5.2. Индуцированные защитные свойства
Реакция сверхчувствительности. Реакция сверхчувствительности (СВЧ) является универсальной защитной реакцией растений. Она лежит в основе большинства известных в настоящее время случаев проявления фитоиммунитета не только в отношении специфических патогенов, но также и неспецифических. В биологическом смысле реакция СВЧ служит типичным примером отторжения чужеродного начала, происходящего в результате распознавания "несвоего". Внешним выражением реакции СВЧ является быстрая гибель клеток, в которые проникли гифы несовместимого патогена, а затем и гибель гиф. Чем устойчивее растение, тем быстрее гибнут клетки с находящейся в них гифой и тем меньшее число слоев клеток отмирает. Эти отмершие клетки отторгаются четко выраженной плотно упакованной отграничивающей перидермой.
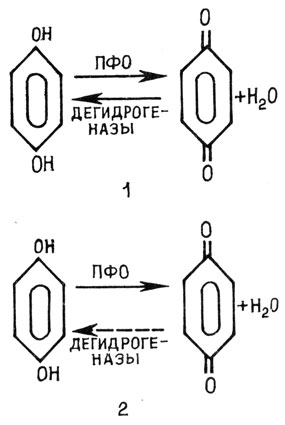
Рис. 18. Схема окисления полифенолов в здоровой (1) и инфицированной (2) клетках
При изучении причин отмирания клеток прежде всего обратили внимание на продукты окисления фенолов с помощью полифенолоксидазы. Разберем этот процесс на примере картофеля, инфицированного Phytophthora infestans. В неповрежденной клетке фенолы и полифенолоксидаза разобщены: первые находятся в вакуоле, вторая - в цитоплазме. Поступление фенолов в цитоплазму и их окисление ферментом ограничено полупроницаемостью тонопласта. При повреждении клетки (внедрение патогена) усиливаются процессы накопления фенолов, а также увеличивается активность окислительных ферментов - полифенолоксидазы и пероксидазы за счет процессов синтеза их белка de novo. Полифенолы концентрируются на внутренней стороне тонопласта, повреждая его и увеличивая проницаемость. В результате фенолы ускоренно поступают в цитоплазму, где бурно окисляются полифенол- оксидазой и пероксидазой. Продукты окисления фенолов - хиноны - подавляют активность дегидрогеназ, и в результате процесс окисления фенолов становится необратимым и сдвинутым в сторону образования хинонов (рис. 18). Последние вызывают серьезные нарушения в структуре клетки, повреждая мембраны лизосом. В результате гидролитические ферменты, содержащиеся в этих структурах, оказываются в цитоплазме, и начинается автолиз, самопереваривание содержимого клетки, в результате чего клетки отмирают. Продукты окисления фенолов легко взаимодействуют с функциональными группировками белков, такими как NH-2, OH-, SH-, и тем самым могут инактивировать белки-ферменты, в том числе гидролазы патогенов, а также повреждать белки мембран. Установлено также, что хиноны являются разобщителями окисления и фосфорилирования.
Хиноны (в частности О-хиноны), образующиеся в ходе окисления, характеризуются слишком коротким периодом жизни, в течение которого только они могут проявлять свои повреждающие свойства. Они быстро окисляются дальше, а продукты глубокого окисления фенолов уже мало токсичны и не ингибируют полностью гриб, а лишь ослабляют его.
Когда были открыты фитоалексины - антибиотические индуцированные вещества растительных клеток, - оказалось, что они также накапливаются в некротизированных клетках. Эти вещества обладают фунгицидной активностью и убивают гриб.
В настоящее время считается общепризнанным, что как хиноны, так и фитоалексины, обнаруженные в некрозе, являются вторичными продуктами. Включение их синтеза осуществляется другими реакциями. В частности, некротизация является триггером для процесса образования фитоалексинов. Например, вызванная несовместимой расой гриба Phytophthora infestans гибель нескольких поверхностных клеток клубня картофеля является сигналом, идущим вглубь ткани, где и начинается синтез фитоалексинов. Последние транспортируются и накапливаются в некротизированных клетках, где находится паразит.
