Механизм флоэмного транспорта
За длительный период изучения флоэмного транспорта было выдвинуто множество теорий о его механизме. Большинство из них можно отвергнуть, так как они были основаны на опытах, вызывающих, как было показано позднее, возникновение артефактов. Что касается остальных теорий, то обсуждаемые в них данные можно удовлетворительно объяснить действием других механизмов, а не тех, которые предложили их авторы. Только одна теория, по общему мнению, согласуется почти со всеми известными фактами, а оставшиеся небольшие расхождения, очевидно, будут объяснены в дальнейшем путем незначительной модификации основной теории. Поэтому для простоты мы опишем лишь теорию перетекания растворов (или теорию тока под давлением Мюнха), предложенную первоначально немецким физиологом Карлом Мюнхом.
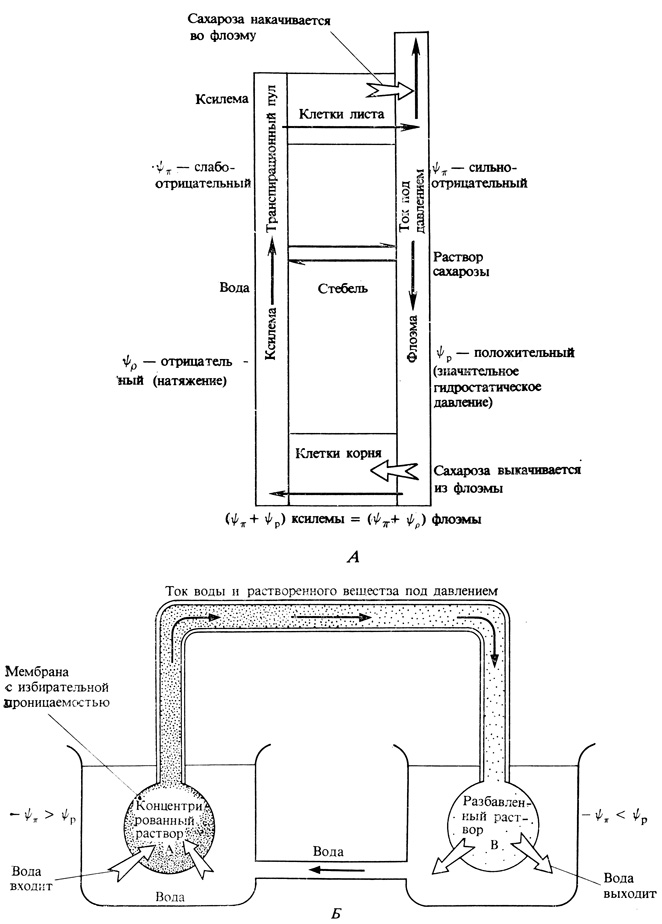
Рис. 8.5. А. Диаграмма, изображающая транспорт сахарозы в растении согласно гипотезе перетекания растворов. Б. Физическая модель гипотезы перетекания растворов
Согласно этой теории, перетекание растворов по флоэме полностью основано на простых принципах осмоса, что лучше всего можно показать, рассмотрев чисто физическую аналоговую систему. Две жесткие сферы, сконструированные из мембраны с избирательной проницаемостью, погружают в воду и соединяют между собой непроницаемой трубкой (рис. 8.5,5). Первоначально одна сфера заполняется концентрированным раствором сахарозы (А), а другая - разбавленным (В). В результате осмоса вода начинает поступать в обе сферы и в системе возникает давление. Поскольку давление возрастает в более концентрированном растворе А, оно будет передаваться по трубке к менее концентрированному раствору (В). Если давление, передаваемое от А, превысит давление, возникшее в В, то вода вместо того, чтобы поступать в В, будет вынуждена выходить из него. Так как вода в этом случае поступает в А и выходит из В, перетекание раствора сахарозы будет происходить от А к В. Оно будет продолжаться до тех пор, пока концентрации растворов А и В не выравняются. В этом состоянии вся система будет находиться под равновесным давлением и направленный нетто-поток раствора прекратится. Однако если мы будем добавлять сахарозу к А и удалять ее из В, то перетекание раствора сахарозы от А к В продолжится. Если при этом вода, удаленная из В, сможет через наружный раствор поступать обратно в А, то возникнет непрерывно действующая закрытая система. Для того чтобы показать это, растворы, окружающие А и В, соединены на рисунке трубкой.
В растении сахароза, образовавшаяся при фотосинтезе, активно накачивается в ситовидные трубки мелких листовых жилок в ходе процесса, называемого загрузкой флоэмы. Этот процесс приводит к снижению водного потенциала ситовидных трубок, и вода начинает поступать в них путем осмоса. В результате возникает давление, что вынуждает раствор перемещаться к потребляющим клеткам по градиенту давления. Из места потребления сахароза активно удаляется, что вызывает повышение водного потенциала ситовидных трубок. Затем вода диффундирует из ситовидных трубок по градиенту водного потенциала обусловленного главным образом наличием в них давления. Она быстро возвращается по ксилеме к источнику сахарозы.
Эта теория хорошо согласуется с основными известными о флоэме данными: флоэма находится под положительным давлением; между донором и акцептором существуют как градиенты давления, так и осмотические градиенты; содержимое ситовидных трубок перетекает по ним сплошным потоком; открытые ситовидные поры благоприятствуют такому перетеканию раствора. Остается одна проблема, заключающаяся в том, что, как известно, флоэмный транспорт требует более активного метаболизма, чем тот, который необходим для простого поддержания плазмалеммы ситовидных трубок. Эту дилемму можно решить, если принять во внимание тот факт, что метаболизм требуется для трех отдельных процессов: загрузки флоэмы в листьях сахарозой, переноса сахарозы к местам потребления и удержания ее внутри мембраны ситовидных трубок. Разное соотношение этих трех процессов определяет направление движения содержимого ситовидных трубок, расположенных вертикальными примыкающими друг к другу рядами.
