Грибы, болезни растении и устойчивость к болезням
Большинство болезней растений вызывается паразитными грибами. Для того чтобы вызвать инфекцию, спора одного из таких грибов (довольно мелких, бесцветных, нитевидных организмов, близких по своей природе к растениям) должна попасть на поверхность растения и здесь прорасти (рис. 15.7). Вырастающая из споры гифа проникает в растительные клетки, из которых гетеротрофный гриб добывает питательные вещества. При этом гриб вызывает в растении симптомы болезни, связанные либо с повреждением клеток, либо с действием грибных токсинов. У растений существуют различные механизмы для защиты от такого вторжения; в сущности, уже сам тот факт, что грибы, как правило, поражают лишь один какой-нибудь вид растений или группу родственных видов, свидетельствует о том, что другие растения устойчивы к ним в силу тех или иных причин. Даже среди восприимчивых видов часто можно обнаружить экземпляры растений, в какой-то мере устойчивые. Селекционеры используют это обстоятельство при выведении новых, болезнеустойчивых сортов методом отбора и скрещивания.
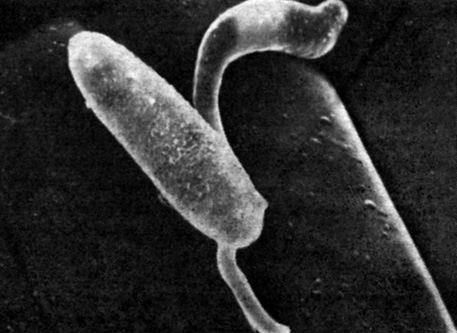
Рис 15.7. Электронная микрофотография (X5000; растровый электронный микроскоп) прорастающей споры (конидии) возбудителя ложной мучнистой росы ячменя (Erisyphe graminis hordei) на эпидермисе листа ячменя (Hordeum vulgare). Бороздки указывают границы клеток эпидермиса. Верхняя ростковая трубка плотно прижалась к поверхности во впадине, образованной клеточными стенками двух соседних эпидермальных клеток. Гриб проникает в эпидермальную клетку, которая находится позади кончика ростковой трубки. Клетка растения реагирует на это выделением с внутренней стороны клеточной стенки особых веществ, препятствующих заражению. Если заражение осуществилось, грифы гриба, ветвясь по поверхности листа, из клеток которого они добывают питательные вещества, образуют на листе налет, характерный для ложной мучнистой росы. (С любезного разрешения H. Kunoh, Cornell University.)
Болезнеустойчивость растений может иметь самую разную природу. Чаще других встречается так называемая структурная устойчивость. Если растение имеет толстую кутикулу, через которую прорастающая спора гриба не в состоянии проникнуть, то уже одно это может предотвратить заражение. Вторую линию обороны растений составляют различные химические их компоненты, опять-таки из числа вторичных метаболитов (которые, как мы уже отмечали, служат растениям также и для защиты от насекомых и прочих растительноядных животных). Для удобства эти химические соединения делят на две группы, в зависимости от того присутствуют ли они в растении еще до заражения (предсуществующие вещества) или образуются в нем в ответ на заражение. Эти две группы в какой-то мере перекрываются, но те вещества, содержание которых до заражения бывает очень низким и резко возрастает после заражения, принято относить ко второй группе.
Предсуществующие вещества
Многие предсуществующие вещества обеспечивают растениям некоторую степень устойчивости к паразитным грибам. Главную роль играют в этом, очевидно, сапонины (рис. 15.8), к которым принадлежат алкалоиды и тритерпены; в их молекуле имеется особым образом присоединенная гликозильная группа, придающая им водорастворимость. Среди защитных веществ много гликозидов этого типа; при отщеплении гликозильной группы их водорастворимость утрачивается. Эти соединения обладают выраженным антигрибным действием и, кроме того, способностью влиять на проницаемость и специфические транспортные свойства мембран. По-видимому, эти две их особенности как-то связаны, потому что сапонины заключены в вакуоли, и если гриб проникает в клетку, то клеточные мембраны повреждаются и высвобождают соответствующий сапонин. Нередко сапонин хранится в неактивной форме и активируется, только вступив в контакт с особым ферментом цитоплазмы. Если в мембране гриба присутствует фермент (3-гликозидаза, то он отщепляет от молекулы сапонина гидроксильную группу, в результате чего сапонин утрачивает водорастворимость и переходит теперь уже в липидную часть клеточной мембраны. Здесь он воздействует на некоторые стерины мембраны, переводя их из жидкого в более плотное состояние, - это повреждает клеточную мембрану гриба и убивает грибную клетку. Не все грибы, однако, в равной мере чувствительны к такому воздействию; у некоторых грибов в мембране отсутствуют соответствующие стерины, другие же обладают способностью инактивировать сапонин и вследствие этого сохраняют устойчивость к такого рода защите.
