2.5. Жароустойчивость (действие высоких положительных температур)
Жароустойчивость - способность теплолюбивых растений длительно, а умеренно теплолюбивых кратковременно переносить действие очень высоких температур.
Температурные границы активной жизнедеятельности организмов узки: от 1°С до 45°С. Наиболее благоприятными температурами для жизни большинства высших наземных организмов являются температуры 15 - 30°С. Гораздо шире температурные границы переживания неблагоприятных условий в покоящемся состоянии. Многие семена могут выносить температуру до 100°С в течение ряда часов. Практически они переносят температуру, близкую к температуре абсолютного нуля. Среди бактериальных организмов существуют особые термофильные формы, для которых температурный максимум достигает значений 70 - 80°С, а оптимум - 50 - 65°С. Сухие споры бактерий могут выдерживать температуры порядка 140°С. У высших растений настоящих термофильных форм нет, и скорее можно говорить о термотолерантных формах, способных переносить сравнительно высокие температуры. Они подразделяются на несколько групп:
1) Водные растения с температурным максимумом 38,5 - 41,5°С. Представителем этой группы является лотос желтый (Гавайские острова).
2) Теневые наземные растения - кислица, бальзамины - с температурным максимумом 40,5 - 42,5°С.
3) Растения, растущие в затененных местах - гравилат, чистотел, - имеют температурный порог 45 - 46°С.
4) Растения солнечных и сухих местообитаний - виды гвоздик, коровяк - с температурным максимумом до 48°С.
5) Суккуленты имеют высокий температурный порог - очиток (48,5 - 50°С), у других суккулентных форм температурный максимум может достигать значений 50°С и выше, но не более 54°С.
6) Синезеленые водоросли, в частности водоросль Lyngbia thermalis, живет в горячих источниках при температуре 80 - 83°С.
7) Лишайники (гигрофильные), растущие на увлажненных почвах, достигают температурного порога 70 - 74°С.
8) Лишайники (ксерофитные) имеют температурный максимум 92 - 101°С.
Существует определенная связь между условиями местообитания растений и их жароустойчивостью. Чем суше местообитание, тем выше температурный максимум, то есть тем больше жароустойчивость растений. Из высших растений наиболее жароустойчивыми являются суккуленты. Они характеризуются специфическими анатомо-морфологическими особенностями строения отдельных органов (толстые мясистые листья или стебли, поверхностная корневая система) и определенными физиологическими признаками (низкий уровень метаболических процессов, высокая вязкость цитоплазмы, запасание воды на засушливый период и ее экономное расходование и др.).
Большинство культурных растений относятся к типу мезофитных организмов. Среди мезофитов отдельные виды растений отличаются большей (кукуруза, фасоль, сорго, просо, морковь и др.) или меньшей (горох, томаты, огурцы и др.) жаро- и засухоустойчивостью. К мезофитам относятся растения, обладающие большой пластичностью и приспособляемостью к окружающим условиям, в той или иной мере выносящие кратковременные водные дефициты и перегревы и характеризующиеся сбалансированным водным режимом и мезофитной структурой. У мезофитов жароустойчивость обусловлена более высокой интенсивностью обмена веществ и повышенной вязкостью протоплазмы и концентрацией клеточного сока.
Высокая температура воздуха и резкие ее колебания нередко выступают в качестве факторов, лимитирующих жизнедеятельность отдельных растений в их естественных местообитаниях и распространение растительных видов [12; 64].
Известно, что на четвертой части суши возможен среднегодовой температурный максимум воздуха выше 40°С. Перегрев представляет собой одну из составляющих неблагоприятного действия засухи на растение и нередко наблюдается при достаточной влагообеспеченности на поливных землях в жаркие полуденные часы, что снижает продуктивность орошаемых культур. В условиях действия высоких температур в результате усиленной транспирации температура органов и тканей растений снижается иногда на 10 - 15°С (например, у листьев). Однако во время жары при достаточно высокой влажности воздуха регуляция температуры листьев за счет транспирации ограничена. В таких условиях превышение оптимального температурного уровня приводит к значительным нарушениям протоплазматических структур и внутриклеточных функций. Дж. Левитт, обобщая данные по действию высоких температур на растительные организмы, указывает, что денатурация белков и нарушения структуры мембран являются первичными звеньями повреждения клеток при гипертермии. При чрезвычайно высокой температуре в клетках останавливается движение цитоплазмы, подавляются синтетические (синтез РНК, белка) и усиливаются гидролитические и деструктивные процессы и прекращается рост растений. Жароустойчивость растений во многом определяется продолжительностью действия высоких температур и их абсолютным значением. Кратковременное влияние очень высоких температур (43 - 45°С) может быть таким же губительным, как и продолжительное воздействие более низких, но превышающих их оптимальные значения. Жароустойчивость связана также со стадией развития растений; молодые, активно растущие ткани менее устойчивы, чем старые. В связи с этим наибольший вред высокие температуры наносят растениям на ранних этапах их развития. У пшеницы в фазу кущения в конусе нарастания идет дифференцировка колосков. Высокая температура тормозит этот процесс, что приводит к уменьшению числа цветков в колоске, а также числа колосков в колосе. Для многих растений высокие температуры особенно опасны в период цветения, так как вызывают стерильность цветков и опадание завязей. Органы растений различаются по своей жароустойчивости: более устойчивы побеги и ночки, менее устойчива корневая система. Из тканей наиболее устойчивы камбиальные.
Явления, наблюдающиеся при действии высоких температур на живые клетки, В. Я. Александров классифицирует следующим образом:
1) первичное повреждение нагреванием, проявляющееся изменением структуры клеточных компонентов;
2) вторично развивающееся нарушение клеточных структур и процессов, прямо или косвенно зависящих от первичных повреждений;
3) реактивное повышение стабильности клеточных структур в ответ на нагревание - явление адаптации к высокой температуре;
4) восстановление клетки от тепловых повреждений, называемое репарацией [1; 2].
