2.3.1. Влияние низких отрицательных температур (морозоустойчивость растений)
Морозоустойчивость - способность клеток, тканей и целых растений благодаря ряду физиолого-биохимических механизмов, которые выработались у них в процессе эволюции под влиянием условий существования, переносить охлаждение ниже 0°С.
В ряде монографий и обзоров широко представлено состояние проблемы морозоустойчивости озимых культур с физиолого-биохимических позиций [35; 40; 42; 50; 49; 18; 26; 33].
Значительный вклад в проблему морозустойчивости внес Н. А. Максимов. Он развивал представления об обезвоживании клеток под действием мороза и показал, что истинной причиной повреждения и гибели растений при отрицательных температурах является давление кристаллов льда на обезвоженные клетки. Чем больше образуется льда в межклетниках, тем больше гибнет клеток. Кроме того, Н. А. Максимов наметил пути сохранения растений от губительного действия льда. Важную роль он придавал химической защите клеток (растворам сахаров и солей, которые имеют более низкую температуру замерзания по сравнению с водой). Максимов выдвинул положение о том, что образующийся в клетках лед оказывает не только водоотнимающее действие, то есть обезвоживает протоплазму, но и вызывает коагуляцию биоколлоидов протоплазмы.
Положительное влияние сахаров на морозостойкость растений неоднократно отмечал Дж. Левитт. Он показал, что листья капусты, обработанные глицерином или раствором сахаров при температуре - 30°С, повреждались льдом меньше но сравнению с листьями, не обработанными осмотиками [62].
Для сохранения целостности мембран растительных клеток при температурах замерзания большое значение имеют защитные вещества - протекторы. Они предотвращают изменения в биомембранах, взаимодействуя неспецифически с образуемыми токсическими соединениями и специфически - с самими мембранами. Эффективными протекторами мембран хлоропластов при их промораживании в течение 3 - 4 часов при -25°С являются раффиноза (0,02 - 0,06 M) и сахароза (0,04 - 0,06 M) в присутствии 0,1 M NaCl, а также янтарная, лимонная и уксусная кислоты в определенных концентрациях в присутствии NaCl.
В настоящее время показано, что сахара обладают хорошо выраженными протекторными свойствами и повышают устойчивость растений к низким отрицательным температурам. Предотвращение летального обезвоживания клеток при действии низких отрицательных температур происходит за счет накопления сахаров и других осмотиков, которые повышают водоудерживающую способность клеток и повышают устойчивость к охлаждению. Одной из главных защитных реакций является повышение осмотического давления в тканях. В многочисленных исследованиях показано, что уменьшение количества крахмала и повышение содержания сахаров в растениях связаны с морозостойкостью. Сахара, по-видимому, играют причинную роль в развитии морозостойкости. Сахара препятствуют образованию или уменьшают концентрацию токсических веществ, которые образуются при обезвоживании клеток посредством льдообразования; они являются основным субстратом для энергетических и биосинтетических процессов, происходящих в период закаливания при низких положительных температурах. Высказывается предположение о стабилизирующем влиянии сахаров на структурную организацию клеток, подвергающихся действию отрицательных температур. Изменения в структуре белков во время закаливания повышают их сродство к сахарам, связанные сахара способны защищать белки от денатурации во время обезвоживания. Показано, что при действии низких температур снижается удерживающая способность клеток, увеличивается выход электролитов, аминокислот и т. д. В период закаливания под влиянием сахаров увеличивается удерживающая способность клеток и уменьшается выход электролитов.
У морозостойкого сорта озимой пшеницы "Мироновская 808" повышение концентрации сахара резко снижает активность РНКазного белка, что ведет к большему сохранению рибонуклеиновой кислоты и лучшей регенерации растений в весенний период.
Высокая водообеспеченность плазмы приводит на морозе к возникновению большого количества льда, который может оказывать не только водоотнимающее действие, но и механически повреждать протоплазму. В литературе большое внимание уделяется вопросам состояния воды внутри клетки в связи с проблемой морозостойкости. Известно, что вода является основным субстратом, в котором развиваются реакции, управляющие метаболизмом клетки. Следовательно, от состояния и структуры молекул воды зависит уровень активности биохимических процессов. По мнению К. С. Тринчера, "структурные изменения клетки, сопровождающие физиологические процессы, это главным образом структурные изменения внутриклеточной воды, этого главного по массе компонента клетки" [48]. В большинстве литературных источников, опубликованных до 1966 г., преобладает мнение о том, что вся или по крайней мере большая часть воды, находящейся в цитоплазме, составляет единую структуру с биополимерами. С использованием методов диэлектрической спектроскопии, электронного парамагнитного и ядерного магнитного резонанса стали появляться работы, в которых высказывается суждение о том, что с биополимерами взаимодействует лишь небольшая часть внутриклеточной воды (гидратационная вода). Количество этой воды колеблется в зависимости от объекта и физиологического состояния в пределах 0,2 - 0,5 г/г или 20 - 50% от сухого веса. Общее же содержание воды в листьях растений составляет 200 - 400% от сухого веса, т. е. в 10 раз превышает количество гидратационной воды. По современным представлениям, негидратационная вода в клетках находится в виде двух микрофаз: решеточно-упорядоченной (или типа кластеров) и плотноупакованной (свободной). Последняя фракция воды располагается в пустотах каркаса и по своим свойствам напоминает свободную воду. Для решеточно-упорядоченной структуры воды такие показатели, как энергия, плотность, подвижность и координационное число, меньше, чем для плотноупакованной. Эти фракции воды не являются стабильными и непрерывно переходят друг в друга. По мнению А. М. Алексеева, важную роль в устойчивости растительных организмов к низким температурам играет соотношение между данными фракциями воды, что определяет степень ее подвижности внутри клетки. При температурах ниже 0°С состояние воды обусловлено в основном решеточо-упорядоченной структурой, с повышением температуры увеличивается доля плотноупакованной воды, достигая максимального содержания при +4°С [3].
