Направление движения питательных веществ по флоэме
В ситовидных трубках присутствует не только сахароза, но и другие сахара, аминокислоты, гормоны и минеральные вещества, правда, в меньшем количестве. Если по ксилеме перемещение ее содержимого происходит исключительно от корней к листьям, то по флоэме отток наблюдается от донора (источника синтезируемых продуктов) вверх или вниз к любой точке, называемой акцептором, т. е. местом, где продукты фотосинтеза потребляются или запасаются*. Сахара необходимы для роста корней, верхушек побегов и молодых листьев. Они транспортируются также к корням и побегам, где откладываются в запас, или к семенам и плодам, где происходит их превращение в вещества, необходимые для роста последующего поколения. У некоторых видов акцепторами могут служить специальные органы запасания, такие, как клубни, корневища или луковицы. Когда, сахароза достигнет этих мясистых запасающих тканей, она обычно превращается здесь в нерастворимый крахмал, что предотвращает образование осмотически активных растворов с высокой концентрацией. Позднее крахмал в этих тканях переходит в сахарозу и экспортируется к новому месту потребления.
* (Фундаментальные данные о механизме транспорта ассимилятов по флоэме и их отложении в запасающих органах получены в результате 30-летней работы крупнейшего физиолога акад. А. Л. Курсанова и его учеников. Итоги их исследований суммированы в монографии: Курсанов А. Л. Транспорт ассимилятов. - М.: Наука, 1976. - Прим. ред.)
Перемещение сахаров по флоэме от донора к акцептору было продемонстрировано с помощью метода включения радиоактивной метки. Радиоактивную двуокись углерода или сахарозу наносили на лист какого-либо растения и через некоторый промежуток времени растение убирали, высушивали и помещали на рентгеновскую пленку. Там, где находился радиоактивный 14C, на пленке возникало изображение черного цвета. Таким путем легко выявляются части растения, которые получают сахарозу от подкормленного листа (рис. 8.1). Как правило, все потребляющие органы обеспечиваются ближайшим к ним доступным источником. Поэтому самые верхние фотосинтезирующие листья снабжают растущие почки и самые молодые листья. Нижние листья обеспечивают корни, а листья, находящиеся близко к плодам, - эти плоды. У многолетнего растения, у которого рост наблюдается главным образом в начале вегетационного периода, все листья снабжают сахарозой расположенные в разных местах запасающие ткани в конце сезона, обеспечивая создание больших запасов питательных веществ для следующего периода вегетации. Совершенно ясно, что движение веществ по флоэме не имеет определенного направления в отличие от их движения по ксилеме. В нижней части стебля это движение обычно направлено вниз к корням. В других частях стебля направление движения зависит от взаиморасположения донора и акцептора. Кроме того, направление транспорта может изменяться в зависимости как от возраста растения, так и от времени года.

Рис. 8.1. Движение 14C-сахарозы по растению бобов. Л. Радиоактивная сахароза синтезировалась в самом молодом, полностью зрелом листе (S2) из 14CO2, которую вводили в течение 2 ч на свету. Б. Затем растение расчленили на отдельные органы, высушили, положили на лист бумаги и поместили на несколько дней на рентгеновскую пленку. В. После проявления пленки и получения отпечатка радиоактивность выявляется в виде белого изображения. Небольшую часть обработанного листа экспонировали на пленке в течение более короткого промежутка времени для обнаружения более высокой концентрации 14C-сахарозы в жилках. Г. Движение от донора (обработанный лист) к разнообразным акцепторам показано стрелками. Междоузлие под листом-донором и самый молодой развивающийся лист (C) на той же стороне растения являются мощными потребителями фотосинтетических продуктов, идущих от обработанного листа. Самый молодой листочек (D) не метится, так как он получает сахарозу от листа-донора S1. Более старые листья тоже не метятся, поскольку они экспортируют, а не импортируют сахарозу. В то время как обработанный лист поставляет определенные вещества к корням, большинство ассимилятов, перемещающихся к корням, идут от самых нижних фотосинтезирующих листьев (S3, S4, S5). (С любезного разрешения D. R. Geiger, сотрудника University of Dayton, который провел данный эксперимент специально для этой книги.)