Клетка является системой, активно поддерживающей свою целостность, поэтому в ответ на действие стрессовых факторов, в частности повышенных температур, в ней реактивно развиваются конструктивные процессы (адаптация, репарация). Состояние клеток через много часов или несколько дней после нагрева будет определяться не только объемом первичного теплового повреждения, но и степенью деструктивного последствия. Очевидно, что чем короче воздействие и чем быстрее фиксируется результат повреждения, тем меньшую роль играют деструктивные последствия. Теплоустойчивость клеток, определимую при условиях, сводящих к минимуму роль деструктивного последействия, называют первичной теплоустойчивостью. Устойчивость, определяемую в длительном эксперименте, где результат зависит от сложного взаимодействия разрушающих и созидающих сил, следует называть общей клеточной теплоустойчивостью.
Для определения первичной теплоустойчивости часто используют регистрацию остановки движения, цитоплазмы, подавление способности к плазмолизу, выход антоцианов из клеток, распределение витальных красителей и другие показатели. Перечисленные реакции очень быстро изменяются под действием высокой температуры. Если производить регистрацию этих показателей после снятия теплового воздействия, то по восстановлению функций можно судить о репарации. Однако растительные клетки могут ликвидировать повреждение, вызванное высокой температурой во время самого прогрева, о чем свидетельствуют многочисленные литературные данные.
Например, при прогреве листьев росянки при 39°С через час наблюдаются патологические изменения структуры митохондрий, но на втором-третьем часу прогрева состояние митохондрий существенно нормализуется. На листьях сахарной свеклы, сахалинской гречихи было показано, что длительное прогревание (90 мин.) приводит к меньшему подавлению фотосинтеза, чем кратковременное (30 мин.).
Таким образом, процессы, развивающиеся в клетке при повреждающем действии нагрева, имеют двойственный характер: одни из них деструктивны (первичные и вторичные повреждения), другие направлены на сохранение целостности клетки (адаптация, репарация).
Для поддержания структуры и функции клетки во время продолжающегося прогрева требуется высокая метаболическая активность. Репарация структуры и функции возможна до тех пор, пока не повреждено энергополноценное дыхание. Во время репарации у теплоустойчивых видов растений обнаруживается значительное накопление органических кислот, антиоксидантов, увеличение синтеза нуклеиновых кислот, белков, углеводов, повышение водоудерживающей способности клеток.
Растительные организмы обладают хорошо выраженной способностью повышать уровень теплоустойчивости в ответ на действие температуры из определенного диапазона, называемого зоной теплового закаливания. Явление повышения теплоустойчивости к супероптимальным нагревам называют закалкой. Величина закалки измеряется разностью температур, характеризующих теплоустойчивость объекта до и после закалки. Она зависит от дозы нагрева. Температура, дающая эффект закалки, должна быть тем выше, чем выше исходная теплоустойчивость объектов. Верхней границей зоны теплового закаливания являются температуры: 38°С - для огурцов; 40°С - для кукурузы и томатов; 43°С - для хлопчатника. Эти температуры индуцируют максимальный закаливающий эффект. Способностью реагировать подъемом первичной теплоустойчивости на супероптимальный нагрев обладают почти все растительные клетки.
Максимальная теплоустойчивость растительных клеток в зоне жарких пустынь проявляется летом после наступления знойных дней. В этих условиях супероптимальные температуры в основном определяют тепловую закалку. Повышение жароустойчивости может происходить не только под действием положительных, но и отрицательных температур (в районах тундры, лесотундры и др.). При действии отрицательных температур на растительные клетки, как и в случае тепловой их закалки, увеличивается обезвоживание протоплазмы и стабильность белковых молекул, что и определяет, по-видимому, повышение устойчивости клеток как к жаре, так и к холоду.
Закаливание - это глубокое воздействие, которое вызывает длительную модификацию, то есть передачу свойств повышенной теплоустойчивости нескольким поколениям.
Жароустойчивость в южных районах означает устойчивость к двум факторам воздействия: к высокой температуре и к прямой солнечной радиации. Повышенная температура наиболее опасна для растений при сильной освещенности. Мезофитные растения борются с перегревом в ясные, солнечные дни с помощью определенного расположения листьев в пространстве и снижения температуры листа за счет транспирации.
Для уменьшения воздействия высокой интенсивности солнечного света растения располагают листья параллельно его лучам, а хлоропласты перемещаются в мезофильных клетках таким образом, чтобы избежать избыточной инсоляции. Кроме транспирации и вертикальной ориентации листьев растения в процессе эволюции выработали и другие приспособления, защищающие их от теплового повреждения: опушение листьев, защищающее от перегрева глубжележащие ткани, большая толщина кутикулярного слоя, малое содержание воды в цитоплазме, высокий уровень водоудерживающей способности протоплазмы и др.
Принято считать, что устойчивость растений к экстремальным температурам тесно связана с особенностями молекулярного состояния воды, с ее структурированностью [16]. Резкое повышение температуры до повреждающих значений приводит к обезвоживанию растений, и тем более сильному, чем значительнее отклонение температурного фактора от оптимального уровня. Например, при действии высокой повреждающей температуры (45°С) содержание воды в семядольных листьях огурца уже через сутки составляло лишь около 50% по сравнению с исходным уровнем. Снижение оводненности тканей растений сопровождается определенными изменениями во фракционном составе воды. Так, при действии высокой закаливающей температуры (40°С) в листьях томата уменьшалось содержание свободной воды, а количество связанной воды увеличивалось. Следовательно, в процессе действия закаливающих температур снижение подвижности воды способствует повышению устойчивости. Однако в условиях высокой температуры растение должно сохранить определенное количество подвижных форм воды, чтобы обеспечить процессы транспирации и протекания метаболических реакций.
Температура существенно влияет на скорость водного обмена у растений. Но мнению одних авторов, это связано с изменением скорости активного поглощения солей и как следствие - с изменением величины движущей силы транспорта воды. Другие полагают, что под действием температуры прежде всего изменяется проницаемость мембран. Кроме того, температура оказывает влияние на скорость биохимических реакций, что также отражается на водообмене.