Возникает вопрос: что же в таком случае является первопричиной некротизации тканей?
Японские [12] и русские [1] исследователи установили, что несовместимая раса патогена Phytophthora infestans или его активные метаболиты индуцируют генерацию радикалов кислорода (O 2) тканями картофеля, что и является причиной сверхчувствительной гибели клеток растения. Ускоренная генерация активных форм кислорода наблюдается уже через 1 час после контакта патогена с растением, а максимум их образования совпадает с началом сверхчувствительной гибели клеток картофеля, т. е. через 4 - 6 часов после заражения.
2) тканями картофеля, что и является причиной сверхчувствительной гибели клеток растения. Ускоренная генерация активных форм кислорода наблюдается уже через 1 час после контакта патогена с растением, а максимум их образования совпадает с началом сверхчувствительной гибели клеток картофеля, т. е. через 4 - 6 часов после заражения.
Ферменты супероксиддисмутаза и каталаза подавляют образование супероксидрадикала и одновременно некрозов и фитоалексинов. Подавление образования супероксидрадикала вызывал также п-хлормеркурийбензоат, причем как таковой, так и связанный с декстраном М.в. 70 000Д. Этот декстран не проникает в клетку, поэтому можно предположить, что образование O

2 происходит непосредственно на плазматической мембране. Образующиеся в несовместимой комбинации на плазматической мембране растения супероксиданион, как и другие активные формы кислорода (свободные радикалы), индуцируют процессы переокисления липидов мембран, являющиеся основным механизмом повреждения мембран клеток. Действительно, как показывают наблюдения, еще до появления некрозов в тканях активируется липоксигеназа (фермент, катализирующий окисление ненасыщенной связи в молекуле жирной кислоты в насыщенную) и уменьшается содержание ненасыщенных жирных кислот. Накапливаются продукты переокисления липидов - липофусциниодобный пигмент и малоновый диальдегид (продукт взаимодействия тиобарбитуровой кислоты с перекисями липидов), т. е. наблюдается переокисление липидов мембран. Если подавить активность липоксигеназы антиоксидантами (витамин E), то некроз не образуется. Переокисление липидов приводит к нарушению липид-белковых взаимодействий в мембране, повреждению белков мембран и, как следствие этого, наступает гибель клетки. Участие процессов переокисления липидов мембран клетки в реакции сверхчувствительности доказано для несовместимой комбинации картофель+Phytophthora infestans, вигны+вирус мозаики огурца, котиледонов тыквы+бактерия Pseudomonas syringae pisi.
Таким образом, индуцируемые свободными радикалами процессы переокисления липидов и связанные с ними повреждения мембран клеток являются одной из первопричин отмирания клеток растений при реакции сверхчувствительности.
В последние годы появился ряд работ, в которых, однако, не подтверждается роль активных форм кислорода в реакции сверхчувствительности. В частности, обнаружено, что гибель несовместимой расы бактерии Pseudomonas syringae Van Hall в процессе сверхчувствительной реакции в листьях табака не связана с образованием O 2. Точно так же не подтверждена роль синглетного кислорода 1O2 в сверхчувствительной гибели суспензии клеток табака, инокулированных несовместимым патогеном бактерией Pseudomonas syringae pv. pisi.
2. Точно так же не подтверждена роль синглетного кислорода 1O2 в сверхчувствительной гибели суспензии клеток табака, инокулированных несовместимым патогеном бактерией Pseudomonas syringae pv. pisi.
Как показывают исследования Морей и Осман, жидкость, в которой прорастали зооспоры гриба Phytophthora infestans, индуцировала выделение дисками клубней и высечками листьев картофеля редуцирующих веществ, которые, как и радикал O2, восстанавливали цитохром с (именно по активности восстановления цитохрома с и определяют содержание этого радикала). В то же время радикал O2 обнаруживался в этих условиях в очень небольших количествах. Авторы считают, что эти наблюдения не согласуются с гипотезой о включении супероксиданиона в реакцию устойчивости картофеля к Ph. infestans [13].
