Циркадные ритмы
О любых событиях, происходящих с правильной периодичностью во времени или пространстве, говорят, что им свойствен ритм. Когда периодичность сохраняется и в отсутствие внешних воздействий, ритм называют эндогенным. Примером могут служить "сонные" движения некоторых растений, особенно из семейства бобовых (Phaseolus, Mimosa, Albizzia, Samanea и др.). Листья этих растений обычно днем бывают расположены горизонтально, а ночью - вертикально (никтинастия, рис. 12.1). Такие движения легко наблюдать, соединив лист тонкой ниткой с писчиком кимографа (рис. 12.2). Записи показывают, что листья начинают раскрываться до наступления светлого периода, а складываться - до наступления темноты. Таким образом, движения листьев, по-видимому, совершаются как бы в предвидении рассвета и сумерек, что указывает на действие каких-то других факторов помимо света и темноты.
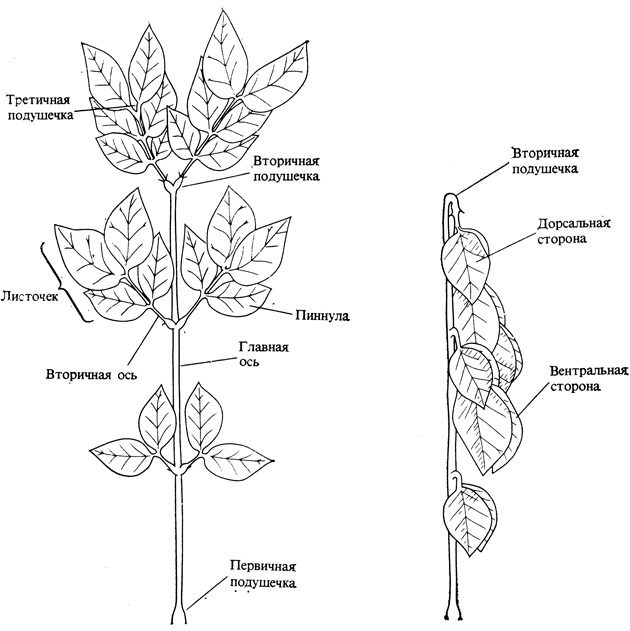
Рис. 12.1. Лист самана, Samanea saman, в дневном (слева) и ночном (справа) положении. Это тропическое бобовое растение имеет дважды перистые листья, состоящие из перистых первичных листочков, разделенных в свою очередь на вторичные листочки. Сонные движения регулируются изменениями тургора в клетках первичных, вторичных и третичных листовых подушечек, расположенных в основании листьев и первичных и вторичных листочков. (Satter et al. 1974. J. Gen. Physiol., 64, 413-430.)
Данные о существовании "внутренних часов" были получены в опытах, в которых лист несколько дней держали в темноте при постоянной температуре. Оказалось, что при этом ритмические движения продолжаются с периодом около 23 ч. Такие ритмы с периодами 20-30 ч (т. е. около суток) получили название циркадных*, от латинских слов circa - около и dies - день. Клеточный механизм, генерирующий ритм, обычно называют биологическими часами; положение листа в любой момент цикла показывает "циркадное время" (время, контролируемое внутренним осциллятором). Так как движения листьев у такого рода растений служат довольно точными индикаторами циркадного времени, их иногда называют "стрелками биологических часов" (рис. 12.3). Циркадные колебания этого типа встречаются по существу у всех эукариотических организмов, включая микроорганизмы, растения и животные.
* (В этом переводе сохранен привычный термин циркадные, хотя начинает уже входить в употребление более правильный вариант - циркадианные (который передает латинскую основу dies, в слове "циркадные" фактически пропадающую - от нее остаётся одна лишь буква "d"). - Прим. ред.)