П. А. Генкелем и Н. А. Шеламовой было введено понятие гомеостатической воды, которая поддерживает гомеостаз растения. Ниже уровня содержания гомеостатической воды они не могут осуществлять свою жизнедеятельность и погибают. Различные экологические группы растений содержат различное количество гомеостатической воды: покрытосеменные пойкилоксерофиты - от 18 до 20%, мезофиты - от 35 до 50% [12; 14].
Высокая температура оказывает влияние на содержание и состав липидов, определяющих структуру и функцию мембран. Данные В. Ф. Альтергота показывают, что под влиянием температуры в 37°С у двухдневных проростков пшеницы увеличивалось почти на 50% содержание фосфолипидов (контроль 24°С). Прогрев при 50°С закаленных проростков пшеницы приводил к увеличению содержание фосфолипидов в мембранах. Обнаружено также увеличение содержания нейтральных липидов как в хлоропластах, так и в митохондриях у пшениц при действии высоких температур (45°С, 3 часов) [5; 6].
Характер изменений содержания липидов мембран под действием экстремальных температур зависит от устойчивости растений. Теплоустойчивые сорта люцерны содержали большие количества сульфо- и фосфолипидов при 30°С, чем при 15°С. У холодоустойчивых сортов люцерны уровень липидов при нагревании не изменялся.
Следовательно, у устойчивых растений при действии высоких температур происходят существенные изменения в липидном составе мембран: накапливаются нейтральные липиды, богатые энергией вещества, и фосфолипиды, количественные соотношения которых определяют мембранную проницаемость.
Повышение температуры оказывает влияние и на жирнокислотный состав липидов. У ряда растений в режиме более высоких температур (43/30°С) увеличивается содержание насыщенных жирных кислот по сравнению с более низкими температурами (23/18°С).
Текучесть или вязкость мембран имеет решающее значение для целого ряда их функций, в том числе ферментативного катализа, транспорта воды и ионов, рецепции экзогенных сигналов и др. Для того чтобы при повышении температуры мембрана у устойчивых растений могла нормально функционировать в ее липидном составе, должны преобладать насыщенные жирные кислоты. Чем меньше число двойных связей, тем выше может быть температура, при которой сохраняется необходимая вязкость мембран. Стратегия адаптации у жароустойчивых растений определяется главным образом путем изменения либо числа двойных связей в молекулах жирных кислот, либо типов жирных кислот, включающихся в молекулы фосфолипидов во время их синтеза. Функционирование отдельных ферментов, связанных с мембраной, часто зависит от локальной вязкости их микроокружения. При действии повышенной температуры, близкой к оптимальному значению для растительных организмов, высокая текучесть мембран способствует большей каталитической эффективности мембранно-связанных ферментов. В этих условиях ферментный белок является подвижным и имеет возможность изменять свою конформацию или перемещаться в пределах мембраны во время катализа. В слишком жидком липидном бислое (при субмаксимальной температуре) липидная фаза менее упорядоченна, что может приводить к дезинтеграции структуры мембран и как следствие - к инактивации мембранно-связанных ферментов. Установлено, что текучесть мембранных липидов определяет активность связанных с мембранами ферментов. Так, Na+, K+-зависимая АТФаза с уменьшением текучести (ненасыщенности) липидов теряет активность, а Ca2+-зависимая - увеличивает. В то же время Mg2+-зависимая АТФаза почти не чувствительна к изменению вязкости мембранных липидов.
Белки, являясь важными ингредиентами клетки, участвуют практически во всех проявлениях ее жизнедеятельности [1; 2; 55; 64]. Температура оказывает существенное влияние на структуру белковых молекул, на скорость процессов биосинтеза белка, спектр синтезируемых белков, свойства белков, имеющихся в клетке.
Репарация теплового повреждения возможна без синтеза белка в случае слабого повреждения клетки, при этом происходит ренатурация денатурированных белков. Принцип полустабильности (семистабильности или семилабильности) положен в основу конформационной гипотезы. В. Я. Александров рассматривает терморезистентность как производное устойчивости наиболее важных компонентов клетки: в первую очередь белков, а также нуклеиновых кислот и липидов [1; 2]. Уровень стабильности данных макромолекул является температурозависимым. Повышение температуры за пределы оптимума приводит к уменьшению их жесткости и к значительной лабилизации. Чем сильнее отклонение значения температуры от оптимального уровня, тем большие изменения обнаруживаются в структуре макромолекул (больший разрыв слабых связей и значительная лабилизация белковых молекул). Адаптация белковых молекул (как и других макромолекул) к повышенной температуре осуществляется за счет увеличения их жесткости. По- видимому, в природе в процессе эволюции поддерживается корреляция между уровнем конформационной гибкости белковых молекул и усредненной температурой среды. Гибкость и подвижность конформации белковой молекулы обеспечивает способность ее адекватно отвечать на присутствие в среде тех или иных факторов, т. е. быть регулируемой этими факторами. Уровень конформационной гибкости макромолекул должен быть согласован с температурным режимом работы белка в клетке. Для объяснения повышения теплоустойчивости клеток в ответ на тепловой шок Левитт предлагает гипотезу, основанную на том, что при повышении температуры вплоть до 60 - 70°С прочность гидрофобных взаимодействий возрастает, а водородные связи ослабляются. При быстром тепловом супероптимальном воздействии происходит частичное развертывание белковых молекул, после чего следует их свертывание в форму, обогащенную гидрофобными связями и тем самым более термостабильную. При долговременных адаптациях к высокой температуре синтезируются термостабильные белки, у которых также за счет уменьшения водородных связей увеличено число гидрофобных взаимодействий, что делает макромолекулу белка более термоустойчивой [64].
Таким образом, живые организмы способны регулировать уровень жесткости своих макромолекул, создавая в протоплазме условия, влияющие на прочность слабых связей (pH, ионный состав, содержание воды и др.). Тем самым снижается или увеличивается конфирмационная подвижность макромолекул, что позволяет изменять биохимическую активность. Ферментный белок, изменяя свою конформацию под влиянием температуры, меняет средство к субстрату или кофактору, что безусловно сказывается на его активности.