В зависимости от степени подвижности внутриклеточной воды наблюдаются различные тины льдообразования в растительных тканях при действии низких отрицательных температур. В образовании внутриклеточного льда некоторые исследователи видят главную причину повреждения растений. Предполагается, что кристаллы быстрообразующегося льда разрушают интегральную структуру клетки, и гибель ее происходит вследствие механических повреждений протоплазмы.
По данным Г. А. Самыгина, в зависимости от скорости охлаждения обнаруживаются различные типы льдообразования: 1) при медленном замораживании лед образуется вне клеток, и они по большей части остаются живыми после оттаивания, гибель же некоторых клеток при этом происходит из-за их высушивания; 2) при быстром охлаждении вода замерзает между клеточной стенкой и протопластом, и в этом случае происходит повреждение поверхностных слоев протоплазмы; 3) при очень быстром замораживании вода не успевает выйти из протопласта и лед образуется внутри плазмы клетки, при этом клетки в основном погибают [42].
Таким образом, повреждение клеток при замораживании связано, с одной стороны, с образованием внутриклеточного льда, а с другой - с дегидратацией клеток. Лед образуется вначале во внешнем растворе вокруг клеток. Причем вероятность начала кристаллизации существенно повышается по мере переохлаждения. Для образования кристалла льда при -1°С необходимо, чтобы около 4 млн молекул воды заняли места в соответствии с кристаллической решеткой льда, а при - 30°С для данного процесса достаточно 100 молекул. Известно, что давление паров надо льдом меньше, чем над водой при той же температуре. В связи с этим возникшие центры кристаллизации начинают быстро расти. Для клеток опасен рост этих центров в более крупные кристаллы, т. к. их грани могут разрушать многочисленные мембраны клеток.
Если при замораживании эти кристаллики не успели вырасти до опасных размеров, то они могут это сделать во время оттаивания, когда в соответствующей температурной зоне происходит рекристаллизация. Достаточно медленное замораживание может полностью исключить кристаллизацию воды в клетке, но при этом неизбежно происходит значительная дегидратация и сжатие протопласта. Обезвоживание клетки происходит вследствие концентрирования внешнего раствора благодаря образованию в нем льда. При вымерзании воды концентрация солей и других веществ как вне, так и внутри клеток значительно возрастает, и данные соединения в этих условиях оказывают повреждающее действие на внутриклеточную структуру. Следовательно, если клетки не погибают в результате образования льда, то они могут погибнуть вследствие дегидратации и главным образом в результате чрезмерного осмотического стресса.
Дж. Левитт различал три периода, в течение которых возможны повреждения клеток и тканей при низких отрицательных температурах: периоды замерзания, оттаивания и сразу после оттаивания. Показано, что быстрое замораживание опаснее, чем медленное. Японским исследователям удалось охладить закаленные листья шелковицы до -210°С без повреждения, при условии, что вначале температуру снижали очень медленно. Процесс оттаивания тканей следует проводить крайне медленно. Замороженные ткани, температуру которых повышали до 0°С в течение 30 - 60 мин., повреждались слабее, чем ткани, у которых доведение до нулевой температуры осуществлялось в течение секунд. После оттаивания ткани необходимо помещать в растворы криопротекторов, в частности сахаров [62; 63].
Среди различных элементов клетки плазмалемму следует рассматривать как одну из основных мишеней при действии низкотемпературного фактора. Значение свойств плазмаллемы в проблеме устойчивости было выявлено классическими исследованиями Н. А. Максимова. Повреждение плазмаллемы в условиях низкотемпературного стресса приводит к повышению проницаемости, что ведет в свою очередь к нарушению водного режима растений, и листья после оттаивания теряют тургор, приобретая признаки увядания. Кроме того, в тканях затрудняется газообмен, так как межклетники остаются заполненными жидкостью. Выход ионов в межклетники нарушает ионный состав внутри клеток [35].
Следовательно, основными признаками повреждения растений от замерзания являются потеря клетками тургора, инфильтрация межклетников водой и вымывание ионов из клеток.
Многие исследователи считают ключевыми для понимания проблемы морозостойкости изменения структуры плазмалеммы [33].
Структура плазмалеммы крайне лабильная и чувствительная ко всем изменениям внешних условий, в частности, к снижению температуры и обезвоживанию клеток внеклеточным льдом. В процессе охлаждения вязкость липидного бислоя мембраны значительно повышается. Низкотемпературные фазовые переходы их могут вызвать необратимые структурные нарушения мембран и гибель клеток. Показано, что фазовые переходы липидов мембран из жидкокристаллического состояния в кристаллическое снижают проницаемость липидных мембран, а это увеличивает опасность замерзания воды внутри клеток, так как вода из них будет медленнее двигаться через клеточную мембрану к центрам кристаллизации вне клеток. Адаптация к низким температурам достигается снижением вязкости липидного компонента мембран путем изменения его состава, замены в составе липидов насыщенных жирных кислот ненасыщенными, в результате чего повышается показатель ненасыщенности липидов [21]. Это в свою очередь приводит к снижению температуры фазовых переходов липидов, что и защищает растения от образования внутриклеточного льда. Известно, что при выдерживании растений в условиях закаливающей температуры ненасыщенность жирных кислот возрастает. Такая реакция на закаливающую температуру является общей и проявляется в одинаковой степени как у морозостойких, так и у менее устойчивых сортов и видов. По-видимому, у растений в процессе адаптации к низким отрицательным температурам проявляются какие-то внутриклеточные механизмы, которые обеспечивают в этих условиях снижение температуры фазового перехода мембранных липидов и предотвращение внутриклеточного замерзания при скоростях охлаждения, встречающихся в природе.
Однако адаптация некоторых организмов (в частности, пшеницы) к низким температурам может осуществляться при неизменном уровне ненасыщенности жирных кислот.
Медленное промораживание растительных тканей приводит в первую очередь к инактивации систем активного транспорта и усилению пассивного выхода из клеток ионов и сахаров.