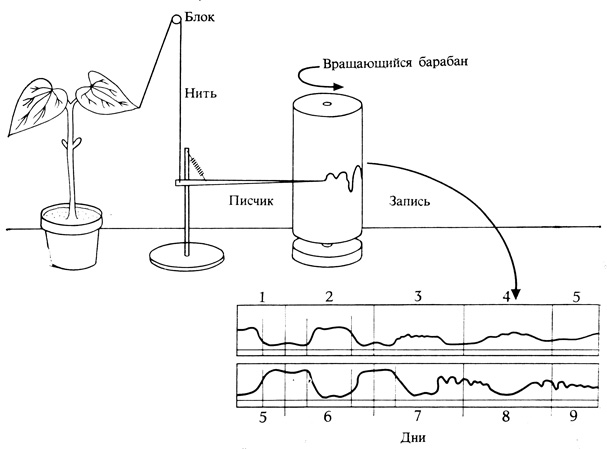
Рис. 12.2. Регистрация движений листа у бобов с помощью кимографа. Запись показывает движения во время циклов с 12 ч света (белые полоски внизу) и с 12 ч темноты (черные полоски) и в последующий период непрерывной темноты. Обратите внимание, что листья продолжают подниматься и опускаться и при постоянной темноте, однако перемена фаз свето-темнового цикла (нижняя запись) сдвигает фазу ритма на 12 ч
Природа осциллятора
Какова физическая основа этих эндогенных циркадных ритмов? Это трудно выяснить, так как каждая клетка гораздо сложнее, чем мы себе представляем, а весь организм - агрегат из многих клеток - еще более сложен. Некоторые аспекты этой сложности можно непосредственно видеть, например, с помощью электронного микроскопа; другие можно выявить физическими и химическими методами, как, скажем, при изучении структуры макромолекул белка и нуклеиновых кислот; о третьих удается заключить лишь косвенным образом, исходя из сложности поведения биологических систем. Именно с этой последней ситуацией мы и сталкиваемся, наблюдая эндогенные ритмы.
Чтобы узнать что-нибудь о природе осциллятора, многие исследователи пытались выяснить, какие условия могут изменить период (время, необходимое для одного полного цикла) или фазу ритма. Предполагается, что любой эффективный фактор нарушает внутреннюю работу часов. Если бы мы воспрепятствовали колебанию листа Samanea, неподвижно закрепив его на несколько часов, то после освобождения он быстро принял бы то же угловое положение, что и незакрепленный контрольный лист. Таким образом, механическое препятствие мешает проявлению ритма, но не влияет на отсчет времени часами. Напротив, изменения освещенности и температуры, а также некоторые химические вещества (ионы Li+, тяжелая вода, этанол) могут изменять фазу ритма. Обо всех эффективных в этом отношении веществах известно, что они влияют на свойства клеточных мембран. Это привело многих исследователей к мысли, что периодические изменения мембран являются составной частью "часового механизма". Мембранные поверхности окружают всю клетку и отдельные компартменты клеточного содержимого. Ритмически регулируя потоки метаболитов в клетку и из клетки, в органеллы и из органелл, биологические часы могли бы создавать ритмические изменения в химизме и физиологии клеток.
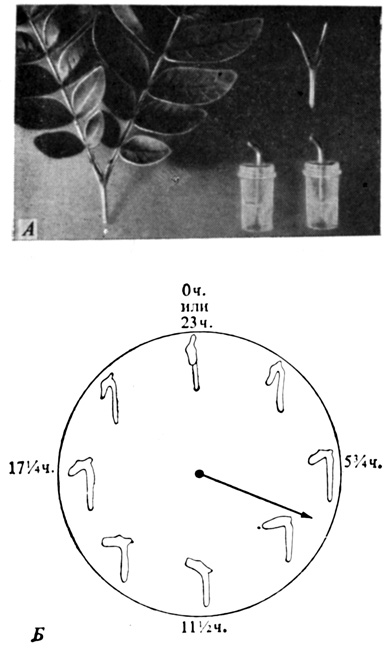
Рис. 12.3. Циркадный ритм движения листочков, регулируемый биологическими часами, у Samanea
Подобного рода изменения в структуре мембраны действительно наблюдали у одноклеточной водоросли Gonyaulax. По-видимому, встречаемость в мембране некоторых белковых частиц, наблюдаемых в электронном микроскопе после замораживания и скалывания, изменяется с околосуточной периодичностью. Однако мы не знаем, как генерируются эти ритмические изменения в структуре мембраны. Это ключевой вопрос, на который нужно ответить, если мы хотим понять природу биологических часов.