Следовательно, при быстрой адаптации растительных организмов к высокой температуре (в случае секундных закалок) прирост теплоустойчивости, но всей видимости, обеспечивается за счет конфирмационных изменений макромолекул клеток, и в первую очередь белков. Для приспособления растений к более высоким температурам необходимо повышение структурной стабильности белка. Оно может быть связано с изменением числа (или тина) слабых взаимодействий. При высоких температурах в белках происходит перестройка системы водородных связей и гидрофобных взаимодействий между боковыми группами. Белок принимает одну из энергетически выгодных конформаций.
Известно, что некоторые термофилы (микроорганизмы) содержат белки, устойчивые к высоким температурам. Эти белки-ферменты слабо денатурируют при нагревании. Если такие организмы поместить в условия низких температур, то они погибают. Полагают, что молекулы белков у данных организмов содержат большее число взаимодействующих между собой аминокислотных остатков [22]. В результате даже при очень высокой температуре сохраняется достаточное для поддержания активной конформации число связей. При понижении температуры в белке появляется избыток водородных и других слабых связей, что может изменить конформацию ферментов и привести к инактивации белка. Термофильные бактерии способны значительно увеличивать скорость синтеза белков-ферментов при высокой температуре. Бактерии, обитающие в горячих источниках (+70, +80°С), отличаются значительной скоростью репарации - вместо инактивированных белков в структурные элементы клетки встраиваются вновь синтезируемые молекулы.
Возможно, что высшие наземные растения, живущие в условиях жаркого климата, приспосабливаются к высоким температурам аналогичным способом, т. е. способны к синтезу термостойких белков - белков теплового шока (БТШ). Об этом свидетельствуют многочисленные данные. Так, К. Мотесом было показано, что листья махорки под влиянием высокой температуры (+50°С) преждевременно стареют и желтеют. В то же время в обработанных кинетином (1 - 2 мг/л) листьях даже при действии повреждающей температуры (+52°С) усиливался синтез белка и не наблюдалось процессов старения клеток [68].
В настоящее время известно, что в ответ на температурный стресс синтезируются новые мРНК и соответствующие им "шоковые" белки, тогда как синтез мРНК и белков, характерных для обычного состояния организма, резко тормозится или полностью прекращается [60]. Перестройка белкового аппарата клеток при действии высоких температур связана с модификацией матричной активности хроматина, переводом в нетранслируемое состояние "дошоковых" мРНК и с изменением свойств самих рибосом. Данные электронно-микроскопического анализа свидетельствует о том, что при закаливающих температурах в клетках растений увеличивается поверхность эндоплазматического ретикулума и наблюдается изменение компартментации рибосом- в гиалоплазме уменьшается число свободных моно- и полисом и увеличивается доля рибосом, связанных с мембранами ЭПР. Учитывая то, что на свободных и мембранно-связанных рибосомах синтезируются разные классы белков, можно полагать, что при термическом закаливании происходит новобразование белков с новыми свойствами.
В растительных клетках при действии теплового стресса обнаружено образование специфических БТШ и прекращение (или сильное замедление) синтеза всех остальных белков. В частности, прогревание проростков сои в течение 1 - 4 часов при температуре 40°С приводит к образованию новых белков (по включению 3H-лейцина). Синтез белков, характерных для условий с нормальной температурой (28 - 30°С), снижается. Образование БТШ отмечается в первый час действия теплового шока и продолжается в течение последующих 4 - 6 часов. При перенесении проростков сои в "номальные" температурные условия (28 - 30°С) после теплового шока (40°С 4 часа) наблюдается прогрессивное уменьшение образования БТШ и восстановление синтеза обычных белков [9; 28; 60; 61].
Белки теплового шока синтезируются у растений, относящихся к разным семействам. При действии высоких температур (40°С и выше) проростки сои, гороха, подсолнечника, пшеницы, ячменя, овса, риса, кукурузы, проса способны к синтезу БТШ. Например, прогревание проростков кукурузы в течение 1 часа от 27°С до 44°С приводило к незначительным изменениям в синтезе общих белков. При более высокой температуре (выше 44°С) образование их резко снижалось, а при 50°С составляло 15% от контрольного уровня (27°С). В то же время тепловой шок индуцировал образование новых полипептидов (по включению 14С-лейцина) с молекулярными массами 108, 89, 84, 76, 73 и 18 кД. Причем, включение метки в полипептиды, образование которых индуцируется гипертермией, обнаруживается уже при 35°С. Интенсивность этого процесса возрастает с повышением температуры до 44°С. При возвращении проростков кукурузы после шока в нормальные температурные условия прекращается синтез БТШ и восстанавливается синтез белков, характерных для оптимальных температур. Сходные результаты получены и для других растительных объектов.
Длительность температурного воздействия влияет как на количество, так и на качество белков теплового шока. В колеоптилях и корнях 3-дневных проростков злаков (пшеницы, ячменя, овса) в ответ на тепловой шок (40°С) синтезируются 12 типов БТШ, относящихся к трем группам: с высокой (103 - 70 кД), средней (62 - 32 кД) и низкой (17 - 16 кД) молекулярной массой. Существенно, что синтез БТШ, образующихся в начальный период действия температуры, тормозился через 6 часов и одновременно с этим появлялись 12 новых (поздних) БТШ с высокой (99 - 83 кД), средней (69 - 35 кД) и низкой (15 - 14 кД) молекулярной массой. Через 12 часов действия повышенной температуры синтезируются БТШ с молекулярной массой 89, 45 и 38 кД.
Таким образом, синтез БТШ в ответ на тепловой шок представляет многофазную реакцию: образуются как ранние белковые молекулы (синтез в начальный период действия шоковой температуры), так и поздние формы белков с иной молекулярной массой.
Полагают, что одна из основных функций БТШ - структурная. Белки теплового шока, являясь составной частью структуры отдельных фрагментов ядерного и цитоплазматического скелета, стабилизируют состояние содержимого клетки в стрессовых условиях.
Можно полагать, что термоустойчивость клеток возрастает в связи с синтезом БТШ. Так, помещение проростков сои в условиях экстремальной температуры (52°С 1 часа) сразу же после оптимальной (25°С) приводит к 100%-ной их гибели. Однако, если проростки сои выдерживать вначале 1 час при температуре 34°С (после 25°С), а затем поместить их в стрессовые условия (52°С), то наблюдается значительно меньший процент гибели растений (50%). Выживаемость клеток была почти полная, когда температуру поднимали постепенно, со скоростью 3°/ч. до высоких значений (52°С). В двух последних случаях в проростках сои отмечается синтез белков теплового шока.