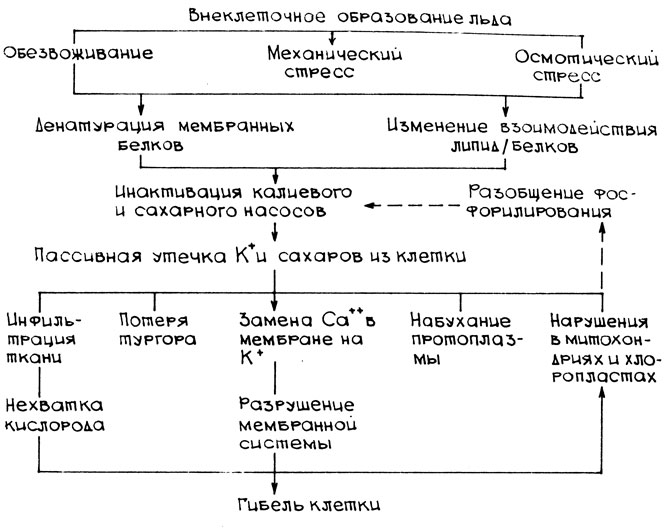
На схеме показана возможная последовательность событий, приводящих к гибели клеток вследствие повреждений, вызванных медленным внеклеточным льдообразованием с последующим медленным оттаиванием, которая приведена в работе Дж. П. Палта и П. Х. Ли [37].
Действие низких отрицательных температур приводит к нарушениям работы K+ -активируемых АТФаз плазмалеммы, тонопласта и механизма активного транспорта сахаров. Это может быть следствием денатурации мембранных белков и (или) изменений взаимодействий липид/белок. При этом значительно возрастает пассивный выход K+ и сахаров из клеток через поврежденные клеточные мембраны. При оттаивании, когда лед тает в межклетниках, ионы и сахара движутся по градиенту их концентрации (вакуоль - внеклеточный раствор), так как активная транспортная система не способна накачивать их обратно в вакуоль, и происходит потеря тургора клетками. Высокая концентрация K+ во внеклеточном растворе ведет к инфильтрации ткани водой, набуханию протоплазмы, хлоропластов, митохондрий. Увеличивающаяся утечка Ca2+ из поврежденных морозом клеток приводит к значительным повреждениям мембранных структур.
В зависимости от степени повреждения клеток все эти изменения могут быть обратимыми или необратимыми. В период после оттаивания обратимо поврежденные клетки полностью восстанавливаются, в то время как необратимо поврежденные клетки погибают.
При замораживании и оттаивании обнаруживаются существенные изменения внутри клеточных органелл и связанных с ними физиологических процессов. Устойчивость озимых культур в основном определяется структурным состоянием растительных клеток.
Известно, что целостность митохондриальных мембран является главным условием существования электрохимического потенциала ионов водорода, связывающего дыхание и фосфорилирование. Однако структура митохондрий чувствительна к различным воздействиям. Методом электронной микроскопии М. Квитковской были обнаружены изменения в ультраструктуре митохондрий у всех морозоустойчивых растений (рожь, пшеница, пырей, тритикале и др.) после охлаждения их при температуре 0÷-4°С. Вначале при охлаждении растений обнаруживаются удлиненные митохондрии с хорошо развитыми кристами, а затем при температуре -4°С они обратимо набухают [9].
Для всех морозоустойчивых форм характерна более быстрая реакция митохондрий на охлаждение по сравнению с морозочувствительными растениями. Результаты, полученные Н. Б. Христолюбивой с соавторами, свидетельствуют о том, что в клетках высокоморозоустойчивого пырея набухание митохондрий происходит уже через 20 мин., а у менее морозоустойчивой пшеницы - только через 6 часов после охлаждения растений. Многие исследователи связывают данные изменения с липидными перестройками мембран митохондрий, в частности, с более высоким уровнем ненасыщенных жирных кислот в мембранах митохондрий у устойчивых растений по сравнению с неустойчивыми.
Кратковременное охлаждение морозоустойчивых и чувствительных растений вызывает разнонаправленные изменения энергетической эффективности митохондрий [9]. В частности, в ответ на одночасовое охлаждение неустойчивых к морозу форм озимых злаков возрастает энергетический контроль дыхания (рост сопряженности процессов окисления и фосфорилирования и энергетической эффективности дыхания). В митохондриях морозоустойчивых озимых злаков при кратковременном охлаждении обнаруживается ослабление сопряженности окисления и фосфорилирования. При этом у митохондрий, изолированных из озимой пшеницы, возрастает скорость дыхания в состоянии 4 (по Чансу-Вильямсу - нефосфорилирующее дыхание) и снижается величина дыхательного контроля. Эти изменения свидетельствуют о переходе митохондрий в низкоэнергетическое состояние. При этом следует отметить, что ослабление энергетического контроля дыхания у морозоустойчивых растений приводит не к снижению, а к заметному увеличению скорости фосфорилирования за счет повышения дыхания митохондрий. Подобные изменения активности митохондрий обнаружены у животных организмов при гипотермии. Сходство реакций митохондрий растений и животных на температурное воздействие указывает, что переход в низкоэнергетическое состояние, по-видимому, является "аварийным" механизмом, который действует на первых этапах гипотермии у высших эукариотических организмов. Включение такого механизма имеет важное биологическое значение, так как позволяет растениям обеспечивать энергией многие реакции, необходимые для осуществления процессов репарации внутриклеточных структур при охлаждении. Кроме того, частичное разобщение окислительного фосфорилирования при переходе митохондрий в низкоэнергетическое состояние приводит к генерации тепловой энергии, освобождающейся при окислении субстратов. В результате этого температура побегов морозоустойчивых злаков оказывается более высокой по сравнению с температурой окружающей среды. Переход митохондрий морозоустойчивых растений в низкоэнергетическое состояние после кратковременной (1 час) гипотермии обусловлен, по-видимому, действием свободных жирных кислот. Холод приводит к резкому увеличению (в 2 - 3 раза) количества этих соединений, причем более значительно увеличивается содержание ненасыщенных жирных кислот по сравнению с насыщенными.
