индуктором фитоалексинов - ионы Ca2+ стимулировали накопление ришитина и любимина. Добавление ЭГТА, которая связывает Ca2+ клубней, подавляло накопление фитоалексинов. Эти наблюдения позволяют предположить, что ионы Ca2+ каким-то путем могут регулировать первичные процессы взаимодействия паразита с растением-хозяином - процессы распознавания, включающие затем защитную реакцию, т. е. Ca2+ выступает как вторичные мессенджеры.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'