Следовательно, БТШ могут предохранять растительные клетки от гибели при действии повреждающих температур. По-видимому, у высших и низших организмов реакция синтеза стрессовых (шоковых) белков на сигналы внешней среды относится к универсальным механизмам быстрой адаптации. В процессе эволюции сохраняется способность организмов к синтезу БТШ, что может указывать на большую значимость этой реакции клеток на стресс.
Таким образом, при действии температуры в высокой дозе очень быстро изменяется экспрессия генов. Существует несколько уровней ответа генома клетки на стресс. Клетка воспринимает температурный фактор с помощью рецепторов. Результатом такого взаимодействия является образование сигналов, которые передаются в ядро, взаимодействуют с соответствующими генами и изменяют экспрессию последних. Затем включается вся цепочка внутриклеточных процессов по схеме: ДНК→шоковые мРНК→белки теплового шока→длительные адаптивные реакции→термоустойчивость.
Первичной мишенью действия температур, по-видимому, могут быть протеинкиназы, которые после активации факторами внешней среды фосфорилируют цитоплазматические белки. Такие активированные (фосфорилированные) цитоплазматические белки, как показано на изолированных ядрах дрозофиллы, участвуют в активации генов БТШ. Высокая скорость синтеза специфических белков при тепловом шоке определяется тем, что при стрессе активация хроматина не связана с существенными изменениями структуры нуклеосом. Отсутствие гистонов в промоторной гиперчувствительной зоне хроматина, видимо, обеспечивает быструю реакцию генома на температурный стресс. Считают также, что стрессовые мРНК имеют повышенное сродство к рибосомам.
Синтез БТШ рассматривается как пример быстрой адаптации растительных клеток к действию высоких температур.
При значительной дозе гипертермии, при которой БТШ не могут уже обеспечивать сохранность внутриклеточных структур, происходит разрушение жизненно важных биополимеров.
Основной причиной повреждения и гибели растений под действием высокой температуры является распад белка с образованием аммиака, вызывающим отравление растительных клеток и их гибель. По мнению Н. С. Петинова и Ю. Г. Молотковского, обезвреживание аммиака происходит в процессе дыхания. Под влиянием высоких температур у жароустойчивых растений снижается дыхательный коэффициент и накапливаются органические кислоты, образующие с аммиаком аммонийные соли. Кроме того, происходит образование амидов и аминокислот при аминировании кетокислот [38]. Наконец, аммиак может связываться с аланином, синтез которого резко возрастает при +30÷+40°С ("аланиновый эффект").
Для репарации структуры протоплазмы в неблагоприятных температурных условиях необходим постоянный приток энергии. Поэтому устойчивость растений к действию экстремальной температуры во многом определяется состоянием основных энергодающих процессов - фотосинтеза и дыхания.
Зависимость нетто-газообмена от температуры определяется разностью между скоростями фотосинтетического усвоения CO2 и процессов дыхания при той же температуре. Следует отметить, что фотосинтез более чувствителен к температуре, чем дыхание (рис. 9).
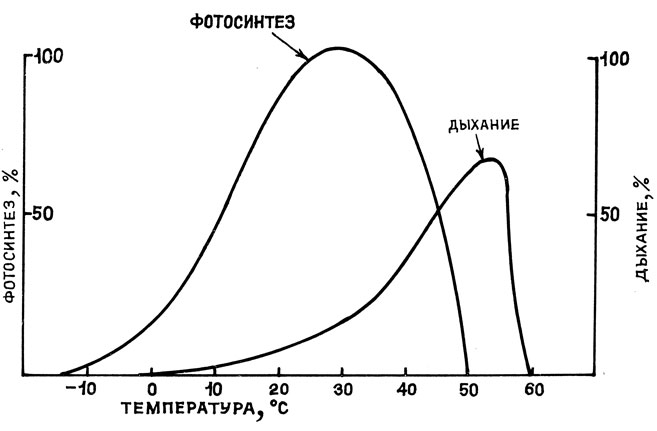
Рис. 9. Схема зависимости интенсивности фотосинтеза и дыхания от температуры (по В. Лархеру, 1978)
В пределах оптимальной области температур нетто-фотосинтез достигает более 90% своей максимальной величины. У C3-растений температурный оптимум фотосинтеза зависит не столько от вида, сколько от теплового режима в естественном местообитании в период активной жизнедеятельности. Растения, которые произрастают в тени и меньше нагреваются, имеют оптимум между 10 и 20°С. Это относится как к раноцветущим, так и высокогорным формам, растущим в такое время года или в таких местах, для которых характерны низкие средние температуры воздуха. Травянистые растения солнечных мест обитания имеют наивысшую продуктивность фотосинтеза между 20 и 30°С (табл. 5). От температуры зависит прежде всего скорость темновых реакций (Q10 от 2 до 4). Световые реакции меньше зависят от температуры и для них соответствующие значения Q10 составляют 1.0 - 1.2.
![Таблица 5. Температурные условия нетто-фотосинтеза растений различных экологических групп [32]](pic/000024.jpg)
Таблица 5. Температурные условия нетто-фотосинтеза растений различных экологических групп [32]
У C4-растений оптимум продуктивности лежит при температуре выше 35°С, в отдельных случаях даже при 50°С. C4-путь ассимиляции углерода служит генотипической предпосылкой для заселения крайне жарких местообитаний.
У CAM-растений, прежде всего у кактусов, температура во время светового периода не имеет, по-видимому, особого значения для интенсивности фотосинтеза. Темновая же фиксация CO2 обнаруживает температурную зависимость. Ее оптимум соответствует более низким ночным температурам и лежит между 5°С и 15°С, а выше 25°С этот процесс резко замедляется.