Исследования с использованием соответствующих меченых растворенных веществ показали, что транспорт по флоэме может происходить одновременно в двух направлениях. Эта видимая "двунаправленность" является результатом одностороннего тока в отдельных, но смежных ситовидных трубках, соединенных с различными донорами и акцепторами. Направление движения по флоэме определяется исключительно расположением места потребления питательных веществ относительно их источника. Вместе с транспортируемой водой и сахарозой от донора к акцептору движутся и другие соединения. Так, например, ростовые вещества, нанесенные на листья, перемещаются вместе с раствором сахарозы к месту ее потребления. Важность этого процесса иллюстрируется следующим фактом. Некоторые ростовые вещества, нанесенные на листья, находящиеся в темноте, не перемещаются за пределы листа. Но если на лист наносят сахарозу или освещают его, то ростовые вещества начинают перемещаться.
Четкое представление о передвижении веществ от донора к акцептору по флоэме важно иметь при использовании системных- пестицидов (инсектицидов, фунгицидов, гербицидов) и исследовании их эффективности. Системными называют пестициды, которые перемещаются в растении по ксилеме или флоэме либо по обеим системам одновременно. Пестициды, передвигающиеся только по ксилеме, не выходят за пределы листа, на который они нанесены. Они перераспределяются по всему растению лишь после их внесения в почву и поглощения корнями. Если инсектициды или фунгициды переносятся исключительно по флоэме, то они будут действовать эффективнее при распылении на листья. Такое защитное вещество будет перемещаться с потоком раствора сахарозы из листа к молодым развивающимся побегам и обеспечивать их постоянную защиту. Флоэмный транспорт обеспечивает также непрерывное передвижение защитных веществ от заканчивающих рост листьев к новым развивающимся побегам до тех пор, пока эти вещества не разрушатся растением. Чтобы гербицид с максимальной эффективностью действовал против сорняков с хорошо развитой системой вегетативно размножающихся столонов или корневищ, он должен перемещаться по флоэме. Такой гербицид передвигается из листа, места его нанесения, к акцепторам сахарозы в точках роста столонов или корневищ, что и приводит к их уничтожению. По существу одна из главных проблем, стоящая перед исследователями, создающими новые гербициды, заключается не в изыскании токсичных химикатов, а скорее в том, чтобы найти такой гербицид, который успевал бы доходить до точек роста столонов и корневищ, прежде чем он разрушит проводящую систему флоэмы.
Благодаря сочетанию ксилемного и флоэмного транспорта по растению циркулирует множество разнообразных веществ, в том числе минеральные элементы, азотистые соединения и растительные гормоны (рис. 8.2). Минеральные ионы, например, сначала поглощаются из почвы, а затем транспортируются в надземную часть растения главным образом по ксилеме. По мере старения листьев из них выводятся некоторые подвижные элементы (такие, как K+, H2PO4-, Mg2+), которые с потоком транспортируемой сахарозы переносятся по флоэме к акцептору. При недостатке питательных веществ эти элементы могут быть вновь направлены к растущему побегу, в результате чего он может остаться относительно здоровым, тогда как более старые листья истощаются и обнаруживают симптомы нехватки тех или иных, веществ.
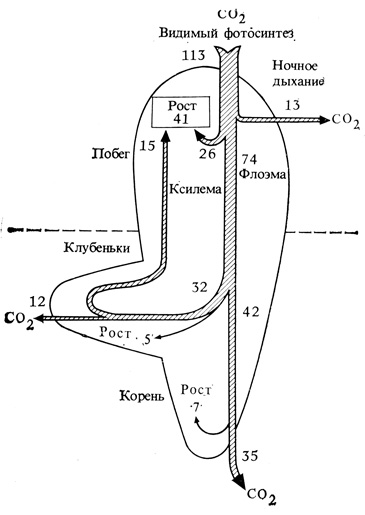
Рис. 8.2. Баланс углерода в растении гороха с корневыми клубеньками перед бобообразованием. Цифры обозначают проценты углеродных соединений, перемещающихся к каждому акцептору, в расчете на 100 единиц нетто-фотосинтеза. Обратите внимание, что некоторое количество фиксированного CO2 перемещается к корневым клубенькам, а затем возвращается к надземным органам в виде азотсодержащих соединений. С появлением плодов они становятся основными потребителями фотосинтетических продуктов. (J. S. Pate, Crop Physiology, Some Case Histories, L. T. Evans, ed., Cambridge, England University Press, 1975.)
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'