4.8.2. Фосфорный обмен
Метаболизм фосфата связан с превращением всех органических соединений, а также с энергообменом растений. Фосфор поступает в корневую систему растений в виде высшего окисла PO3-4 и не изменяется, включаясь в органические соединения. Считается, что растения могут потреблять фосфор из растворов, содержащих 1 мкг/л P, но при концентрации его ниже 1 мг/л рост их начинает угнетаться. Концентрации выше 10 мкм P в растворе уже являются токсичными для многих растений.
Изучение фосфорного обмена растений в условиях засоления проводилось многими исследователями.
Установлено, что поглощение фосфата растениями из засоленного субстрата тормозится [16]. Имеются данные о том, что в условиях хлоридного засоления отмечается повышенное содержание фосфора в тканях, а внесение удобрений увеличивает накопление биомассы [5; 16; 53].
Н. В. Жуковская, изучая фосфорный обмен растений в условиях засоления, показала, что хлоридное и сульфатное засоление нарушает фосфатное питание растений. Характер этих нарушений зависит как от особенностей метаболизма растений, так и от типа засоления. В условиях хлоридного засоления у одних видов растений поглощение фосфата из почвы подавляется незначительно, но резко ослабляется передвижение поглощенного фосфата из корней в надземные органы. У других видов подавление поглощения фосфора из почвы выражено значительно сильнее, в то время как ослабление поступления его в растение выражено достаточно слабо. В условиях сульфатного засоления накопление фосфора в тканях растений задерживается. Однако независимо от того, ослабляется поступление фосфора в растение или нет, весь фосфор не используется растением и накапливается в неорганической форме. При этом содержание фосфорорганических соединений в растении снижается.
Наиболее неблагоприятное влияние почвенного засоления на фосфорный обмен растений проявляется на ранних этапах онтогенеза, в период активных ростовых и формообразовательных процессов. По мере приспособления растений к почвенному засолению в онтогенезе способность их к синтезу фосфорорганических соединений повышается. Однако это уже не может компенсировать фосфорный дефицит у растений, начавшийся в начальный период вегетации [16].
К аналогичному заключению приходит В. В. Кабанов, изучавший метаболизм фосфорсодержащих соединений в тканях растений при засолении субстрата хлористым натрием. Автором было исследовано содержание органического и неорганического фосфора в листьях, корнях, стеблях, молодых верхушечных тканях пяти видов высших растений в широком диапазоне концентраций NaCl. Полученные данные показали, что во всех случаях содержание общего и органического фосфора превышало уровень, свойственный дефициту, примерно в 10 раз. Содержание общего и органического фосфора у различных объектов не снижается при высоком содержании в среде NaCl столь резко, чтобы можно было говорить о дефиците фосфора в тканях. Содержание неорганического фосфора практически не коррелирует с содержанием органического.
Изучение содержания моно-, ди- и трифосфорилированных свободных нуклеотидов, ДНК, р-РНК и т-РНК показало, что фосфорный обмен гликофитов обладает очень высокой стабильностью даже при концентрации хлористого натрия, превышающей предел биологической устойчивости вида. При значительном угнетении накопления биомассы листьями гороха (при 0,4% NaCl в субстрате угнетение роста близко к 50%) метаболизм фосфорсодержащих соединений весьма стабилен. В нуклеотидной фракции при избытке NaCl в субстрате замедляется использование ГТФ и УТФ, что можно объяснить снижением интенсивности синтеза белка и замедлением гидролиза крахмала. Кроме того, установлено, что при возрастании содержания NaCl в субстрате увеличивается содержание ДНК в клетках, но наблюдается тенденция к падению РНК [19|.
В реально функционирующих биологических системах обмен фосфорсодержащих соединений представлен на рис. 13.
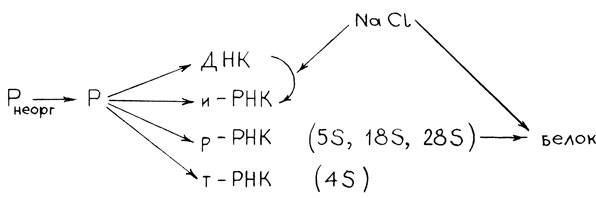
Рис. 13. Обмен фосфорсодержащих соединений в биологических системах при засолении
Особенности действия засоления на все эти звенья синтеза биополимеров, кроме вышеприведенной схемы В. В. Кабанова, в литературе практически не освещены.
Экспериментальные данные свидетельствуют о том, что изменения во многих звеньях фосфорного обмена можно скорее рассматривать как адаптационные, чем как деградационные процессы.
Исследование влияния засоления на содержание низкомолекулярных фосфорсодержащих соединений и включение Р-32 в ткани пшеницы, ячменя, бобов, виноградной лозы показало, что доминирует тенденция к ингибированию включения меченого фосфора и снижению содержания некоторых фракций [16; 31]. Однако в ряде случаев наблюдается существенное повышение в условиях засоления содержания гексозофосфатов, свободных нуклеотидов и включение Р-32 в липиды [53].
Проверка функционирования системы, включающей никотинамидадениндинуклеотид, позволила установить, что содержание восстановленных форм НАДФ-Н2 и НАД-Н2 увеличивается, а окисленных - снижается [68]. У неадаптированных к засолению гликофитов может также существенно нарушаться сопряженность фосфорного обмена и дыхания [21].
Большинство экспериментальных данных свидетельствует о том, что изменения во многих звеньях фосфорного обмена можно рассматривать скорее как адаптационные, чем как деградационные процессы.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'