Фитоалексины. Фитоалексины (ФА) - низкомолекулярные антимикробные соединения, которые синтезируются и аккумулируются в растениях после контакта с микроорганизмами. Они обладают широким спектром антибиотического действия, подавляя рост различных фитонатогенов. Кроме того, они оказывают токсическое действие на организм млекопитающих. ФА были открыты около 45 лет назад, к настоящему времени изучено достаточно много веществ такой природы.
По своим химическим свойствам ФА принадлежат к различным классам соединений. Например, ФА семейства бобовых (пизатин, фазеоллин, медикарпин и др.) относятся к изофлавоноидам; семейства пасленовых (ришитин, любимин, кансидиол) - к сесквитерпеноидам; ФА сложноцветных - полиацетилены (рис. 19). Иногда у одного растения обнаруживается несколько ФА, это помогает ему успешнее бороться с патогенами, поскольку последние легко приобретают нечувствительность к антибиотикам, а привыкнуть к нескольким фитоалексинам гораздо труднее, чем к одному. Кроме того, различные фитоалексины часто обладают разным механизмом повреждающего действия на патоген.
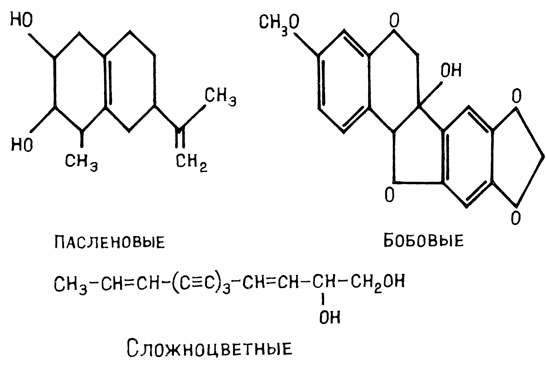
Рис. 19. Разнообразие химической природы фитоалексинов
ФА сосредоточены в инфицированных тканях, и большинство из них локализовано в тканях некроза. По своей биологической активности ФА являются слабыми антибиотиками, т. е. их антибиотическое действие проявляется в концентрациях более высоких, чем антибиотиков. Различные грибы чувствительны к ФА в разной мере. Патогены растения, из которого выделен ФА, значительно более толерантны к его действию, чем непатогены. Например, ЭД50 (доза, ингибирующая патоген на 50%) фазеоллина для непатогенов менее 10 мкг/мл, патогенов - 20 - 50 мкг/мл.
Меньшая чувствительность патогенов к ФА растения-хозяина может объясняться тем, что они содержат ферменты, превращающие ФА в менее фунгитоксичные продукты. Однако не все патогены способны метаболизировать ФА растения-хозяина. Некоторые из них выработали толерантность, нечувствительность к количествам ФА, накапливаемым в зараженных тканях. Совместимые патогены могут тормозить включение защитных реакций растений, в том числе и синтез ФА, выделяя супрессоры, которые блокируют индукторы. Наконец, токсины некоторых патогенов могут повреждать растительные ткани, препятствуя образованию ФА. Так, из культуральной жидкости гриба Mycosphaerella pinodes выделено 2 низкомолекулярных пептида, которые подавляли образование пизатина у гороха.
Образование ФА в тканях растений вызывают самые различные агенты биотической и абиотической природы.
Биогенные индукторы ФА выделены из мицелия патогенов. К ним относятся: полипептиды (мониликолин), белки (индуцирующая активность подавляется проназой), гликопротеиды, ферменты-гликопротеиды - пектинтрансэлиминаза и пектинполигалактуроназа, протеазы (индуктор ФА яблони), белковолипополисахаридный комплекс из мицелия Verticillium albo-atrum, состоящий на 70% из углеводов, липогликопротеид из мицелия Phytophthora infestans, состоящий на 60% из липидов. В составе липидов обнаружены С:20 ненасыщенные жирные кислоты - арахидоновая и эйкозонентаеновая. Именно эти кислоты обладают индукторной активностью.
Кроме того, в клеточных стенках грибов содержатся индукторы ФА, которые выделяются в культуральную среду при росте грибов. Это низко- и высокомолекулярные β-1,3-глюканы, гликопротеиды, β-1,6-глюканы, хитозан - β-1,4-связанный полимер Д-глюкозамина (неацилированное производное хитина).
