Материальная база фотосинтеза
CO2, используемая для фотосинтеза, попадает в зеленую" клетку листа или стебля через устьица и сильно разветвленную сеть межклеточных воздухоносных каналов. Лист (см. рис. 3.9) - главный орган растения, осуществляющий фотосинтез, - состоит из нескольких слоев активно фотосинтезирующих клеток (клеток мезофилла), окруженных защитным слоем (эпидермисом) и снабженных проводящими элементами (жилками), предназначенными для переноса веществ в двух противоположных направлениях: по жилкам вода и питательные вещества поступают в лист и по ним же ассимиляты и прочие продукты жизнедеятельности удаляются из листа. Жилки настолько сильно ветвятся, что в мезофилле практически нет ни одной клетки, которая отстояла бы от ближайшей к ней жилки дальше, чем на несколько клеточных диаметров.
Устьица, пронизывающие эпидермис (рис. 4.2), открываются и закрываются под влиянием изменений в тургоре замыкающих клеток. Каждое устьичное отверстие окружено двумя такими клетками, имеющими форму боба (рис. 4.3). Ясно, что способность замыкающих клеток к подобным движениям обусловлена их структурными и физиологическими особенностями. Стенки этих клеток весьма эластичны в отличие от жестких стенок, окружающих большинство зрелых растительных клеток. Кроме того, целлюлозные микрофибриллы, составляющие каркас клеточной стенки, ориентированы здесь не продольно, как это обычно бывает, а радиально. Далее, стенки у замыкающих клеток со стороны, обращенной к устьичной щели, толстые и не поддающиеся растяжению, а с противоположной стороны - более тонкие и более растяжимые. Вследствие этого, когда замыкающие клетки в результате усиленного осмотического поглощения воды становятся тургесцентными (см. гл. 6), они набухают и изгибаются, выпячиваясь наружу, так что устьичная щель открывается. Когда же эти клетки теряют воду, а следовательно, и тургор, они сжимаются, и устьичная щель при этом плотно закрывается. Сравнительно недавно удалось показать, что этот осмотический приток и отток воды, а значит, и тургесцентность замыкающих клеток регулируются интенсивным перемещением ионов калия, хлора и водорода между замыкающими клетками устьиц и другими клетками эпидермиса.
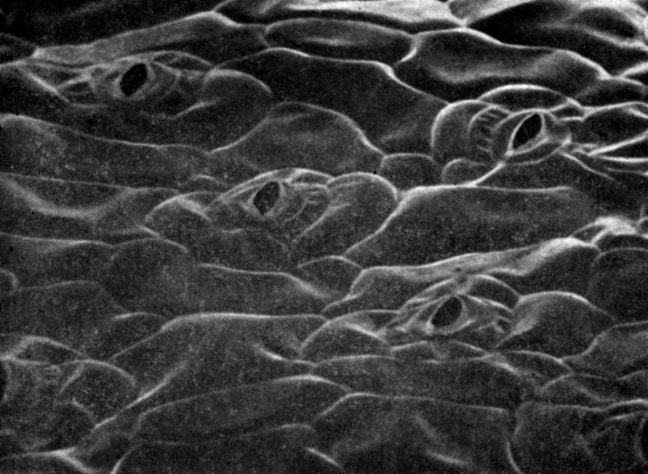
Рис. 4.2. Открытые устьица на эпидермисе нижней стороны лиофилизированных листьев Commelina communis. (С любезного разрешения H. Saxe, Copenhagen University.) Изображение получено при помощи сканирующего электронного микроскопа; X700
Пройдя сквозь устьичный барьер, атмосферная CO2 попадает в подустьичную воздушную полость, а затем в сообщающиеся воздушные ходы, по которым она диффундирует через весь мезофилл листа. На влажной клеточной поверхности CO2 растворяется в воде, гидратируется и превращается в угольную кислоту (H2CO3). Часть этой угольной кислоты нейтрализуется катионами клетки с образованием бикарбонат-ионов (HCO3-). Этот бикарбонат служит для клетки резервом потенциальной CO2, которую клетка может использовать для фотосинтеза.
Двуокись углерода, от которой мы все зависим, присутствует в атмосфере в следовых количествах: на десять тысяч частей воздуха приходится всего лишь около трех частей CO2 (0,03%). В разных местах земного шара эта концентрация несколько варьирует; она выше над городами, т. е. там, где сжигаются большие количества угля, нефти и бензина, и ниже в сельских местностях, где идет интенсивный фотосинтез. В теплицах в светлое время дня концентрация CO2 в воздухе также падает, а потому иногда ее здесь искусственно повышают, чтобы таким путем добиться усиления фотосинтеза у растений, достаточно хорошо обеспеченных и светом, и влагой. Правда, это таит в себе известную опасность: листья особо чувствительных растений могут при этом пострадать; быть может, это объясняется тем, что при высоком уровне CO2 в воздухе у некоторых растений закрываются устьица.