Митохондрии неустойчивых к морозу растений переходят к низкоэнергетическому состоянию либо после длительного действия температур, либо вообще не переходят. Высокий уровень свободных жирных кислот при гипотермии приводит к резкому увеличению дыхания в состоянии 4 и некоторому снижению интенсивности дыхания в состоянии 3 (фосфорилирующее дыхание), при этом падает величина дыхательного контроля и снижается отношение АДФ/О.
Увеличение длительности охлаждения (2 - 6 часов) приводит к снижению окислительной и фосфорилирующей активности митохондрий морозоустойчивых злаков. Снижение окислительной активности обусловлено действием оксалацетатного ингибирования сукцинатдегидрогеназы, которое ограничивает окисление таких метаболитов цикла Кребса, как сукцинат, а-кетоглутарат, малат [9].
Таким образом, при гипотермии у морозоустойчивых озимых злаков происходят существенные изменения в функциональной активности митохондрий. Вызванное холодом частичное разобщение окисления и фосфорилирования реализует одну из нормальных ответных реакций митохондрий на гипотермию. При этом митохондрии переходят на менее интенсивный, но устойчивый режим их работы, который сохраняется при действии более низких отрицательных температур (-18÷-20°С). Полагают, что подобная реакция митохондрий на холодовое воздействие формируется у проростков озимых культур и закрепляется в онтогенезе. У неустойчивых к гипотермии растений при значительном охлаждении (-18°С) нарушается ультраструктура митохондрий, резко снижается интенсивность дыхания и прекращается фосфорилирование.
Важную роль в криоповреждениях играют мембраны эндоплазматического ретикулума и рибосомальный аппарат, так как нарушение данных структур приводит к значительному снижению синтеза и транспорта цитоплазматических белков. С помощью методов электронно-микроскопического анализа обнаружено, что при действии низких отрицательных температур, в частности при закаливании, в клетках увеличивается поверхность эндоплазматического ретикулума. Одновременно с этим наблюдается изменение комнартментации рибосом: в гиалоплазме уменьшается число свободных моносом и полисом и увеличивается доля мембранно-связанных рибосом. Учитывая то, что свободные и связанные с мембранами рибосомы продуцируют разные классы белков, можно полагать, что при термическом закаливании происходит новообразование белков с новыми свойствами. Многочисленные литературные данные свидетельствуют о том, что закаливание растений сопровождается значительным увеличением содержания водорастворимых белков. Продуцирование адаптивных форм белков и ферментов (изоферментов) позволяет устойчивым растениям более эффективно выполнять свои физиологические функции в условиях неблагоприятного температурного режима.
Одним из способов защиты растительных клеток от повреждений, связанных с действием низких отрицательных температур, является синтез больших количеств белков нового типа, содержащих SH-группы. Эти белки способны к гидратации. Гидратационная вода практически не замерзает, она удерживается вблизи этих молекул белка силами, которые предотвращают образование кристаллов льда. Таким образом, чем больше в клетке таких белков, тем устойчивее клетки к замерзанию. Как правило, такие белки синтезируются осенью, когда происходит закалка, а весной растения, по-видимому, используют их в своем метаболизме.
Состояние пигментной системы может быть одним из чувствительных показателей морозных повреждений. Первым крупным исследователем пигментной системы в экологическом аспекте был Н. А. Любименко [34]. Среди достижений последних лет следует отметить работы минской школы физиологов под руководством Т. Н. Годнева [15]. В них показано, что в молодых побегах ели при низких отрицательных температурах (-14°С) идет фотосинтез, но накопления хлорофиллов не происходит. Имеются данные о том, что даже у теплолюбивых растений (огурцы, фасоль, кукуруза) общее содержание хлорофиллов под влиянием охлаждения не изменяется.
В ряде работ обнаружена способность пигментной системы приспосабливаться к зимним условиям. Прочность связей в пигментно-белковолипоидном комплексе хвои ели весной с увеличением интенсивности и продолжительности освещения снижается, в то же время осенью при подготовке к зимним условиям преобладают формы пигментов, прочно связанные с белками и липидами. Прочность связи пигментов с липопротеидным комплексом может служить показателем термоустойчивости пигментной системы.
У хвойных деревьев действие температурного стресса зимой сочетается с повреждающим действием света. В хлоропластах хвои в первую очередь повреждается электрон-транспортная цепь, но эти повреждения обратимы. В хлоропластах зимующих растений увеличивается содержание каротиноидов, что следует рассматривать как защиту хлорофилла от зимнего фотодинамического эффекта.
Способность к фотосинтезу у древесных растений незначительно снижается в процессе закаливания еще до воздействия морозов. У адаптированной к холоду озимой пшеницы при отрицательных температурах (-5÷-9°С) на слабом свету (2 - 6 клк) фотосинтез достигает 10 - 20% от максимального уровня, наблюдаемого в оптимальных для роста условиях. Фотосинтез необходим устойчивым организмам не только осенью, когда при низких положительных температурах происходит синтез протекторных соединений, в частности сахаров, но и для перезимовки растений. При низких отрицательных температурах за счет фотосинтеза у озимых злаков происходит частичная компенсация материальных и энергетических затрат для поддержания жизнедеятельности растений в стрессовых условиях.
Таким образом, у устойчивых к морозу растений имеются защитные механизмы, в основе которых лежат определенные биохимические процессы. Устойчивость растений к гипотермии определяется: 1) повышением содержания сахаров; 2) увеличением количества полярных липидов и повышением уровня ненасыщенности их жирнокислотных остатков; 3) увеличением количества защитных адаптивных белков; 4) переходом митохондрий вначале в низкоэнергетическое состояние, а затем на устойчивый режим работы; 5) снижением оводненности клеток.
Морозостойкость не постоянное свойство растений. Она зависит от физиологического состояния организма и условий внешней среды. Растения, выращенные при относительно низких положительных температурах, более устойчивы, чем те, которые выращены при относительно высоких осенних температурах. Жизненный цикл развития озимых многолетних растений контролируется сезонным ритмом светового и температурного периодов. Они начинают готовиться к перенесению неблагоприятных зимних условий с момента ослабления ростовых процессов и в течение осени во время наступления пониженных температур. Повышение морозостойкости тесно связано с процессами закаливания, то есть постепенной подготовкой растений к воздействию низких температур.
