Адаптация к дефициту воды
Поскольку иссушение всегда представляет для растений главную угрозу, виды, произрастающие в засушливых местностях, располагают обычно какими-нибудь особыми механизмами, обеспечивающими их выживание в этих условиях. В пустынях дожди редко бывают настолько сильными, чтобы почва оставалась влажной хотя бы на протяжении .нескольких недель, т. е. в течение времени, необходимого для развития только что появившихся проростков; поэтому в пустынях семена многих однолетних растений не выходят из состояния покоя при слабых дождях и прорастают, только когда дождь достаточно продолжителен. Достигается такая регуляция прорастания различными способами. У одних видов семена имеют очень толстые, плохо проницаемые оболочки, которые пропитываются водой лишь при продолжительном дожде. У других в семенах содержатся водорастворимые ингибиторы прорастания - они вымываются из семян во время дождя; такие семена прорастают только после удаления большей части ингибитора, а это возможно лишь при дождях, пропитывающих почву медленно и основательно. Есть виды, у которых продолжительность обязательного периода покоя семян очень сильно варьирует, поэтому у них одновременно прорастает лишь очень небольшая доля семян; это увеличивает вероятность того, что хоть какие-то семена прорастут в то время, когда условия для плодоношения данного вида окажутся оптимальными. Однолетники пустыни после прорастания обычно растут быстро; репродуктивный цикл завершается у них иногда за несколько недель, так что к моменту, когда вокруг опять воцаряется сушь, растение уже успевает обеспечить себе непрерывность существования - образовать засухоустойчивые покоящиеся семена.
Для растительности пустынь характерен карликовый рост и малая площадь листовой поверхности, что снижает интенсивность транспирации. Уменьшают транспирацию и некоторые структурные приспособления: растения пустынь имеют обычно мелкие, толстые, сильно кутинизированные листья и малочисленные погруженные устьица (рис. 6.11). Самое замечательное, что устьица у некоторых ксерофитов открываются ночью, т. е. при слабой транспирации, и закрываются днем, о чем уже говорилось в гл. 4. Поскольку испарение воды сопряжено с охлаждением, можно было бы думать, что у ксерофитов с их низкой интенсивностью транспирации листья должны перегреваться, особенно в жаркие солнечные дни. Известно, однако, что у многих ксерофитов верхняя сторона листьев покрыта многочисленными мелкими волосками, отражающими лучи солнца, - благодаря этому тепловая нагрузка снижается (рис. 6.20). Сильная опушенность, таким образом, представляет собой приспособительный механизм; она очень часто встречается у растений засушливых мест и, видимо, служит растению для терморегуляции.
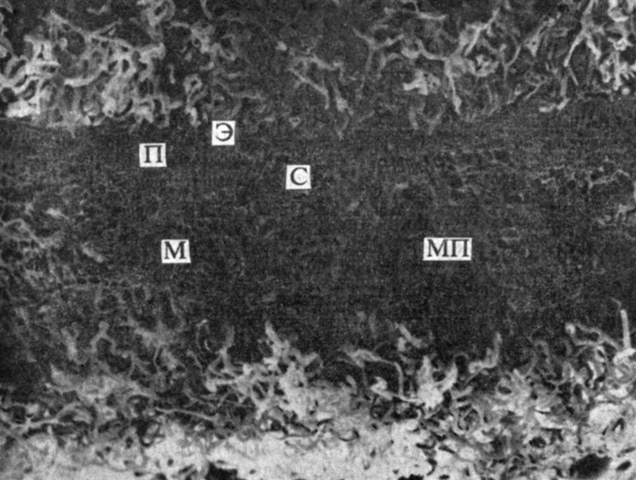
Рис. 6.20. Поперечный срез листа ксерофита Encelia farinosa; микрофотография, полученная при помощи сканирующего электронного микроскопа. (С любезного разрешения Ehleringer J., University of Utah.). Обратите внимание на обилие волосков с обеих сторон листа. С - сосудистый пучок; Э - клетка эпидермиса; П - клетка палисадной паренхимы; М - клетка мезофилла; МП - межклеточное пространство
Среди различных причин, вызывающих водный дефицит, важную роль играет засоленность почвы, потому что присутствие растворенных веществ снижает ψ почвенного раствора. В наше время засоленность почвы становится одной из серьезных мировых проблем. Причины этого в расширении орошаемых площадей, в усиленном применении удобрений и, наконец, в том, что с ростом населения неуклонно растет и число растворимых отходов. Поскольку корни растений поглощают воду из почвы преимущественно осмотическим путем, вода поступает в корни лишь в том случае, если ψ корневых клеток ниже, чем ψ почвы. Успешно растут на засоленных почвах растения с низкой величиной внутреннего ψ, которая обусловливается накоплением необычно больших количеств растворенных веществ в вакуолях корневых клеток. Если дать вызреть таким растениям, а затем убрать их с той почвы, на которой они росли, то пахотные ее свойства несколько улучшатся, потому что, извлекая из почвы соли, эти растения тем самым снижают ее засоленность. У многих растений этого типа имеются особые выделительные железки (рис. 6.21), через которые из растения активно выделяются большие количества солей. В некоторых случаях выводятся предпочтительно ионы натрия и хлора, которые могли бы подавлять клеточный метаболизм, а прочие ионы удерживаются, что поддерживает соответствующий осмотический потенциал. Ботаники стараются выявить как можно больше таких растений, способных извлекать из почвы соли, с тем чтобы их можно было использовать для мелиорации почв. При интродукции новых видов их водный баланс следует считать одним из главных факторов, определяющих их выживаемость и продуктивность, а значит, в конечном счете и ценность этих новых видов для человека.
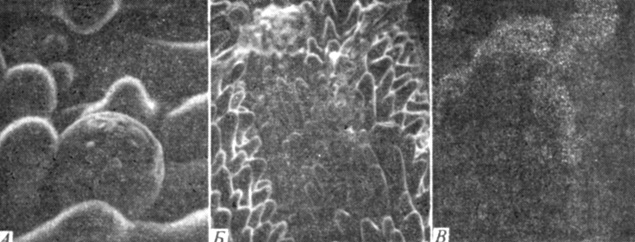
Рис. 6.21. А и Б. Микрофотографии выделительных железок на поверхности листа Distichlis spicata, полученные при помощи сканирующего электронного микроскопа. На фото слева (X1230) видно, что железка выделила небольшое количество соли, а на фото в центре (X500) видны многочисленные солевые выросты. В. Радиоавтограф, показывающий распределение хлорида в солевых выростах. Краткое описание радиоавтографии дано в подписи к рис. 6.14. (Hausen et al., 1976. Amer. J. Botany, 63, 635-650.)
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'