При перегреве продуктивность фотосинтеза резко падает, а дыхание продолжает расти. В результате высвобождаются большие количества CO2. У верхней границы нетто-фотосинтеза образующаяся при дыхании углекислота еще реассимилируется, однако при дальнейшем повышении температуры происходит уже выделение CO2. Поэтому температурный максимум (верхний предел) нетто-фотосинтеза можно рассматривать как тепловую точку компенсации, которая достигается тем раньше, чем чувствительнее к теплу фотосинтез и чем круче подъем дыхания. Область верхнего предела нетто-фотосинтеза подвержена сезонным и адаптивным изменениям [23].
Показано, что наиболее чувствительным звеном к действию повышенной температуры в аппарате фотосинтеза является фотосистема II. Повреждение фотосинтетических мембран высокой температурой выражается в диссоциации светопоглощающего комплекса и реакционного Центра фотосистемы II, что приводит к изменению эффективности светопоглощения [36].
Важнейшими структурами, определяющими температурный оптимум и температурные пределы реакций фотосинтеза у растений, являются тилакоидные мембраны хлоропластов - структуры, где происходят многие процессы переноса энергии, связанные с поглощением света и фиксацией CO2. Потенциальная эффективность и температурные пределы процессов фотосинтеза у зеленых растений меняются при изменении температуры среды. Полагают, что непосредственной реакцией на температурное воздействие является изменение текучести липидов, входящих в состав мембран. Сохранение эффективности фотосинтеза у растений, адаптированных к теплу, обусловлено изменениями вязкости липидов. У видов или популяций растений, переносящих более высокую температуру, повышенное содержание насыщенных жирных кислот в фосфолипидах и галактолипидах обеспечивает термостабильность тилакоидных мембран. Кроме того, повышение стабильности хлорофилл- белково-липидного комплекса у термотолерантных форм обусловлено высокой устойчивостью мембранных белковых компонентов к действию супероптимальных температур.
От состояния липидной структуры тилакоидов в значительной степени зависит активность фотохимических реакций и фотофосфорилирования. Прогрев хлоропластов шпината при температуре около 30°С приводит к снижению интенсивности фотофосфорилирования. Более высокая температура (около 40°С) стимулирует фотохимическую активность хлоропластов и подавляет циклическое фотофосфорилирование.
Реакция растений на высокую температуру зависит от их адаптивных возможностей. В частности, прогрев (45°С) стимулирует или практически не изменяет фотохимическую активность хлоропластов пшеницы устойчивых сортов. У неустойчивых сортов пшеницы фотохимическая активность значительно ингибируется при действии температуры 45°С. Если фотохимическая активность ингибируется на 2/3 от исходного уровня, то фотосинтез подавляется почти полностью. Причем фотохимическая активность восстанавливается после теплового стресса у закаленных растений значительно быстрее, чем у незакаленных.
Основной фермент темповых реакций фотосинтеза - рибулезо-1,5-бисфосфаткарбоксилаза - крайне жароустойчивый белок, который инактивируется только при температурах, вызывающих некроз на листьях.
Дыхание более устойчиво к повышенным температурам и подавляется при значительно большей дозе воздействия нагрева, чем фотосинтез (рис. 9). Следовательно, дыхание может быть поставщиком энергии для репарации более чувствительных к температуре внутриклеточных структур и функций. Результаты исследования энергетики дыхания растений при повышенной температуре противоречивы [43; 44; 6; 10; 19; 20]. Согласно одним данным, нарушение энергетики опережает необратимое нарушение дыхательного газообмена, по другим - энергетика не претерпевает сколько-нибудь существенных изменений даже при значительном ингибировании дыхания.
По данным О. А. Семихатовой с сотрудниками, реакция газообмена у листьев разных растений на воздействие высоких температур неоднозначна.
У гороха, примулы, яснотки белой, астрагала, лисохвоста при повышении температуры наблюдается двухфазное изменение интенсивности дыхания: некоторое возрастание вначале сменяется в дальнейшем спадом. Величина дыхательного коэффициента (ДК=CO2/O2) остается близкой к единице при коротких экспозициях (15 - 60 мин.) почти до критических температур (43° и 45°С). Это свидетельствует об отсутствии у растений кардинальных изменений химизма дыхания в данном диапазоне температур. При более высокой температуре (49°С) интенсивность поглощения кислорода снижается в большей степени, чем выделение CO2, и дыхательный коэффициент при этом значительно повышается (ДК=1,5-1,7). Увеличение ДК связано с необратимым повреждением клеток.
Имеется группа растений (незабудка, купальница, пижма обыкновенная, лютик едкий и др.), у которых повышение температуры до 40°С приводит к вспышке интенсивности газообмена. Причем, уровень поглощения кислорода тем выше, чем больше температура. При этом ДК падает до значений 0,4 - 0,5. Значительное снижение величины дыхательного коэффициента свидетельствует о нарушениях в дыхательном метаболизме в листьях данных растений при субкритических температурах. У некоторых растений (лютик золотистый, остролодочник, солянка восточная) при действии высоких температур обнаруживается незначительное снижение интенсивности дыхания.
Степень сопряжения окисления и фосфорилирования зависит от структуры мембран митохондрий: их целостности и особенностей их белкового и липидного компонентов. Как свидетельствуют данные О. А. Семихатовой, повышенная температура не оказывает влияния на энергетическую эффективность дыхания у гороха. Даже при критической для дыхания температуре (47°С) относительная величина скорости генерации макроэргических фосфорных соединений возрастала примерно в той же мере, что и интенсивность поглощения кислорода. Увеличение длительности воздействия повышенной температуры не вызывало изменений в энергетической эффективности дыхания до тех пор, пока у растений не появлялись видимые признаки повреждения [43; 44].
Однако В. Н. Жолкевич и З. С. Сагатов приходят к выводу, что продолжительное действие субкритических температур (5 часов при 43°С и 4 часа при 45°С) вызывает у гороха снижение степени сопряжения окисления и фосфорилирования. При этом наблюдалось уменьшение содержания АТФ, особенно значительно при 45°С. Для нарушения окислительного фосфорилирования при критической температуре (47°С для растений гороха) оказалось достаточно одночасовой экспозиции [20].