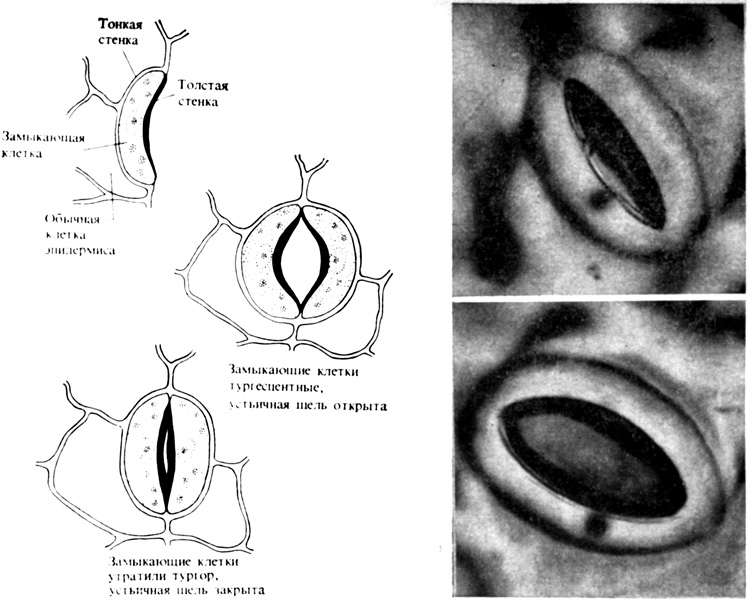
Рис. 4.3. Открывание и закрывание устьиц W. H. Freeman and Company.) На фотографии справа - изображения открытого и закрытого устьица табака, полученные методом наложения пленки из силиконовой резины. (С любезного разрешения I. Zelitch, Connecticut Agricultural Experiment Station.)
Эволюционисты высказывают мнение, что в сравнительно недавнее геологическое время содержание CO2 в атмосфере менялось довольно сильно и что эти изменения, возможно, сказывались на характере растительности и климате. Ясно, например, что повышение уровня CO2 в атмосфере могло бы привести не только к усилению фотосинтеза, а значит, и к усиленному накоплению растительной массы на Земле, но и к потеплению климата. Дело в том, что нагреваемая Солнцем Земля отдает часть поглощенной энергии обратно в космическое пространство в виде инфракрасного излучения. Двуокись углерода интенсивно поглощает именно инфракрасное излучение, задерживая тем самым эту тепловую энергию в атмосфере и создавая так называемый парниковый эффект (рис. 4.4). Нагревание Земли в результате такого эффекта могло бы привести к частичному таянию ледников и снеговых шапок на полюсах, а следствием этого явилось бы затопление равнинных местностей, на которых расположено большинство крупных городов. Таким образом, наблюдающееся сейчас поистине чудовищное потребление запасов ископаемого топлива и связанное с этим выбрасывание в атмосферу больших количеств CO2 могут иметь весьма важные последствия для человека. Правда, процесс, о котором идет речь, принадлежит, по-видимом,у, к саморегулирующимся, а возможно, даже, что это - циклический процесс. Повышение температуры и увеличение концентрации CO2 должно в конечном счете привести к усилению фотосинтеза и буйному росту растений, подобно тому что имело место в каменноугольном периоде. Такое усиление фиксации CO2 в процессе фотосинтеза со временем будет, конечно, заметно снижать уровень CO2 в атмосфере, что вызовет охлаждение Земли и обращение описанного цикла.
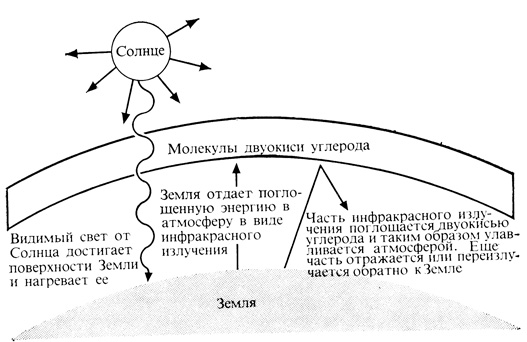
Рис. 4.4. 'Парниковый эффект', обусловленный присутствием CO2 в земной атмосфере
Количество воды, расходуемой на фотосинтез, составляет лишь очень малую часть от общего ее количества, поглощаемого и испаряемого растением. Для того чтобы фотосинтез происходил наиболее эффективно, листья должны быть тургесцентными и устьица открыты. Поэтому нехватка воды снижает скорость фотосинтеза, хотя и косвенно: процесс тормозится из-за того, что с закрыванием устьиц прекращается поступление CO2 в лист.