Закаливание растений. И. П. Туманов открыл и теоретически обосновал явление закаливания растений. Теория закаливания послужила толчком к проведению исследований по выявлению защитных механизмов растений к морозу.
Озимые культуры, двухлетние и многолетние травянистые растения, в отличие от древесных, не входят в конце лета в глубокий покой, а сохраняют способность к росту в течение зимы, несмотря на вынужденную приостановку, вызванную условиями внешней среды. Неблагоприятные зимние условия вызывают у них ряд последовательных изменений - снижение водного потенциала, нарушение метаболизма и др. Повышение морозостойкости тесно связано с процессом закаливания, то есть постепенной подготовкой растений к воздействию неблагоприятных зимних факторов. Морозостойкость является приобретенным признаком в процессе подготовки к зиме. Озимые культуры при своевременном посеве и хороших дружных всходах осенью успевают закалиться и приобрести устойчивость для зимовки. Разница в степени морозостойкости растений до и после закалки часто бывает значительной. Сравним: зимой озимая пшеница и рожь выдерживают морозы в -20÷-25°С, а осенью без предварительной закалки могут погибнуть уже при 5° мороза. Способность к закаливанию у древесных и зимующих травянистых растений в период летней вегетации отсутствует и проявляется только во время наступления осенних пониженных температур. Закаливание способствует осуществлению процессов жизнедеятельности в условиях зимы, а также проявлению скороспелости и повышению урожайности сельскохозяйственных культур. Разные органы растений имеют неодинаковую способность к закаливанию, например, листья листопадных деревьев не обладают способностью к закаливанию, цветочные почки закаливаются хуже, чем листовые.
Н. А. Максимов и И. И. Туманов выделили у озимых злаков две фазы закаливания - световую и темповую [35; 50].
Первая (световая) фаза закаливания протекает на свету и при низких положительных температурах (лучше всего +5÷+7°C). В осеннее время начинается процесс закаливания под влиянием изменения окружающих условий - укорачивания длины дня и снижения температуры. Оптимальный срок первой фазы закаливания - около 2 недель.
В этой фазе за счет фотосинтеза образуются сахара, а пониженные температуры значительно снижают их расход на дыхание. Фотосинтез, как процесс в первую очередь фотохимический, значительно меньше зависит от температуры, чем биохимический процесс дыхания. В результате происходит значительное накопление сахаров, которые являются питательными, энергетическими веществами и выполняют защитные функции. За время первой фазы закаливания количество сахаров в растении возрастает примерно до 40 - 60% на сухую массу клетки.
В зоне действия закаливающих температур свет необходим не только для синтеза сахаров в процессе фотосинтеза, но и для осуществления фотохимических реакций регуляторного типа. Т. И. Трунова с сотрудниками обоснованно доказала регуляторное действие света при закаливании озимых злаков. Основным рецептором, воспринимающим свет, является лист, освещение которого в течение 5 мин. в сутки достаточно для образования светового импульса, который обеспечивает у узлов кущения злаков способность к закаливанию. При этом молекула фитохрома, поглотив квант света, переходит в возбужденное состояние, взаимодействует с мембраной клетки и вызывает изменение ее конформации. В результате изменяется проницаемость мембраны, ее заряд и активность связанных с ней ферментов. Все это может быть причиной изменения путей общего метаболизма клетки, ее энергетических возможностей и уровня внутриклеточного гормонального баланса. Кроме того, активный фитохром может оказывать непосредственное влияние на генетический аппарат клетки, превращая потенциально активные гены в активные и тем самым способствуя образованию новых мРНК и биосинтезу белков с новыми свойствами. Показано, что низкие температуры и сокращение фотопериода являются для растений сигналом к торможению или прекращению роста и стимулом для накопления ингибиторов. Вероятно с этих процессов начинается формирование свойства морозоустойчивости растений [8; 49].
Установлено, что непродолжительное выдерживание проростков озимого рапса на свету и (или) на холоде (+5°) приводит к повышению содержания ингибиторов (возможно АВК). Гормоны непосредственно влияют на свойства клеточных мембран. Известно, что АБК увеличивает проницаемость мембран, что важно для своевременного оттока воды из клеток. Абсцизовая кислота снижает ростовую активность и способствует накоплению в клетках защитных веществ, что приводит к повышению устойчивости растений.
Отсутствие света в какой-то степени можно компенсировать действием физиологически активных веществ, в частности 6-бензиладенина (6-ВА). В настоящее время показано, что АБК и цитокинины можно рассматривать в качестве адаптагенных агентов. Причем АБК и цитокинины способны увеличивать устойчивость и у незакаленных растений (на фоновых температурах) и повышать эффективность холодовой закалки. Адаптагенные свойства цитокининов прежде всего связаны с их способностью стимулировать биосинтез белка. Свет и низкие температуры могут не только изменять уровень физиологически активных веществ, но и оказывать влияние на свойства рецепторов гормонов, направляя метаболизм клетки на специфический, холодозависимый путь [49].
Дж. Левитт неоднократно отмечал, что для закаливания растений необходимы энергия (АТФ) и восстановители (НАДФНг).
Так, в листьях озимого рапса после воздействия холодом (+2°С) как на свету, так и в темноте происходит увеличение содержания АТФ и уровня "энергетического заряда":
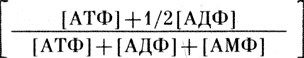
Это свидетельствует о том, что не только фотосинтез, но и процесс дыхания может способствовать обогащению клеток энергией в холодных условиях.