Загрузка флоэмы
В листьях сахароза, первичный подвижный продукт фотосинтеза, должна поступать в ситовидные трубки против градиентам концентрации. Примечательно то, что листовые жилки ветвятся многократно до тех пор, пока диаметр их окончаний не оказывается равным толщине нескольких сосудов и ситовидных трубок. В этом месте они тесно примыкают к мезофильным клеткам, принимающим активное участие в фотосинтезе. У сахарной свеклы (Beta vulgaris) сахароза накачивается в эти концевые трубки непосредственно из стенок окружающих клеток мезофилла и мелких межклетников, куда она сначала транспортируется из клеток мезофилла (рис. 8.6). Транспорт сахарозы во флоэму избирателен и сопряжен с активным метаболизмом. Вероятно, при этом происходит совместное проникновение (котранспорт) сахарозы и водорода (H+) через специфическую пермеазу, присутствующую в плазмалемме ситовидных трубок, благодаря градиенту pH и электрохимическому градиенту. Позднее при разгрузке энергия необходима для откачивания ионов водорода из ситовидных трубок с помощью ATP-зависимого переносчика ионов H+. Этот фермент использует для перемещения ионов H+ через мембрану энергию, высвобождаемую при гидролизе АТР. Ионы водорода затем могут диффундировать назад в клетку вместе с сахарозой, используя специфическую протонно-сахарозную пермеазу и двигаясь по электрохимическому градиенту ионов H+.
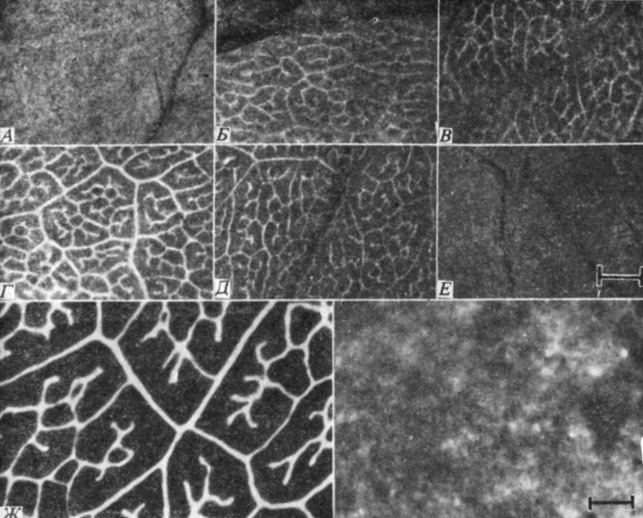
Рис. 8.6. Радиоавтографы, показывающие загрузку 14C-сахарозы в мелкие жилки листа сахарной свеклы после импульсного введения в него в течение 3 мин 14CO2; X1/2. Белый цвет соответствует меченым участкам листа. Листья были экспонированы в атмосфере 14CO2: А - 1,5 мин, Б - 5 мин, 5-8 мин, Г - 12 мин, Д - 20 мин, Е - 100 мин. Обратите внимание, что вначале фиксированные продукты распределяются по всему листу, затем они поступают в жилки и наконец выводятся из них. На фотографии Ж показан лист, который метился 14C-сахарозой в течение 180 мин. (С любезного разрешения D. R. Geiger, University of Dayton.)
Удержание сахарозы в ситовидных трубках в процессе транспорта
Во время транспорта по флоэме наблюдается высокоизбирательное удержание сахарозы в ситовидных трубках. Большинство других молекул могут свободно диффундировать как в ситовидные трубки, так и из них; при этом они находятся в состоянии равновесия с окружающими молекулами того же типа. Удержание сахарозы в ситовидных трубках происходит против значительного градиента концентрации и требует постоянной затраты метаболической энергии. Предполагают, что ситовидные трубки содержат большое количество ATP-азы, которая, возможно, участвует в этом процессе. Исходя из того, что клетки-спутницы выглядят очень плотными и изобилуют митохондриями, можно думать, что в них происходят высокоактивные метаболические процессы. Одна из их функций, вероятно, состоит в том, что они обеспечивают смежные ситовидные трубки энергией, необходимой для поддержания избирательной проницаемости мембран.
Несмотря на то что для поддержания данной системы требуется большое количество энергии, сам то себе транспорт в ситовидных трубках не зависит от метаболизма. Скорость транспорта изменяется в соответствии с уровнем потребления. Таким образом, метаболизм обеспечивает лишь удаление сахарозы и& флоэмы. Этот процесс вместе с метаболической загрузкой и поставляет энергию для непрерывного функционирования системы. Само перетекание растворов питательных веществ от их источников к местам потребления или запасания происходит по чисто физическим градиентам.
Следовательно, теория перетекания растворов может объяснить основные особенности флоэмного транспорта. Единственная структура, роль которой еще не понята, - это Ф-белок. Было высказано предположение, что микрофиламенты Ф^белка создают поверхность для транспорта веществ на границе раздела двух фаз, способствуя тем самым быстрому движению раствора по флоэме. Но до сих пор отсутствует экспериментальное подтверждение этого предположения. Хотя существуют, по-видимому, какие-то еще дополнительные механизмы флоэмного транспорта и теория перетекания растворов, вероятно, несколько изменится в будущем, в настоящее время она представляет наилучшее описание транспорта ассимилятов по ситовидным трубкам флоэмы.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'