Выделяющийся в процессе фотосинтеза кислород попадает в окружающую среду через устьица, пройдя путь от поверхности клетки мезофилла до подустьичной полости по одному из связанных с ней воздушных ходов. Закрывание устьиц, вызванное ослаблением тургора замыкающих клеток, прекращает этот газообмен, но не подавляет полностью ни фотосинтеза, ни дыхания, поскольку и внутри листа эти процессы взаимно питают друг друга, будучи замкнуты в цикл, так что кислород или CO2, выделяющиеся в одном из них, поглощаются в другом. Фотосинтез, однако, в этих условиях (т. е. при закрытых устьицах) ограничен объемом дыхания, тогда как в оптимальных условиях он может происходить с интенсивностью, превышающей максимальную интенсивность дыхания в 10 и даже 20 раз.
Итак, для того чтобы фотосинтез протекал в оптимальном режиме, лист должен получать достаточно световой энергии, воды и двуокиси углерода. Если это условие не выполняется, то фактор, которого в сравнении с другими недостает больше всего, становится лимитирующим (рис. 4.5). Важно также, чтобы отток продуктов фотосинтеза из листа происходил с достаточной скоростью, потому что накопление углеводов будет тормозить процесс. Большинство растений лучше всего растет при чередовании световых и темновых периодов, так как в этих условиях продукты фотосинтеза, накопившиеся на свету, в темноте удаляются из листьев. Исключение составляют растения самых высоких северных и южных широт, которые должны успеть завершить вегетацию за отпущенный им короткий летний срок; растительность этих мест развивается наиболее хороша при непрерывном освещении.
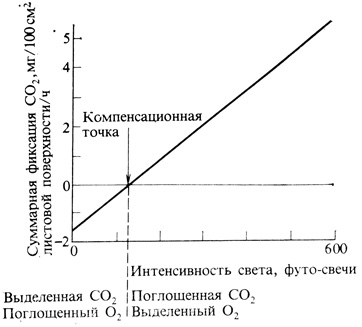
Рис. 4.5. Влияние света низкой интенсивности на фотосинтез типичного теневыносливого растения. В компенсационной точке (130 футо-свечей=0,7 Вт/м2) суммарный газообмен равен нулю: количество кислорода, выделившегося при фотосинтезе, равно количеству кислорода, поглощенного в процессе дыхания, а количество фиксированной в процессе фотосинтеза CO2 равно количеству CO2, выделившейся при дыхании
"Световое насыщение" фотосинтеза достигается почти у всех растений не на ярком солнечном, свету, соответствующем примерно 10 000 футо-свечей (400-500 Вт/м2), а при несколько меньшей интенсивности света. У отдельных листьев оно обычно отмечается при 1000 футо-свечей и ниже, в зависимости от вида растения, но, поскольку листья затеняют друг друга, для всего растения в целом световое насыщение наступает лишь при интенсивности света порядка нескольких тысяч футо-свечей (см. гл. 14). Растения, принадлежащие к так называемому теневыносливому типу, способны расти при сравнительно слабом освещении (рис. 4.6); им достаточно, например, света, проникающего сквозь полог леса. Естественно, что в затененных местах такие виды успешно конкурируют с растениями светолюбивого типа; светолюбивым видам для оптимального фото-синтеза требуются гораздо более высокие интенсивности света и потому в тени они растут плохо.
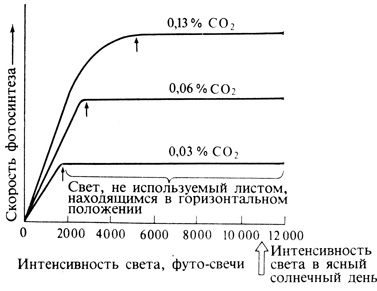
Рис. 4.6. Влияние света высокой интенсивности на фотосинтез при трех разных концентрациях CO2 (две из них превышают уровни, характерные почти для всех природных условий). Стрелки указывают световое насыщение. Слева от точки, отмеченной стрелкой, скорость фотосинтеза лимитируется светом
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'