Кинетика роста
Если высоту или массу растения представить как функцию времени, прошедшего с момента прорастания, то мы получим кривую, подобную той, какая изображена на рис. 3.4. Эта S-образная, или сигмоидная, кривая характерна для роста всех органов, всех растений, популяций растений или животных и даже для человеческих цивилизаций. Кривая состоит по меньшей мере из четырех четко разграниченных отрезков: а) начальной лаг-фазы, на протяжении которой происходят внутренние изменения, служащие подготовкой к росту; б) лог-фазы, или периода, когда зависимость логарифма скорости роста от времени описывается прямой; в) фазы постепенного снижения скорости роста и г) фазы, в течение которой организм достигает зрелости, рост прекращается и размеры стабилизируются. Если продолжить эту кривую, то в какой-то момент ход ее может вновь измениться, отражая период старения и смерть.
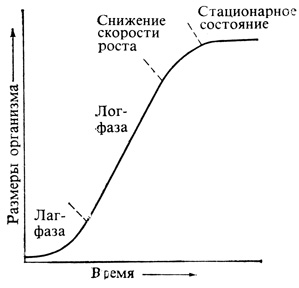
Рис. 3.4. S-образная, или сигмоидная, кривая роста. Такой тип роста свойствен отдельным клеткам, тканям, органам, организмам и популяциям
Существуют растения, для которых старение и смерть не являются обязательной частью жизненного цикла. В западной част Северной Америки произрастают остистая сосна (Pinus aristata) и секвойя, возраст которых может значительно превышать 3000 лет. Погибают эти деревья, по-видимому, от случайных причин - от удара молнии, от инфекции или же от того, что основание дерева в какой-то момент оказывается недостаточно прочным, чтобы выдержать тяжесть ствола. Если бы можно было как-то предотвратить подобные вредные воздействия, то, видимо, рост такого дерева был бы потенциально бесконечным. Это потенциальное бессмертие растительных клеток можно продемонстрировать методом культуры тканей - от целого растения отделяют какие-нибудь его части и выращивают их на искусственных средах. Французские исследователи в 1937 г. показали, например, что если кусочки корней моркови отделить в асептических условиях и поместить в соответствующую стерильную химическую среду, то через некоторое время в них возобновится клеточное деление и возникнет быстро растущая масса недифференцированных клеток, так называемый каллус. Если затем кусочки этого каллуса достаточно часто переносить на свежую среду, то он будет продолжать расти все с той же постоянной скоростью. Проследить такой рост удалось на протяжении сорока с лишним лет. Морковь в норме - двулетнее растение, и, значит, тот экземпляр, от которого некогда была взята ткань, должен был давным-давно завершить свой цикл развития. Естественно напрашивается вывод, что обычное прекращение роста какого-либо растения, за которым следуют старение и смерть, связано с каким-то ингибированием и что если это ингибирование тем или иным способом устранить или обойти, то мы получим растение, способное расти непрерывно, иными словами - потенциально бессмертное. Исследования в этой области продолжаются и, очевидно, рано или поздно дадут интересные и важные результаты.
Кривые роста ценны тем, что они свидетельствуют о существовании разных типов физиологической регуляции роста. По длительности лаг-фазы мы можем, например, судить о характере тех изменений, которые должны произойти, прежде чем начнется рост. У многих семян лаг-фаза длится не более нескольких часов, и это показывает, что здесь уже имеются в наличии все системы, необходимые для роста, и требуется только вода для активации. Известны, однако, и семена, у которых лаг-фаза измеряется сутками, неделями и даже месяцами. У таких покоящихся семян в тканях обычно присутствуют какие-то ингибиторы роста и прорастание не может начаться до тех пор, пока эти вещества не будут разрушены в процессе метаболизма или удалены каким-нибудь механическим способом - вымыванием и т. п. Скорость роста в лог-фазе нередко определяется гормонами, о чем мы еще будем говорить далее. По наклону кривой можно часто довольно успешно судить о генетическом фоне, определяющем ростовой потенциал данного растения, а также о том, насколько хорошо отвечают потребностям растения условия, в которых оно растет. Конечная высота растения и сроки наступления стационарной фазы часто также бывают заданы генетически, но эти характеристики могут в какой-то мере меняться под влиянием среды. Старение и смерть организма не обязательно жестко определяются его генотипом, их сроки тоже может определять среда.