Измерение времени, независимое от температуры
Одна из интригующих особенностей циркадных ритмов - то, что длина периода в широких пределах практически не зависит от температуры (рис. 12.4). С функциональной точки зрения это неудивительно, так как любые часы были бы плохим измерителем времени, если бы скорость их хода зависела от таких внешних факторов, как колебания температуры! И все же трудно представить себе биологический механизм компенсации температуры; поэтому число принципиальных схем, которые логически могли бы быть предложены для объяснения ритмов, ограничено. Так как большинство метаболических реакций сильно зависит от температур, в одной из теорий предполагается, что в часах есть два разных процесса, подверженных влиянию температуры, один из которых при ее повышении ускоряется, а другой замедляется. В результате этой внутренней компенсации скорость хода часов может быть независимой от температуры.
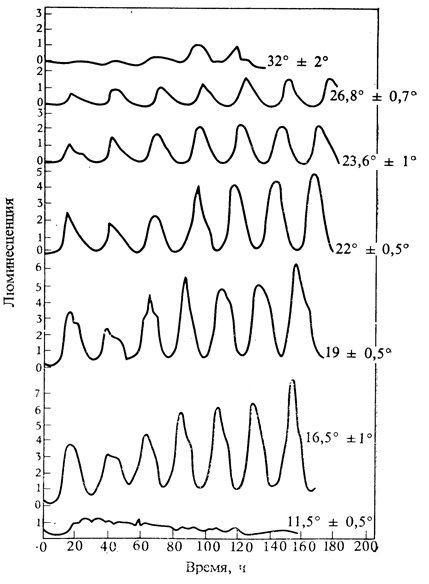
Рис. 12.4. Ритм возбуждения люминесценция у Gonyaulax polyedra на непрерывном свету (1000 лк) при разных температурах, указанных около каждой кривой. При повышении температуры с 16,5 до 26,5 °С период изменяется лишь на 12%. Очень низкие и очень высокие температуры подавляют ритм. (Hastings, Sweeney. 1957. Proc. Natl. Acad. Sci., 43, 804-811.)
Другая теория отводит главную роль липидам в мембране. Если липиды содержат жирные кислоты с длинной цепью, которые различаются по длине, а также по числу и положению двойных связей (см. рис. 5.11), то степень насыщенности и длина цепей жирных кислот могут регулировать текучесть мембраны. Более короткие цепи и ненасыщенность понижают температуру затвердевания жидких жиров (рис. 12.5). Изменения в жирных кислотах мембран происходят in situ в ответ на изменения температуры, способствуя поддержанию относительно постоянной текучести мембран в широком диапазоне температур (табл. 12.1). Некоторые исследователи полагают, что изменения в длине цепей и в степени насыщенности мембранных липидов происходят и на протяжении каждого суточного цикла и составляют часть механизма "часов". Если это так, то нам будет понятно, каким образом измерение времени могло бы быть относительно независимым от температуры.
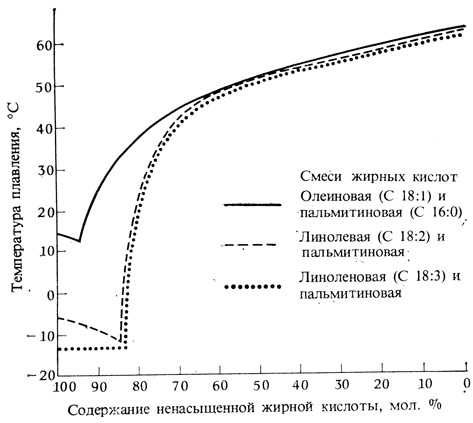
Рис. 12.5. Точка плавления смеси ненасыщенных и насыщенных жирных кислот (для каждой кислоты цифра слева от двоеточия указывает число углеродных атомов в цепи, а цифра справа от двоеточия - число двойных связей; структурные формулы см. на рис. 5.12). Обратите внимание на то, что полиненасыщенность снижает температуру плавления смеси. (По Lyons, Amundson. 1965. J. Amer. Oil Chemists Soc., 42, 1056-1058.)