Повышение уровня энергетического заряда является важным фактором, контролирующим метаболические процессы, в частности, синтез белков и липидов в условиях пониженных температур. После воздействия низких температур (+2+5°С) в листьях озимого рапса происходит накопление специфических растворимых белков с низкой молекулярной массой (32 - 26 кД), обогащенных пролином, метионином и локализованных в цитозольной фракции. Их образование обусловлено, вероятно, синтезом de novo и в основном на 80 S рибосомах [24; 58; 59]. В исследованиях Т. И. Труновой при использовании ингибитора циклогексимида показано, что развитие морозоустойчивости озимой пшеницы в большей степени зависит от синтеза белка на 80 S рибосомах, локализованных в цитоплазме. Блокирование синтеза белка с помощью хлорамфеникола на 70 S рибосомах, локализованных в хлоропластах и митохондриях, в меньшей степени снижает способность пшеницы к повышению морозустойчивости. Данные, полученные А. Ф. Титовым, свидетельствуют о том, что закаливание растений сопровождается трансформацией электрофоретического спектра белков: содержание одних белков увеличивается, других - уменьшается и наблюдается появление новых белковых компонентов с иными свойствами. Следовательно, ядро через белковый синтез участвует в регуляции метаболической активности клеток в низкотемпературных условиях. Фракция растворимых белков представлена различными ферментами, и их накопление при закаливающих температурах имеет большое значение как для образования структурных элементов клетки, так и для реорганизации метаболизма при гипотермии.
Специфическим ответом растительных клеток на пониженные температуры среды является повышение уровня и изменение структуры фосфолипидов. Закаливание приводит в основном к повышению содержания ненасыщенных жирных кислот.
Результаты электронномикроскопических исследований свидетельствуют о том, что в течение первой фазы закаливания не происходит существенных изменений в ультраструктуре клеток. В этой фазе идут активные метаболические процессы, которые подготавливают биохимическую основу для структурообразовательных процессов, происходящих во второй фазе закаливания.
Вторая (темповая) фаза закаливания происходит после понижения температуры воздуха до -2÷-5°С. Длится она около двух недель при постепенном повышении отрицательных температур. Во второй фазе закаливания происходят физико-химические процессы, связанные с обезвоживанием клетки. Происходит отток из клетки воды, которая может превратиться в лед при низких отрицательных температурах. При постепенном замерзании растительные клетки и ткани обезвоживаются в результате образования внеклеточного льда.
В этой фазе происходит перестройка структуры протопласта: перемещение органелл внутри клетки и формирование развитой сети мембран. Возросший уровень белков и фосфолипидов в клетке обеспечивает новообразование мембран, особенно гладкого эндоплазматического ретикулума, и увеличение поверхности плазмалеммы за счет образования в ней складок и инвагинаций [26]. Для морозоустойчивых видов увеличение поверхности плазмалеммы и повышение ее проницаемости важно для своевременного оттока свободной воды из клеток в межклетники во время замерзания. При обезвоживании могут освобождаться вредные токсические вещества, но развитая сеть внутриклеточных мембран способствует их изоляции.
Обезвоживание клеток под действием низких отрицательных температур ведет к повышению концентрации солей и отрицательному влиянию их на биоколлоиды протоплазмы. Защита от повышенных концентраций солей лежит в усилении экзоосмоса части зольных веществ за пределы клетки. В результате этого происходит уменьшение отрицательного действия солей на биоколлоиды протоплазмы. Морозоустойчивые сорта выделяют в экзоосмосе больше зольных веществ по сравнению с неморозостойкими.
Особенностью морозоустойчивых форм растений является увеличение относительного объема протоплазмы и образование структурно-волокнистых структур в цитоплазме и в вакуолях. Эти структуры состоят из белков и других полимеров, они могут быть очень сложными (аналогичны микротрубочкам и микрофиламентам). Их увеличение характеризует переход структуры цитоплазмы из золя в гель (процесс гелификации). Значение гелификации состоит в улучшении механических свойств протоплазмы и в предотвращении внутриклеточного замерзания. Кроме того, гелификация делает содержимое клетки более инертным в химическом отношении и предупреждает резкие сдвиги в метаболизме. Растворимые белки, которые образуются при низких положительных температурах, используются во второй фазе закаливания для образования гелированных структур.
При отрицательных температурах у древесиных растений происходит гидролиз крахмала и других полисахаридов. Сахара не только понижают температуру клетки, но и выполняют роль пластификатора геля. Устойчивые растения (яблоня, береза) содержат осенью много сахаров, и их количество сохраняется в течение зимы на высоком уровне. Неустойчивые формы (дуб) также накапливают осенью много сахаров, но к декабрю - январю их количество резко падает.
После прохождения второй фазы закаливания растения становятся морозоустойчивыми. У них отмечаются значительные изменения в структуре воды. При низких отрицательных температурах наблюдаются процессы глубокого переохлаждения воды и процессы витрификации.
В литературе много данных о высоком содержании связанной воды у закаленных растений. У закаленных древесных растений и озимых злаков основная масса воды находится в незамерзшем состоянии. Если незамерзшая вода не является связанной, то ее сохранение можно объяснить процессом глубокого переохлаждения, который представляет собой определенные фазовые переходы молекул воды. Оно может происходить в клетках при снижении температуры до - 35÷-40°С. При дальнейшем понижении температуры (ниже -40°С) в данных клетках обнаруживается процесс льдообразования. У многих видов плодовых растений (яблони, груши) некоторые ткани (сердцевинные лучи древесины, зачатки цветочных почек) избегают внутриклеточного замерзания вследствие глубокого переохлаждения воды. Возможность образования переохлажденной воды в клетках обусловлена: особенностями их субмикроскопической структуры в закаленном состоянии, в частности, гелированием структуры цитоплазмы; частичным обезвоживанием этих клеток; отсутствием центров гетерогенной кристаллизации льда как в цитоплазме, так и в клеточных стенках.
В лаборатории И. И. Туманова исследовали динамику замерзания разных участков закаленных к морозу озимых злаков. Было выявлено, что в живых зачаточных побегах большая часть воды сохраняется в состоянии глубокого переохлаждения, что позволяет этим, структурам выносить низкие температуры. Для сохранения в зачаточных побегах определенного уровня незамерзшей воды большое значение имеют свойства соседних частично обезвоженных тканей узла кущения, которые предохраняют зачаточные побеги не только от льдообразования, но и от обезвоживания. Это соответствует общим закономерностям сушки - обезвоженные внешние зоны препятствуют обезвоживанию центральных частей объекта. При подсушивании узлов кущения за счет внеклеточного льда зачаточные побеги сохраняют высокую оводненность. Это сохранение незамерзшей воды в зачаточных побегах необходимо рассматривать как адаптацию к морозу, так как исключается один из главных факторов - сильное обезвоживание клеток. Переохлажденные клетки не подвергаются структурным и функциональным нарушениям, и поэтому нет необходимости в явлениях репарации, что важно для быстрого роста зачаточных побегов весной [27].