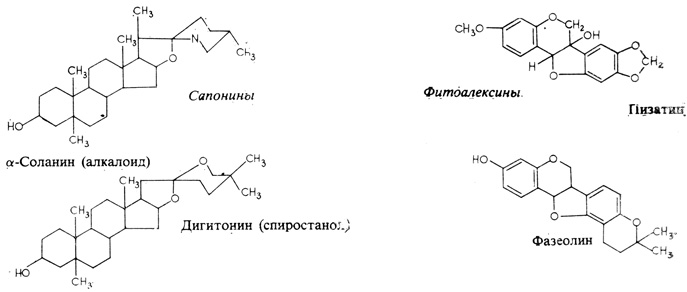
Рис. 15.8. Природные соединения, обладающие антигрибным действием
Почему растения, обладающие таким защитным средством, как сапонины, могут быть тем не менее восприимчивы к болезни? Дело в том, что эти механизмы, по определению, неэффективны против настоящих патогенов. Их роль заключается в том, чтобы не дать другим организмам стать патогенами. Таким образом, каждое растение поражается какими-то специфическими патогенными организмами и остается устойчивым к другим, которые могут в свою очередь поражать другие виды растений. Предсуществующие защитные вещества действуют главным образом на те грибы, которые сильно повреждают клетки растения-хозяина и при этом высвобождают или же активируют эти защитные вещества. Паразиты, которым удается обосноваться в растении, часто вводят в клетку свои гаустории - небольшие отростки гиф; гаустории проникают сквозь клеточную мембрану и извлекают из клетки питательные вещества, не слишком ее повреждая и потому не вызывая в ней никаких защитных реакций. В других случаях успешный паразитизм обеспечивается устойчивостью гриба к антигрибным веществам или его способностью разрушать такие вещества, благодаря чему инфекционный процесс развивается беспрепятственно.
Фитоалексины
Фитоалексины - это антигрибные соединения, образующиеся в растении в ответ на его заражение грибом. Название "фитоалексины", означающее в переводе с греческого "отвращающие вещества", закрепилось в науке в то время, когда исследователи неправильно полагали, будто в подобных веществах проявляется иммунный ответ растения, равноценный иммунному ответу животных. В действительности же в этих веществах проявляется вовсе не специфический ответ на грибную инвазию, а некий общий ответ - на любое повреждение.
В настоящее время из широкого круга растений выделен целый ряд соединений этого типа. По своей химической структуре они оказались очень различными. Каждое из них, по-видимому, специфично для того растения, в котором оно образуется, путь же образования зависит от природы данного фитоалексина. Так, для образования пизатина (рис. 15.8), вырабатываемого в растениях гороха, не требуется особых изменений в метаболизме растительных тканей, потому что у представителей семейства Leguminosae, к которому принадлежит горох, широко распространены изофлавоноиды, по своей структуре близкие к пизатину. Очевидно, пизатин образуется из аминокислоты фенилаланина, которая при этом сначала дезаминируется под действием фенилаланин-аммиаклиазы с образованием коричной кислоты, а это - путь, общий для всех флавоноидов (рис. 15.9).
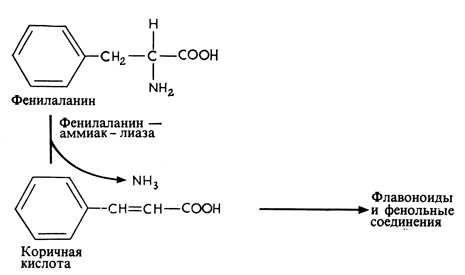
Рис. 15.9. Фенилаланин служит главным предшественником флавоноидов и фенольных соединений, многие из которых играют важную роль в устойчивости растений к болезням
Сможет ли данный фитоалексин сообщить растению устойчивость, это зависит от многих факторов. Чрезвычайно важно, насколько быстро растение распознаёт инфекционный агент и реагирует на него. Нередко фитоалексин начинает вырабатываться после заражения слишком поздно, когда гриб уже успел распространиться по ткани. В других случаях оказывается, что гриб обладает способностью разлагать данный фитоалексин и благодаря этому также распространяется. Гриб Botrytis cinerea чувствителен к фитоалексину конских бобов (Vicia faba) и поэтому ограничен в своем распространении местом заражения, так что на листьях он вызывает лишь появление мелких бурых пятнышек (откуда и само название этой болезни - "шоколадная пятнистость"). Родственный же вид гриба, Botrytis fabae, разрушает этот фитоалексин, что позволяет ему распространяться по всему листу.