Действительно ли ритмы эндогенны?
Некоторые авторы до сих пор полагают, что циркадные ритмы в действительности регулируются не эндогенными, а какими-то еще не выявленными внешними факторами. Против этого представления наиболее убедительно свидетельствуют следующие факты: 1) циркадные ритмы сохраняются и у организмов, быстро обращающихся вокруг Земли в искусственных спутниках; 2) при постоянных внешних условиях продолжительность периода не равна в точности 24 часам. В естественных условиях точная периодичность "навязывается" периодичностью в окружающих условиях. Это означает, что "естественный" период несколько изменяется под действием какого-то внешнего сигнала, "подгоняющего" внутренний ритм к ритму окружающей среды. У большинства организмов ритмы корректируются суточными изменениями освещенности (см. рис. 12.2) и температуры.
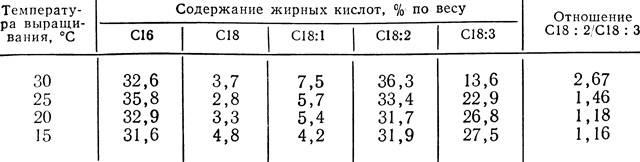
Таблица 12.1. Зависимость содержания различных жирных кислот от температуры)
(Жирнокислотный состав полярных липидов в кончиках корней проростков хлопчатника зависит от температуры. Обратите внимание на то, что растения, выращиваемые при 20 °С, содержат вдвое больше линоленовой кислоты (C18 : 3) и на 13% - меньше линолевой кислоты (C18 : 2) по сравнению с растениями, выращиваемыми при 30 °С. (St. John, Christiansen. 1976. Plant Physiol., 57, 257-259.))
Пигменты, служащие фоторецепторами для корректировки ритма, у разных организмов различны. У многих животных и некоторых растений это пигмент, поглощающий синий свет, но у большинства растений такую роль играет фитохром. Ритмы в этих растениях корректируются превращением Фк в Фдк. Можно было бы ожидать ежедневных взаимопревращений Фк и Фдк в растительной ткани вскоре после восхода и перед заходом солнца. Как видно из рис. 12.6, отношение энергии красного света к энергии дальнего красного света в солнечных лучах около земной поверхности составляет днем 1,3, но при заходе солнца быстро падает до 0,7. Это ведет к небольшому снижению содержания Фдк, которое и дальше продолжает снижаться в течение ночи в результате темнового превращения Фдк в Фк. Утром, вскоре после восхода солнца, уровень Фдк по мере увеличения упомянутого соотношения энергий опять повышается. Из этого понятно, каким образом повышение уровня Фдк с восходом солнца могло бы "подстраивать" биологические часы.
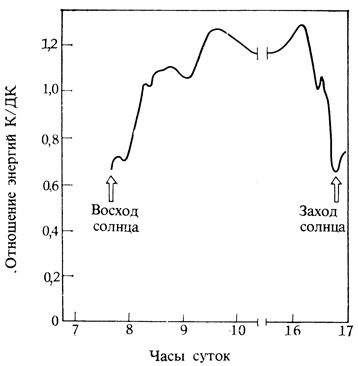
Рис. 12.6. Изменение соотношения энергий красного (660 нм) и дальнего красного (730 нм) света в солнечном спектре от зари до сумерек. Измерения были сделаны в Саттон-Бонингтоне (Англия), 52°50' с. ш., 22 января 1974 г. Восход солнца - в 7 час. 53 мин., заход - в 16 час. 32 мин. по Гринвичскому времени. (Holmes, Smith. 1977. Photochem. and Photobiol., 25, 533-538.)