Обнаружены также т. н. эндогенные индукторы - соединения растительной ткани, освобождающиеся из нее под действием патогена. К ним относятся, например, фрагменты пектинового полисахарида клеточной стенки, освобождаемые из нее пектолитическими ферментами патогена.
Образование ФА может быть индуцировано не только паразитами и их метаболитами, но также некоторыми химикатами: солями тяжелых металлов, ферментными ядами, антибиотиками и т. д. Однако для индукции ФА абиотическими агентами требуется их концентрация на 5 - 10 порядков выше, чем при использовании биотических индукторов, и даже при этом не удается индуцировать ФА в летальных для паразита концентрациях.
Поскольку ФА принадлежат к различным классам химических соединений, они, очевидно, имеют различные механизмы действия. Можно ожидать, что ФА поражают клеточные структуры и звенья обмена, являющиеся общими для живых существ. Неспецифичность токсичности ФА подтверждает широкий спектр организмов, подверженных действию ФА, самые различные физиологические и цитологические изменения, происходящие в обработанных ФА клетках, сравнительно высокие концентрации ФА, необходимые для ингибирования, возможность детоксикации ФА. Предполагается, что ФА, в отличие от антибиотиков и системных фунгицидов, не имеют определенной биохимической мишени, а скорее функционируют как "неснецифические мультисайтовые мембранные антагонисты" [7].
Получены данные, свидетельствующие о том, что токсичность ФА связана с их мембранолитическим действием. Например, установлено, что ФА фасоли фазеоллин вызывает нарушение функций плазмалеммы гриба Rhizoctonia solani, увеличивается вымываемость 14C - соединений из обработанных фитоалексином клеток, нарушается способность мицелия накапливать глюкозу. Любимин и ришитин - ФА картофеля - вызывают деполяризацию мембранного потенциала клеток Nitella. ФА сои глицеоллин подавляет АТФ-зависимый транспорт протона в плазматической мембране и везикулах тонопласта красной свеклы, а также активность Н+АТФазы, т. е. ФА в данном случае оказывает ингибирующее действие на энергизацию транспортных процессов в мембранах плазмы и тонопласта.
ФА повреждают также и митохондриальные мембраны. Например, любимин, ришитин, ипомеамарон (ФА батата) являются разобщителями дыхания и фосфорилирования. Упоминавшийся уже глицеоллин подавляет дыхание и транспорт электронов в ЭТЦ митохондрий сои на участке между НАД. Н-дегидрогеназой и CoQ. Уязвимость клеточных мембран для ФА влечет широкую дисфункцию клеток.
Существует мнение, что ФА принадлежит защитная роль в явлениях видового (неспецифического) иммунитета. Об этом свидетельствует способность различных грибов индуцировать ФА у одного и того же растения, большая чувствительность к ФА непатогенов растений и относительная толерантность к нему специфических патогенов. Вопрос о роли ФА в сортовой устойчивости пока не имеет однозначного ответа, хотя накапливается все больше данных, в которых подтверждается наличие прямой корреляции между продуцированием ФА и устойчивостью сорта. Убедительные данные получены на изогенных линиях растений, отличающихся между собой лишь по гену устойчивости. Оказалось, что линии кукурузы, льна, перца, томатов, сои, содержащие ген устойчивости, образуют ФА вдвое, втрое, а то и в десять раз интенсивнее, чем соответствующие линии, не содержащие гена устойчивости.
В опытах с картофелем установлена роль ФА в вертикальной и горизонтальной устойчивости.
ФА, хотя и являются важным индуцированным защитным механизмом растений, однако их нельзя рассматривать как единственную детерминанту устойчивости, ибо они обнаружены не у всех растений, а их накопление в тканях является результатом предшествующих процессов, которые также могут ограничивать рост патогена.