Многие организмы погибают при температуре -40÷-45°С. Однако при очень быстром замораживании (вблизи точки замерзания воды) клетки могут переходить в состояние витрификации (стекловидное состояние), при этом они выносят температуры -180÷-200°С. Чтобы прошел процесс витрификации, необходимо, чтобы в клетках сохранилось небольшое количество воды. Если воды много, происходит процесс кристаллизации. Процесс витрификации может быть достигнут двумя путями - путем быстрого охлаждения вблизи точки замерзания воды в клетках и путем медленного, ступенчатого охлаждения с разной скоростью. Последний путь возможен у клеток, у которых существует процесс обособления протоплазмы.
Получить витрификацию чистой воды крайне трудно, так как для этого надо "проскочить" интервал температур от 0°С до - 150°С очень быстро. Практически такие скорости охлаждения создать невозможно. Клетка представляет собой раствор солей и других соединений, которые могут связывать воду. Например, одна молекула сахарозы может связывать четыре молекулы воды, причем гидратация увеличивается с понижением температуры. Вода, связанная в виде гидратных оболочек гидрофильными белками, моно- и олигосахаридами, молекулами гемицеллюлоз, не замерзает, и клетки при этом не обезвоживаются. Данные вещества (криопротекторы) обеспечивают возможность осуществления процесса витрификации, затрудняя кристаллизацию. При витрификации у организмов происходит почти полная (или даже полная) приостановка жизненных процессов, то есть клетки переходят в состояние анабиоза. При этом взаимосвязанность и взаимосогласованность реакций обмена веществ сохраняются, но все они постепенно и равномерно снижают свою скорость практически до нуля. Явления витрификации протоплазмы в определенной мере объясняют высокую морозостойкость у некоторых растительных клеток.
Обратный процесс перехода веществ из стекловидного в жидкое состояние называют процессом девитрификации, он должен проходить очень быстро вблизи точки плавления, иначе образуются внутриклеточные кристаллы льда, что приведет к гибели клеток.
Следует отметить, что существует много неясного в вопросах о температурных границах начала повреждения и гибели растений, а также в том, как формируется окончательная устойчивость у активно вегетирующих растений.
Одни исследователи (И. И. Туманов, О. Л. Ланге) считают, что повышение устойчивости формируется в ходе воздействия крайних, но еще не повреждающих температур [31; 50]. Другие (В. Я. Александров, В. Ф. Альтергот) полагают, что повышение устойчивости-ответная реакция на действие повреждающих температур [2; 5]. С. Н. Дроздов с соавторами на основании литературных и собственных экспериментальных данных пришел к выводу, что весь диапазон активной жизнедеятельности холодостойких растений можно разделить на три зоны: фоновую, закаливающую и повреждающую. В частности, показано, что озимая пшеница сорта "Мироновская 808" обладает высоким термоадаптивным потенциалом в области пониженных температур: фоновые температуры от 16° до 28°С; закаливающие температуры от 15° до - 12°С; зона холодового повреждения от -12°С и ниже. Температуры фоновой зоны не влияют на терморезистентность растений, при закаливающих температурах устойчивость повышается, а при повреждающих - снижается [18].
Реакция растительного организма на температуру каждой зоны характеризуется определенным комплексом физиологических изменений. Процесс закаливания нельзя сводить только к постепенному торможению физиологических процессов, это в первую очередь изменения во внутриклеточной структуре и обменных процессах. Существенные изменения наблюдаются в фосфорном и энергетическом обмене. Низкие температуры, индуцирующие холодовое закаливание, вызывают перестройку процессов дыхания: несмотря на снижение интенсивности поглощения кислорода, энергетическая эффективность дыхания сохраняется даже при температурах, близких к повреждающим. Показано, что в листьях канареечника и картофеля при действии закаливающих температур увеличивается содержание органических форм фосфора, особенно кислотнорастворимой фракции, куда входят основные макроэргичеcкие соединения (АТФ, АДФ, АМФ), повышается количество доступной энергии, определяемой "энергетическим зарядом".
Повреждающие же температуры вызывают следующие изменения в обмене веществ: увеличивается содержание неорганического фосфора, в уровень органического (кислоторастворимой фракции) фосфора снижается. При этом поглощение кислорода резко снижается, происходит разобщение процессов окисления и фосфорилирования, дыхательный контроль уменьшается. Эти эффекты являются, по-видимому, следствием нарушения структуры и функции мембран, которые, в частности, могут быть вызваны переходом липидов из жидкокристаллической в нефункциональную кристаллическую форму, а также изменением белковых компонентов мембран. Кроме того, действие повреждающих температур приводит к снижению содержания водорастворимых белков.
Следовательно, наблюдаемые физиологические различия в реакциях растений на действие закаливающих и повреждающих температур носят, как правило, не только количественный, но и качественный характер.
Существует тесная связь между морозостойкостью растений и ростовыми процессами. Переход к состоянию покоя всегда сопровождается повышением устойчивости, а от состояния покоя к росту - снижением. В связи с этим морозостойкость одного и того же вида меняется в течение года: летом она минимальна, осенью увеличивается, а в конце зимы и в начале весны снова снижается.
Покой в проблеме морозостойкости. После закаливания некоторые растения, в частности древесные формы, переходят в состояние осенне- зимнего покоя. Наиболее продолжительный покой наблюдается у клена, вяза, дуба, менее продолжительный - у ивы и березы, небольшой период покоя отмечается у сирени и спиреи. По продолжительности покоя плодовые растения также различаются: короткий период покоя отмечается у вишни, наиболее продолжительный - у яблони. Однако покой никогда не охватывает всех частей растений, и в разных частях он выражен в неодинаковой степени. В наиболее глубоком покое находятся молодые однолетние ночки (их трудно заставить идти в рост при оптимальных условиях). Многолетние ночки почти не имеют покоя или он выражен слабо. Паренхимные клетки стволов и ветвей находятся в слабом покое, они в любое время могут делиться и образовывать камбий.