Взаимодействие фитохрома с биологическими часами
Значительной частью наших знаний о биологических часах мы обязаны острой проницательности и тщательно выполненным опытам немецкого физиолога Эрвина Бюннинга. Наблюдая над 10 сортами сои, он заметил четкую корреляцию между их фотопериодической реакцией и сонными движениями листьев (рис. 12.7). Поэтому он предположил, что оба процесса регулируются одними и теми же внутренними часами. Из множества изученных им сортов формы с наиболее выраженными листовыми движениями были облигатными короткодневными растениями в отношении цветения, а остальные - большей частью нейтральными. По-видимому, те же определяемые фитохромами ритмические реакции, которые управляют движениями листьев, регулируют и фотопериодические процессы. От последних зависит, какая доля ресурсов растения будет направляться на вегетативный рост, размножение, создание резервов и процессы, ведущие к покою. Так как листовые движения менее сложны, чем фотопериодические явления, именно их изучали, чтобы подойти к анализу взаимодействий фитохрома с часами.
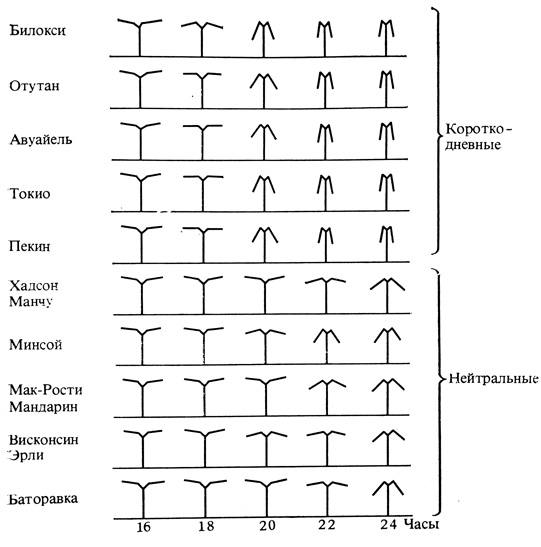
Рис. 12.7. Движения листьев 10 разных сортов сои с 16 до 24 час. Верхние пять сортов короткодневные, нижние пять - нейтральные. (Bimning. 1948. Z. Naturforsch, Teil B, 3b, 457-464.)
При никтинастии движения листьев обусловлены изменениями в объеме моторных клеток листовой подушечки - органа, находящегося у основания листовой пластинки (рис. 12.8). Когда листья раскрыты, моторные клетки, расположенные на одной стороне листовой подушечки, находятся в набухшем состоянии" а на противоположной стороне - в сжатом. Когда листья сложены, наблюдается обратная картина (рис. 12.9). Изменения в объеме моторных клеток в свою очередь регулируются массовым перемещением ионов K+ и Cl- в вакуоли и из вакуолей. Повышение концентрации K+ и Cl- снижает водный потенциал клетки, что ведет к усиленному поглощению воды и набуханию, а снижение концентрации этих ионов - к противоположному эффекту. Перераспределение K+ и Cl-, по-видимому, регулируется изменениями в мембранах моторных клеток.