Дыхание. Роль дыхания в защитных реакциях растений следует рассматривать с 2 позиций: 1) дыхание служит поставщиком исходного пластического материала для различного рода биосинтетических процессов, в том числе для синтеза защитных веществ. Выше отмечалось, что одним из продуктов апотомического (гексозомонофосфатного) пути дыхания является эритрозо-4-фоефат - исходный продукт для синтеза фенолов в клетке; 2) дыхание - процесс, который снабжает клетку энергией за счет окислительного фосфорилирования. Устойчивые к болезням растения способны поддерживать свой энергетический обмен на уровне здоровых растений или даже усиливать его.
Что дает это растению?
Патогены (особенно некротрофы) выделяют мощные токсины, а также экстрацеллюлярные ферменты из группы гидролаз. С помощью токсинов они убивают ткани. Некоторые системы ферментов также разрушают ткани растения-хозяина (клеточную стенку, мембраны, срединную пластинку и т. д.). Кроме того, с помощью ферментов некротрофы превращают клеточное содержимое в удобную для усвоения форму. В результате этих воздействий в ткани растения-хозяина усиливаются гидролитические процессы.
Устойчивые растения противопоставляют гидролитическим процессам, развивающимся в их тканях под влиянием паразитов, процессы синтеза. Синтезируются прежде всего защитные соединения: фитонциды, фитоалексины, белки и пр. Усиление синтетических процессов помогает поддерживать структуру протопласта и жизнедеятельность клетки. Лишь жизнедеятельная клетка способна включать защитные механизмы. Например, реакция сверхчувствительности - это реакция живой, жизнедеятельной клетки. В тканях, отравленных наркотическими веществами (спиртом), реакция сверхчувствительности ингибируется. Она является энергозависимым процессом. Обработка тканей ферментными ядами - ДНФ и NaN3 - подавляет сверхчувствительную реакцию. При этом АТФ (но не АДФ) обращает ингибирующее действие ДНФ и NaN3 на реакцию сверхчувствительности.
Способность устойчивых тканей растений усиливать процессы синтеза лишает также патоген простых соединений, пригодных для питания.
Все синтезы должны быть обеспечены энергией. Усиление же синтетических процессов требует дополнительных ее количеств. Каким образом это достигается? Прежде всего, у устойчивых растений возрастает число энергетически полноценных митохондрий, увеличивается скорость передвижения электронов но электронтранснортной цепи митохондрий, наблюдается также увеличение сопряженности процессов окисления и фосфорилирования (ДК) и коэффициента АДФ/О. В то же время у поражаемых растений наблюдается разобщение процессов окислительного фосфорилирования, снижение коэффициента АДФ/О, подавляется активность мембранно-связанных ферментов.
В основе нарушений функциональной активности митохондрий неустойчивых растений лежат изменения свойств липидных и белковых компонентов их мембран. Так, заражение неустойчивого сорта фасоли возбудителем ризоктониоза грибом Rhizoctonia solani сопровождается т. н. "ожирением" мембран митохондрий, активацией фосфолипазы А и связанной с этим деградацией фосфолипидов и появлением лизоформ. Уменьшается также степень ненасыщенности жирных кислот мембран и увеличивается относительное содержание стеринов, что в конечном счете лишает мембраны присущей им структурной и функциональной пластичности и способствует более легкой повреждаемости.
Наряду с липидным комплексом заболевания вызывают изменения конфирмационных и физико-химических свойств мембранных белков: снижается способность белков связывать липиды, в них обнаруживается большее количество свободных SH-групп, чем в мембранных белках здоровых растений.
Все это свидетельствует о том, что под влиянием инфекции мембранные белки митохондрий претерпевают изменения, сходные с процессами денатурации. У устойчивых форм растений существенных изменений белковых и липидных компонентов мембран митохондрий не наблюдается.
Одной из причин меньшей повреждаемости мембран устойчивых растений по сравнению с неустойчивыми может быть высокое содержание в них а-токоферола, обладающего способностью эффективно связывать свободные радикалы, повреждающие мембраны при патогенезе. Например, в ответ на заражение тканей капусты грибом Botrytis einerea количество а-токоферола в мембранах устойчивого сорта значительно возрастает, тогда как у неустойчивого антирадикальная активность компонентов мембран митохондрий снижается.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'