Глубина покоя в течение осенне-зимнего периода не остается постоянной: она возрастает до января - февраля, а затем начинает падать. Знание покоя, его глубины важно при интродукции растений. Для каждого организма существуют четкие ритмы (чередование периодов покоя и активной вегетации). Важно, чтобы эндогенные ритмы физиологических процессов соответствовали ритмам изменений условий внешней среды. Растения северных широт приспособились к короткому лету и выходят из состояния покоя сразу же после незначительного потепления. Эта способность полезна для арктических форм, чтобы более полно использовать короткий вегетационный период севера. При перемещении этих форм растений на юг эта их особенность уже не является положительной. В условиях юга весенние потепления сопровождаются возвратами холодов, которые губительно действуют на распустившиеся почки. Кроме того, на юге северные формы не могут использовать более длительный вегетационный период и при высокой температуре уходят в покой, то есть у них не происходит закалки.
Наиболее значительные изменения при переходе растений в покой происходят в протоплазме клеток. П. А. Генкелем и Е. З. Окниной (1964) было открыто своеобразное явление, получившее название процесса обособления протоплазмы, которое происходит осенью после накопления запасных питательных веществ и при снижении температуры до -3÷-5°С. Плазмодесмы, соединяющие протопласты соседних клеток, разобщаются и втягиваются внутрь. Поры в клеточной стенке, через которые проходят плазмодесмы, заполняются фосфатидами, и клетки оказываются изолированными (рис. 8). Обособление протоплазмы, которое происходит при значительном обезвоживании клеток, приводит к их физиологическому разобщению, нарушению взаимосвязи между ними. Этот процесс сопровождается изменением физиолого-биохимических процессов внутри клетки. Запасные питательные вещества, отложение которых предшествует обособлению, образуют сложные белково-липидные комплексы. Изменения в клетке идут в сторону увеличения гидрофобности и уменьшения содержания свободной воды. Количество связанной воды в зимний период возрастает [13].
Наличие обособления протоплазмы полностью сводит на нет отрицательное действие кристаллов внеклеточного льда, давящих на оболочку клетки, так как протоплазма уже вышла из клеточных стенок. Кроме того, обособление протоплазмы ведет к прерыванию путей кристаллизации. Отсутствие протоплазматических связей между клетками препятствует процессу кристаллизации воды, который может быстро распространяться при наличии плазмодесм. Таким образом, обособление протоплазмы - это комплекс сложных физико-химических и биохимических процессов, которые приводят к разобщению клеток, уменьшению свободной и увеличению связанной воды, росту вязкости протоплазмы и количества белково-липидных соединений. Все эти изменения в конечном счете приводят к уменьшению напряженности физиологических процессов. Снижение интенсивности физиолого-биохимических процессов и, главное, приостановка роста, то есть значительное сокращение энергетических затрат, позволяют без повреждения пережить неблагоприятные условия в зимний период. Следует отметить, что не все исследователи связывают процесс обособления протоплазмы с морозостойкостью.
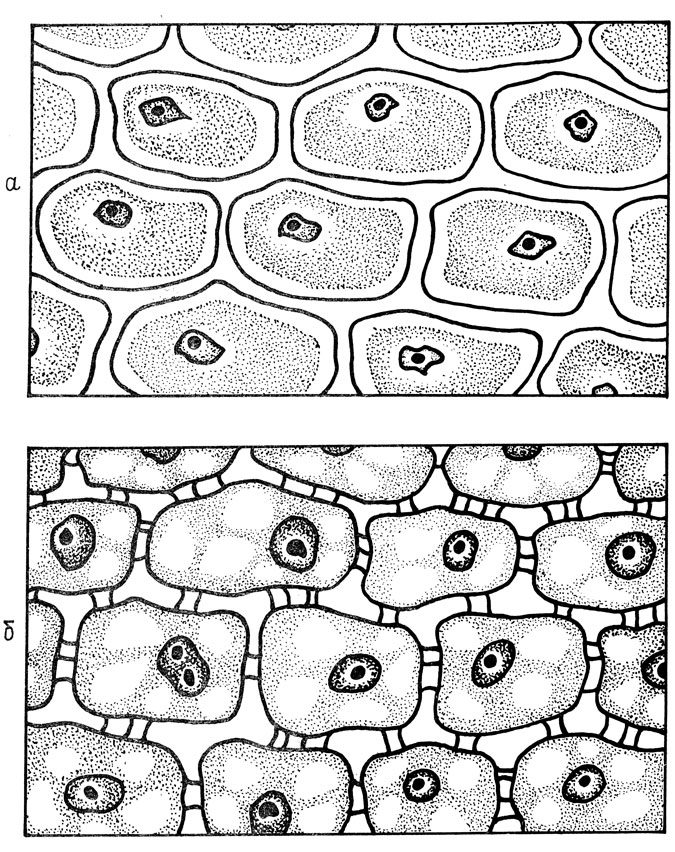
Рис. 8. Обособление протоплазмы в клетках почек яблони (по П. А. Генкелю, Е. З. Окниной): а - обособление протоплазмы в период покоя; б - отсутствие обособления в процессе роста
Таким образом, у растений имеются два основных физиологических состояния - вегетирующее и зимостойкое. Активный рост и высокая зимостойкость несовместимы друг с другом. В теплое время идет активный рост, а в холодный период постепенно формируется свойство зимостойкости. Вхождение древесных растений в состояние глубокого покоя является одним из условий формирования значительной устойчивости.
Покой - это период скрытного, подготовительного роста. В это время происходит активный процесс перестройки белковых структур, нуклео- и липопротеидов клетки, обусловливающий значительное обновление клеточных структур, что и позволяет растениям осуществлять усиленный рост в весеннее время.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'