На примере антракноза фасоли (Phaseolus vulgaris), возбудителем которого является гриб Colletotrichum lindemuthianum, можно продемонстрировать взаимосвязи, существующие между растением-хозяином, грибной инфекцией и образованием фитоалексинов. Устойчивость или восприимчивость различных сортов фасоли к этой болезни не удается связать с усиленным образованием фитоалексина фазеолина (см. рис. 15.8), которое наблюдается у них спустя 24 ч после искусственного заражения растений. Растения устойчивого сорта отвечают на внедрение гриба немедленной гибелью клетки-хозяина, так что ростковая трубка гриба, образовавшая на поверхности клетки шаровидный апрессорий, проникнуть глубже уже не может. Иначе ведет себя гриб в растениях восприимчивых сортов: ростковая трубка прорастает здесь сквозь стенку клетки-хозяина и продолжает расти между этой клеточной стенкой и протопластом, не причиняя клетке существенных повреждений. Таким образом гифы гриба растут на протяжении нескольких дней, распространяясь от клетки к клетке, по-прежнему без особых следов повреждения. Затем внезапно проявляются типичные симптомы антракноза: побурение, некрозы и гибель тканей. У устойчивого растения быстрое образование фазеолина начинается на второй день после заражения, одновременно с появлением мелких бурых пятнышек, свидетельствующих о сверхчувствительности. У восприимчивого растения фазеолин не вырабатывается до тех пор, пока на листьях не появятся типичные антракнозные поражения, а в это время фитоалексин помочь уже не может! Мы видим, таким образом, что, хотя этот фитоалексин, по-видимому, и способен сыграть свою роль в предотвращении болезни, существенное различие между устойчивыми и восприимчивыми сортами определяется их способностью распознавать патогена и соответствующим образом реагировать. И наоборот, специфичность болезни, очевидно, может зависеть от того, способна ли данная раса патогена избежать повреждения клеток (т. е. не спровоцировать ведущие к этому процессы или подавить их) и таким путем избежать ответной реакции растения - образования фитоалексинов. Возможно, следовательно, что реакция сверхчувствительности - это одна из главных реакций, обеспечивающих растению устойчивость к грибной инвазии. При сверхчувствительности быстро отмирает зараженная ткань, а часто и примыкающие к ней участки ткани, так что гриб, оказавшись в изоляции среди мертвых клеток, заразить другие клетки растения уже не может. В настоящее время это интереснейшее явление усиленно изучается.
Защита растений от болезней
Успешное, ведение сельского хозяйства предполагает снижение потерь от болезней растений или предупреждение этих болезней. Как и в мире животных, здесь существуют два подхода: профилактический и лечебный. Профилактические мероприятия там, где они возможны, обходятся дешевле и дают обычно лучший эффект. Различные агротехнические приемы также могут помочь предупредить болезнь; очевиден эффект, например, от уничтожения любого зараженного материала или от чередования различных культур в пространстве и во времени, с тем чтобы ограничить выживание и распространение патогена в посевах. Возбудители некоторых болезней распространяются при помощи других организмов (их называют переносчиками или векторами); в этих случаях желательно также вести борьбу с переносчиками.