Ответная реакция энергетического обмена при дыхании на перегрев зависит от дозы температурного фактора. 3. С. Сагатов на ассимилирующих побегах саксаула показал, что при действии оптимальных (30°С, 6 часов) и субкритических (48°С, 1 час) температур показатели, характеризующие уровень энергообмена, не изменяются. Увеличение дозы нагрева (48°С, 6 часов и 52°С, 1 час) приводит к нарушениям в энергообмене. Разобщение окисления и фосфорилирования происходит при длительном (3 часа) воздействии критических температур (52°С). В. Ф. Альтергот также приходит к выводу, что перегрев приводит к разобщению окисления и фосфорилирования [6].
При неблагоприятных условиях, в частности при повышенных температурах, появляется особая "статья" расхода энергии, связанная с энергетическими затратами на процессы репарации повреждений. Это в основном синтез веществ, которые разрушаются или расходуются в неблагоприятных условиях. Энергия необходима для поддержания структурных элементов протоплазмы клеток в условиях стресса. Насколько успешно идут репарации в клетках, по-видимому, могут указывать изменения такого интегрального показателя энергетического обмена, как Q/E·100, где E выражает то максимальное количество энергии, которым располагает живая система для выполнения энергозависимых функций, Q суммирует энергопотери, обусловленные всем ходом жизнедеятельности, в том числе и энергию, освободившуюся при окислении, но не аккумулированную в АТФ. Как считает В. Н. Жолкевич, величина Q/E·100 свидетельствует о том, насколько процесс дыхания может компенсировать всю сумму энергетических потерь в условиях стресса. Если Q/E·100 меньше 100%, значит идет запасание энергии (активные синтезы, рост), если превышает 100% - энергетический баланс, клеток отрицателен (преобладают деструктивные процессы). Показано, что при действии субкритических температур (43°С, 45°С), как при коротких (1 час), так и при длительных (4 и 5 часов) экспозициях, увеличивается количество суммарных энергопотерь, величина Q значительно превосходит величину E, вследствие чего энергетический баланс становится отрицательным. Следует отметить, что это происходит уже на начальном этапе действия субкритической температуры, несмотря на сохранение определенного уровня сопряженности окисления с фосфорилированием и достаточно высокое содержание АТФ. Значительное увеличение суммарных энергопотерь в клетке в данных температурных условиях, видимо, вызвано интенсификацией гидролитических и деструктивных процессов и ослаблением сопряжения окисления и фосфорилирования.
Следовательно, в процессе эволюции формировались, апробировались и закреплялись различные адаптивные механизмы, делающие растения менее уязвимыми по отношению к высокотемпературному стрессу. Выработка таких механизмов шла в нескольких направлениях: уменьшение перегрева за счет транспирации; защита от тепловых повреждений (опушение листьев, большая толщина кутикулярного слоя); стабилизация метаболических процессов (более жесткая структура мембран, высокая вязкость протоплазмы, низкое содержание воды в клетке и др.), высокий уровень фотосинтеза и энергополноценного дыхания. Целый арсенал защитно-приспособительных реакций обеспечивает существование вида, однако не всегда гарантирует выживание отдельных особей.
Адаптивные возможности организмов определяют скорость метаболических процессов в репарационный период. У устойчивых видов действие повышенных температур вызывает обратимые структурно-функциональные изменения. Репарационные процессы у них происходят как при снятии стрессового фактора, так и в процессе действия высоких температур. У неустойчивых видов тепловой шок приводит часто к необратимым структурно-функциональным изменениям. Репаративные возможности у данных организмов ограничены.
Повышение жароустойчивости. Для повышения термостойкости растений широко используются различные методы закаливания. В результате температурных обработок у растений повышается вязкость цитоплазмы, изменяется метаболизм в направлении, обеспечивающем процессы репарации и адаптации.
Растительные организмы обладают хорошо выраженной способностью повышать уровень теплоустойчивости в ответ на действие температуры из определенного диапазона, называемого зоной теплового закаливания. Более низкие температуры, относящиеся к фоновой зоне и являющиеся оптимальными для роста и развития растений, не сказываются на теплоустойчивости, а более высокие температуры вызывают ее снижение и при длительной экспозиции могут значительно повреждать растительные организмы [19; 20].
В. Ф. Альтергот предложил метод ступенчатого закаливания проростков пшеницы и гороха для повышения их жаро- и засухоустойчивости. Он состоит из следующих этапов: 1) воздействие на растение определенной мерой неблагоприятного фактора, вызывающей обратимые повреждения (фаза стресса); 2) создание оптимальных условий освещения, питания, температуры в течение продолжительного времени для репарации повреждений (фаза восстановления функций); 3) чередование состояния стресса и репарации; 4) усиление состояния стресса на каждой последующей ступени путем увеличения меры воздействия (фактор×времи), но в пределах обратимости повреждений. Если чередовать действие повышенных температур с нормальным режимом (40°С×12 часов; +22°×16 часов; +48°×2 часов, +22°×48 часов; +50°×2 часов; +22°×48 часов; +52°;×2 часов), то происходит восстановление повреждений у проростков пшеницы при тепловом стрессе (52°С). При этом получаются жаростойкие растения с большой регенерационной способностью. Обязательным требованием при разработке режимов закаливания является полная репарация повреждений в фазе восстановления функции.
В. Ф. Альтерготом был разработан метод тепловой обработки семян пшеницы (постепенный подъем температуры от 20°С до 50°С в течение 8 часов). В результате такой обработки снижается влажность зерна с 38 - 40% до 15 - 16%, повышается всхожесть семян на 11 - 13% и увеличивается энергия их прорастания на 17 - 20%.
Повышение жароустойчивости и урожая у картофеля и сахарной свеклы может быть достигнуто за счет теплового закаливания соответственно клубней и обогрева семян при температуре 22 - 30°С.
Адаптированные к высокой температуре растения отличаются от незакаленных высоким уровнем синтеза ВИК и белка, его гетерогенностью и качественными изменениями в спектре, увеличенным содержанием водорастворимых и щелочерастворимых белков. У закаленных растений повышается содержание хлорофилла, увеличивается интенсивность фотосинтеза и уровень АТФ.