Клеточное деление, растяжение и дифференциация
Растение - это сообщество разных клеток, возникших в результате клеточного деления и последующей дифференциации. Активно делящиеся, или меристематические, клетки, сосредоточенные в участках, которые называются меристемами (рис. 3.5), сохраняют на протяжении всей жизни растения способность непрерывно делиться и порождать новые клетки. Меристемы имеются на кончиках корней и на верхушках побегов у всех растений, а также в базальной части листьев и междоузлий у некоторых однодольных. Таким образом, у двудольных стебель растет за счет верхушки, тогда как у однодольных рост может происходить либо за счет верхушки, либо за счет базальной части стебля или междоузлия. Узлы, т. е. те участки стебля, от которых отходят листья, также могут содержать меристематические клетки. Между листом и стеблем, в пазухах листьев, закладываются пазушные почки. Пазушные почки, длительное время не дающие побегов, называются спящими; при определенных условиях они пробуждаются и из них развиваются побеги. Развитие боковых побегов подчинено гормональной регуляции, и мы еще будем говорить об этом позднее, в гл. 9 и 10. Рост в толщину у стеблей и корней двудольных происходит в результате тангенциального деления меристематических клеток камбия - образовательной ткани, расположенной однорядным цилиндрическим слоем, между ксилемой древесины и флоэмой коры. Возникающие в результате деятельности камбия клетки дифференцируются и образуют вторичную флоэму (кнаружи) и вторичную ксилему (кнутри).
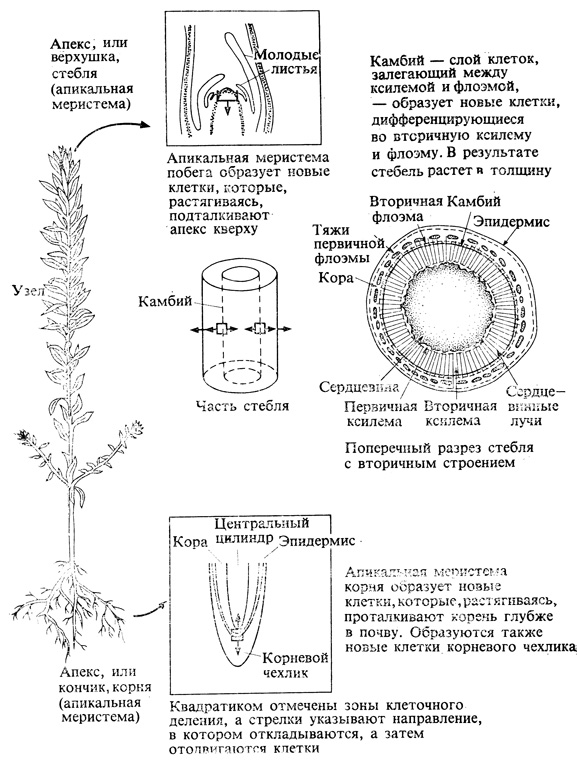
Рис. 3.5. Строение сосудистого растения. 1-е изд., New York, John Wiley and Sons.)
В стебле или корне растения, в этом высокоорганизованном сообществе клеток, каждая клетка в своем развитии проходит (в определенной последовательности) через ряд фаз. Меристематические клетки - мелкие, тонкостенные, приблизительно кубической формы с множеством мелких вакуолей и относительно большим ядром (рис. 3.6). Увеличиваются клетки (в основном вытягиваются в длину) главным образом в результате поступления воды в вакуоли. Каждая из многочисленных мелких вакуолей по мере притока воды растет, и в конце концов все они сливаются в одну большую центральную вакуоль, ограниченную мембраной. Вся остальная клетка вместе с ростом вакуоли тоже увеличивается; растягиваются клеточные стенки, синтезируются новые порции материала для клеточных стенок и цитоплазмы, различные клеточные органеллы делятся и растут. Таким образом, рост путем растяжения под контролем гормонов и сопутствующее ему увеличение сырого веса происходят в основном именно в той зоне, где клетки вытягиваются в длину.