Фунгициды. Эти препараты при их использовании либо создают защитное покрытие на поверхности растений, либо проникают внутрь растения и оказывают на него системное действие. Грибы, развивающиеся на растениях снаружи, подобно возбудителям ложной мучнистой росы, чувствительны к фунгицидам первой группы, и потому воздействовать на них несколько легче, чем на грибы, растущие внутри растительной ткани. Биологически грибы более удалены от человека, чем насекомые, так что фунгицидные вещества обычно не столь ядовиты для него, как инсектициды, но осторожность в их применении все же следует соблюдать. Все фунгициды можно подразделить на три главных класса. В первый класс входят давно известные неорганические соединения, в значительной мере вытесненные в настоящее время другими препаратами, но все еще употребляемые. К ним относится, например, молотая сера очень эффективное средство против некоторых грибов, развивающихся на поверхности растений. Широко применяется до сих пор также и бордоская жидкость - смесь гидроокиси кальция и медного купороса. При смешивании два этих соединения вступают в реакцию, в результате которой медь - активный ингредиент смеси - оказывается в более "доступном" состоянии; в этом состоянии она подавляет рост гриба, но не действует на само растение. Бордоская жидкость была впервые применена для того, чтобы предотвратить хищение винограда; однако при этом выяснилось, что она хорошо защищает виноград от мильдью, и с тех пор ее используют в качестве фунгицида. Второй класс фунгицидов - это органические соединения, действующие на поверхности растений, но более эффективные, нежели неорганические фунгициды. Важнейшую, наиболее разностороннюю и широко используемую группу составляют среди них органические соединения, содержащие серу (рис. 15.10). Очень эффективен, в частности, каптан, широко применяемый для протравливания семян с целью предохранить всходы от болезней, передающихся через семена.
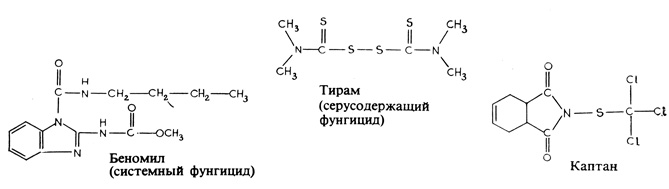
Рис 15.10. Синтетические фунгициды
Самым существенным достижением в области защиты растений следует считать появление третьего типа защитных веществ - фунгицидов или инсектицидов системного действия. Эти вещества проникают в растение и затем транспортируются по ксилеме или флоэме, поэтому они быстро ликвидируют грибную инфекцию или инвазию насекомых-вредителей в самых разных растительных тканях. Защитные вещества системного действия должны обладать такой химической структурой, которая позволяла бы им проникать в растение и двигаться по проводящим тканям вместе с транспирационным током или с током питательных веществ; эти два требования предполагают жирорастворимость и водорастворимость. Кроме того, вещества системного действия должны сохранять в растении относительную стабильность, в противном случае они не смогут обеспечить надежную защиту.
К наилучшим новым системным фунгицидам принадлежит беномил, обладающий широким спектром фунгитоксического действия. Наносить его удобнее всего на листья; введение в почву обеспечивает, правда, длительное поступление фунгицида через корни, но зато расход препарата при этом сильно увеличивается; инъекции в ствол дерева дают некоторый эффект при голландской болезни вяза, но полного излечения таким способом достичь не удается. Беномил транспортируется по ксилеме, поэтому из опрысканных старых листьев в молодые поступают лишь небольшие его количества. Это, конечно, некоторый недостаток данного фунгицида, поскольку новый прирост требуется опрыскивать заново. Однако это означает также, что фунгицид не попадает в плоды, так что с деревьев, обработанных до завязывания плодов, можно собирать плоды без всякого опасения. Механизм действия беномила связан с нарушением образования митотического веретена, т. е. этот фунгицид подавляет клеточное деление гриба. В целом беномил мог бы считаться почти идеальным фунгицидом, если бы не проблема возникновения устойчивости у патогенных грибов. На той стадии клеточного цикла, на которой действует этот фунгицид, одной генной мутации достаточно, чтобы сообщить организму устойчивость. Такие случаи известны, и число их постепенно растет. Приходится думать поэтому, что беномилу недолго суждено играть заметную роль в практической защите растений и что потребуется дальнейшая непрерывная работа над созданием новых соединений такого типа.
Бактерии и вирусы. Некоторые болезни растений вызываются бактериями и вирусами. Защита растений от бактериальных болезней полностью сходна с их защитой от патогенных грибов; устойчивость же растений к вирусам (не являющимся клеточными организмами) зависит от способности растительной клетки помешать вирусу воспользоваться клеточным аппаратом синтеза нуклеиновых кислот. В сельском хозяйстве защита от вирусных болезней достигается частично посредством изоляции растений от источника инфекции и частично с помощью борьбы с насекомыми-переносчиками вирусов (путем опрыскивания инсектицидами).