Жароустойчивость растений может быть также повышена при обработке семян 2,4 Д (0,001 - 0,005%), суперфосфатом (5,0%), хлористым калием (0,2%), хлористым кальцием (0,2%), азотно-кислым аммонием (0,25%), борной кислотой (0,05%), а также при увеличении в составе удобрений фосфора, калия, кальция, меди, марганца, молибдена и цинка. Кроме того, применение мелкодисперсного и импульсного дождевания приводит к устранению водного дефицита. Регулирование форм, соотношений, доз и сроков внесения макро- и микроудобрений способствует усилению деятельности корневой системы растений, улучшению общей оводненности листьев, повышению степени гидратации хлоропластов, продуктивности транспирации, усилению фотосинтеза и активности ферментных систем, главным образом ферментов первичной ассимиляции азота (нитратредуктаза, глутаминсинтетаза, глутаматдегидрогеназа).
Для повышения жароустойчивости было использовано свойство солей кальция повышать вязкость протоплазмы. Обработка семян проса и пшеницы CaCl2 (0,1%) оказывает благоприятное влияние на рост и развитие растений, а также и на повышение урожая. Многочисленные опыты показали, что предпосевная обработка семян мягких и твердых пшениц, проса, подсолнечника раствором хлористого кальция значительно повышала жароустойчивость растений и, как следствие, урожай зерна в засушливых условиях [6].
Большую роль в повышении устойчивости растений к воздействию температур играют микроэлементы. М. Я. Школьник предложил замачивать семена или зерновки не в воде, а в растворе, содержащем бор и другие микроэлементы. Под влиянием бора значительно возрастает вязкость цитоплазмы, что способствует повышению жароустойчивости. Сернокислый цинк (0,05%) сдвигает дыхание в сторону образования органических кислот, что способствует обезвреживанию клеток от аммиака. Кроме того, цинк увеличивает гидрофильную вязкость протоплазмы, что также повышает жароустойчивость [56].
Как считает А. М. Алексеев, действие микроэлементов обусловлено влиянием положительно гидратирующих ионов и накоплением сахаров и низкомолекулярных неэлектролитов, стабилизирующих структуру воды. Бор, марганец, цинк, медь, молибден повышают интенсивность фотосинтеза, особенно при недостаточном водоснабжении. Медь, бор и особенно кобальт существенно повышают содержание адонозинтрифосфата. Полагают, что одним из проявлений положительного влияния бора и цинка на жаро- и засухоустойчивость растений является их способность снижать активность рибонуклеазы, распад полисом и деградацию рибосом [3].
Таким образом, сбалансирование питания растений микро- и макроэлементами повышает их устойчивость к засухе, пониженным и высоким температурам и к различным заболеваниям.
Важным и перспективным способом регулирования устойчивости растений к различным факторам среды является использование наряду с удобрениями и орошением физиологически активных веществ. В частности, при действии высоких температур проявляется защитное действие цитокининов. В работах К. Мотеса было установлено, что обработка листьев табака кинетином способствует увеличению их устойчивости к действию высоких температур, а также активации восстановления структурных компонентов клетки после засухи. Полагают, что защитное действие кинетина связано с увеличением способности клеток растений удерживать подвижные метаболиты и усиливать приток питательных веществ. Кроме того, показано, что обработка листьев кинетином стабилизирует мембраны и тем самым ослабляет действие высоких температур. О. Н. Кулаева считает, что защитное действие цитокининов проявляется через воздействие их па структурное и функциональное состояние различных компонентов клетки и, в частности, на состояние их мембранного аппарата [29].
В настоящее время известно, что брассиностероиды, способствуя поддержанию синтеза белка в условиях гипертермии, повышают устойчивость растений к высоким температурам. В 1989 г. О. Н. Кулаевой с сотрудниками было установлено, что гомобрассинолид в концентрации 10-8M, 10-6M при оптимальной температуре (23°С) значительно активирует синтез белка в листьях пшеницы сорта "Саратовская-29". При тепловом шоке (40°С) происходит синтез основных БТШ (100, 98, 90, 77, 20 кД) и некоторых минорных полипептидов. Гомобрассилолид (10-6M) на фоне высокой температуры (40°С) не препятствует синтезу БТШ и обеспечивает усиление синтеза тех полипептидов, которые обнаруживаются в условиях оптимальной температуры [30]. Следует отметить, что в присутствии данного фитогормона синтез белка в листьях при 43°С сохранялся на том же уровне, что и при 23°С. Кроме того, гомобрассинолид (10-6M) препятствовал резкому падению синтеза белка при экстремальной температуре (45°С).
Следовательно, в условиях гипертермии гомобрассинолид, увеличивая общий синтез белка, сдвигает в область более высоких температур пороговую температуру, при которой начинается резкое торможение этого процесса. Повышение устойчивости белкового синтеза под действием гомобрассинолида в значительной степени определяется спецификой и напряженностью метаболических процессов.
При гипо- и гипертермии в растительном организме возникает сложный комплекс функциональных изменений в интенсивности фотосинтеза, дыхания, наблюдаются отклонения в углеводном, азотном, нуклеиновом обменах, в активности и направленности действия ферментов. Кроме того, изменяется структура органоидов клетки, в частности митохондрий и хлоропластов, выполняющих энергетическую функцию. Направленность и глубина этих сдвигов зависит от степени терморезистентности растений.
Устойчивость растений к низким отрицательным температурам обеспечивается в основном действием протекторных соединений, которые синтезируются в процессе закаливания. Процессы закаливания к холоду и теплу характеризуются наличием определенного лаг периода, который обусловлен временем, необходимым для накопления тех или иных изменений. Этот временной параметр требуется растениям для запуска и развертывания адаптивной генетической программы, продуктами которой являются стрессовые мРНК, стрессовые белки и другие низкомолекулярные протекторные соединения. Адаптация растений к низким и высоким температурам в значительной степени связана с изменениями физико-химического состояния мембран, их функциональной активности, в частности барьерных свойств плазмалеммы. Эти температурозависимые изменения обеспечивают возрастание резистентности клеток и растения в целом к гипо- и гипертермии.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'