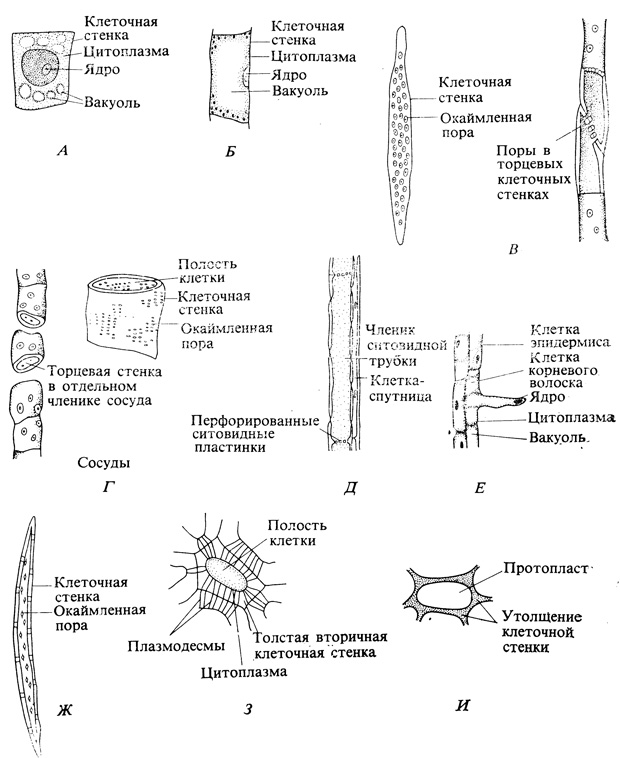
Рис. 3.6. Различные типы растительных клеток (масштаб не соблюден). А. Меристематическая клетка. Б. Паренхимная клетка. В. Две трахеиды; видны поры в их торцевых (соединяющихся) стенках. Г. Несколько члеников сосудов, соединенных конец в конец и образующих трубку. Д. Членик ситовидной трубки и клетки-спутницы. Е. Обычная клетка эпидермиса и клетка корневого волоска. Ж. Волокно. З. Каменистая клетка. Я. Клетка колленхимы
Обычно в одно время с удлинением клеток, а иногда после его завершения происходит и дифференциация. Дифференциацией называют процесс, в результате которого клетки, казавшиеся до того сходными, приобретают морфологические различия и начинают выполнять разные физиологические функции (см. рис. 3.6). Процесс этот - одна из труднейших загадок биологии. Коль скоро каждая клетка многоклеточного организма возникает в конечном счете, как это принято считать, в результате деления одной и той же исходной клетки - зиготы, имеющей двойной набор хромосом, то, очевидно, все эти клетки должны иметь совершенно одинаковые наборы генов. Мы уже знаем, что это не совсем так; из-за нарушений, которые иногда случаются в процессе митоза, число хромосом в разных клетках одного и того же растения может и не совпадать. Однако между наборами генов в тех клетках, которые приобретают в конце концов совершенно разную форму, нет, видимо, все же никаких качественных различий; в культуре можно осуществить регенерацию целого растения из отдельных клеток, откуда бы они ни были взяты - из листа, стебля или корня. Итак, дифференциация, по-видимому, не изменяет основной генетической информации, содержащейся в клетке. Вновь возникшая клетка обладает широкими потенциями и может развиваться (в морфологическом и физиологическом смысле) по любому из нескольких возможных путей, в зависимости от физических и химических воздействий, а также от пространственных соотношений. Пройдя однажды дифференциацию в том или ином направлении, клетка обычно более уже не возвращается в недифференцированное состояние и не переходит в иную форму; однако в условиях культуры тканей может происходить как дедифференциация, так и редифференциация. В настоящее время принято считать, что в любой клетке выражена только какая-то часть общего генетического потенциала, т. е. что одни гены в ней "включены", а другие "выключены". Природа клетки зависит, таким образом, от того, какой комплекс генов оказывается в ней активным. Проблема, следовательно, переходит в иную плоскость и может быть сформулирована так: какой стимул включает и выключает гены?
Знакомясь с физиологией высших растений, мы можем научиться регулировать те или иные процессы дифференциации при помощи химических или физических воздействий. Однако такое умение достигается чисто эмпирическим путем. В сущности, мы похожи при этом на человека, который вставляет ключ в замочную скважину и отпирает дверь, не зная ровным счетом ничего об устройстве самого замка. Совершенно ясно, что будущие успехи экспериментальной биологии должны в значительной мере зависеть от подробного изучения тех явлений, которые происходят в промежутке между воздействием данного специфического агента и появлением соответствующей измененной формы.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'