В садоводстве для избавления от вирусных инфекций есть два пути. Первый из них - это тепловая обработка растительных тканей. Растения, находящиеся в состоянии активного роста, выдерживают в течение длительного времени при 35 °С, а ткани в состоянии покоя прогревают в течение 30 мин при 55 °С; таким путем удается достичь разрушения вируса без заметного повреждения самой растительной ткани (хотя предел безопасности при этом довольно легко переступить). Более безопасный путь заключается в использовании меристематических культур (см. гл. 14), но этот способ пригоден лишь для отдельных видов растений. У некоторых растений, например у хризантем, отделенные концы побегов быстро растут в культуре. Размножение вируса отстает при этом от роста растительной ткани, вследствие чего верхушка побега оказывается через какое-то время свободной от вируса; эту верхушку отделяют и выращивают из нее целое растение, не зараженное вирусом.
Химиотерапия вирусных болезней растений делает пока еще только первые шаги, хотя и в этой области отмечен уже один явный успех. Речь идет о вирусе западной желтухи салата-латука. Листья растений, зараженных этим вирусом, желтеют и опадают; болезнь может уничтожить практически весь урожай. Однако если внести в почву препарат системного действия карбендазим, то он будет поглощаться корнями, что предотвратит вызываемое вирусом разрушение хлорофилла. Таким образом, хотя в клетках вирус и сохранится, листья салата-латука будут иметь вид совершенно здоровых, т. е. сохранят свои товарные качества. В будущем мы, возможно, научимся таким же способом вести борьбу и с другими вирусами.
Паразитические высшие растения. Паразитам из числа высших растений большого значения обычно не придают, однако при сильном поражении они могут извлекать из сосудистой системы растения-хозяина такое количество воды и питательных веществ, что недостаток их становится уже весьма ощутимым. Омела (Phoradendron flavescens), используемая для традиционного украшения дома на рождество, является опасным паразитом дуба и хвойных пород, способным существенно замедлять рост этих деревьев. Повилика (Cuscuta gronovii), вьющееся растение-паразит, тоже может причинять значительный вред, особенно в районах с более теплым климатом (рис. 15.11). Два паразита из этой группы поражают корни растений: стрига (Striga) встречается на субтропических злаках, таких, как кукуруза и сорго, а заразиха (Orobanche) паразитирует на многих видах растений, в том числе на бобовых культурах, томатах и табаке. В некоторых районах земного шара стрига составляет серьезную проблему в сельском хозяйстве. Низкие урожаи кукурузы в Северной Африке, приписываемые обычно бедности здешних почв, на самом деле часто объясняются тем, что посевы кукурузы заражены стригой. В США, особенно в средней части Атлантического побережья, грузы, предназначенные для морских перевозок, часто задерживаются карантинной инспекцией из-за того, что зерно содержит примесь семян этого корневого паразита. Стрига - неполный паразит; у нее имеются мелкие зеленые листья (обычно они видны у основания зараженного растения кукурузы). Поэтому стрига может в какой-то мере осуществлять фотосинтез, но все же она очень сильно истощает растение-хозяина. Борьба со стригой при помощи гербицидов не дала пока результатов. Главным звеном жизненного цикла стриги, на которое следует направить удар, можно считать прорастание семян. Семена стриги долгое время остаются в почве в состоянии покоя и прорастают лишь под стимулирующим воздействием некоторых соединений, вырабатываемых корнями растений-хозяев. Недавно удалось выяснить химическую структуру этих стимуляторов прорастания. Активное вещество, стригол, оказалось в некоторой степени сходным с гиббереллинами. Можно, очевидно, синтезировать это вещество и попробовать бороться со стригой, внося его в почву, чтобы вызвать прорастание семян стриги в то время, когда растений кукурузы на поле нет; молодые проростки паразита не смогут выжить без дополнительного питания, которое они получают от растения- хозяина, и будут, вероятно, обречены на гибель. Обрабатывать почву такими стимуляторами прорастания, для того чтобы уничтожить в ней семена стриги, придется, по-видимому, в течение нескольких лет подряд, потому что маловероятно, чтобы все семена этого паразита прорастали одновременно.

Рис. 15.11. Повилика (Cuscuta gronovii) - паразитическое высшее растение. Здесь изображена повилика, обвившая кустик утесника (Ulex europaeus)
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'