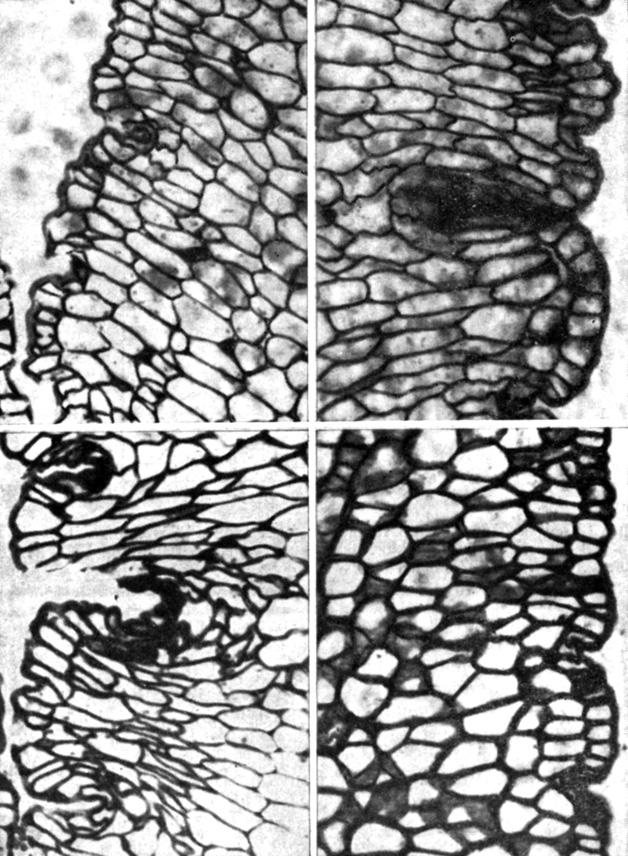
Рис. 12.8. Поперечные срезы третичной листовой подушечки никтинастического растения Albizzia julibrissin, сделанные днем (слева) и ночью (справа). Листочки у Albizzia ночью складываются кверху, что обусловлено сжатием вентральных (верхних) и набуханием дорсальных (нижних) клеток. Однако обратите внимание на то, что дорсальные клетки больше изменяют форму, чем размеры. (Satter et al. 1970. Amer. J. Bot., 57, 374-381.)
У никтинастичного растения Samanea, листья которого в период длительной темноты совершают колебания с околосуточной периодичностью, фазу колебания можно изменить путем изменения уровня Фдк. Количество фитохрома в форме Фдк, вначале определяемое спектральным составом света, предшествовавшего темноте, постепенно снижается по мере превращения его в Фк. Если период темноты прервать кратковременным воздействием красного света, превращающим Фк обратно в Фдк, поведение облученных и необлученных (контроль) листьев будет существенно различаться. Например, если листья облучить красным светом в тот момент, когда они раскрылись примерно на половину максимального угла, они начнут преждевременно складываться, в то время как контрольные еще продолжают раскрываться. Однако, если воздействие красным светом происходит на 12 ч позже, когда листья складываются, его эффект будет сравнительно небольшим. Таким образом, превращение Фк в Фдк в определенные фазы цикла "переводит" часы, тогда как в другое время оно остается безрезультатным.
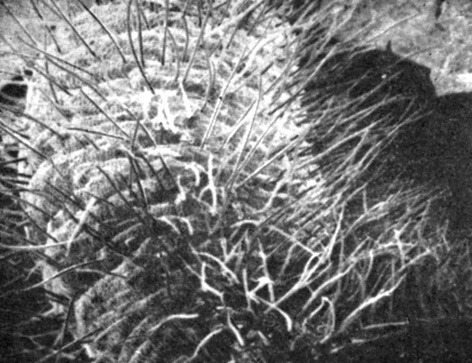
Рис. 12.9. Микрофотография вторичной листовой подушечки Samanea, полученная с помощью сканирующего электронного микроскопа. Обратите внимание на большое число волосков, выходящих из подушкообразных валиков, разделенных бороздками. Глубина бороздок изменяется по мере изменения конфигурации листовой подушечки
Эти взаимодействия можно объяснить, предположив, что и фитохром, и часы изменяют свойства одних и тех же мембран. Согласно этой теории, медленные циркадианные изменения в проницаемости и транспортных свойствах клеточных мембран происходят на протяжении каждого суточного цикла и являются частью самих часов, тогда как превращения Фк⇔Фдк приводят к более быстрым изменениям в структуре и функции мембран. Таким образом, мы можем видеть, как превращения фитохрома могли бы "переводить" часы, изменяя состояние клеточных мембран. Эта гипотеза объяснила бы также временные отношения, влияющие на способность вспышек красного света изменять ход часов, так как эффект превращения фитохрома зависел бы тогда от состояния мембраны в момент воздействия света. Это объяснение кажется разумным и логичным, но нам нужно знать гораздо больше о фитохроме и о часах, чтобы оценить, насколько оно верно.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'