Порядок миртовые (Myrtales)
Семейство дербенниковые (Lythraceae) (В. Н. Гладкова)
Дербенниковые - травы, кустарники и деревья, широко распространенные во всех климатических поясах Земли, кроме холодного, но особенно часто встречающиеся в тропических и субтропических областях. Около 70% всех видов дербенниковых растут в Америке, особенно в Южной и Центральной, где представлены эндемичные для западного полушария куфея (Cuphea), диплузодон (Diplusodon), лафенсия (Lafoensia), гинория (Ginoria). К тропикам Старого Света приурочены виды лагерстремии (Lagerstroemia), встречающиеся от Азии до Северной Австралии. Всего в семействе насчитывается 25 родов и до 550 видов. Самым крупным по числу видов (200-250) является американский род куфея.
Большинство дербенниковых связано с влажными местообитаниями, особенно это относится к травянистым видам, среди которых много болотных и водных растений. По окраинам болот, берегам рек и озер, на сырых лугах растут виды дербенника (Lythrum), на болотистых местах и в мелкой воде встречаются травы из родов аммания (Ammania), ротала (Rotala), бутерлак (Peplis). Листья растущих в воде видов часто диморфные, а погруженные стебли утолщены вследствие развития концентрических слоев аэренхимы. Другие виды травянистых дербенниковых являются ксерофитами. Среди них характерные для засушливых областей Южной Америки виды плеврофоры (Pleurophora) с колючими листьями, многие узколистные виды куфеи бразильского кампоса и др.
Древесные виды дербенниковых произрастают в тропических лесах, особенно часто по берегам рек, как, например, достигающие в высоту свыше 20 м вечнозеленые деревья из рода лагерстремия, или встречаются на открытых сухих пространствах. На сухой сезон некоторые из них сбрасывают листья и часто начинают цветение в безлистном состоянии.
Листья дербенниковых цельные и цельнокрайные, обычно супротивные, редко очередные (у некоторых видов роталы, бутерлака, дербенника, несеи - Nesaea) или мутовчатые (у видов роталы, куфеи), с очень маленькими шиловидными или волосовидными опадающими прилистниками. Соцветия - кисти, метелки или дихазиальные щитки, клубочки, головки, расположенные в пазухах листьев или терминально. Уникальное "междупазушное" положение имеют цветки куфеи, у которой ножка цветка прирастает к выше расположенному междоузлию и цветок кажется прикрепленным к стеблю ниже узла или на узле между листьями. Цветки от темно-фиолетовой до красной и белой окраски, редко желтые, актиноморфные или зигоморфные (у куфеи, плеврофоры), часто с 2 супротивными прицветничками. У некоторых родов цветки ди- или триморфные в отношении длины тычинок и столбиков. Цветочная трубка от колокольчатой до трубчатой формы, у куфеи при основании со шпорцем или горбом, у видов с невзрачными лепестками бывает окрашена в красный или желтый цвет. Чашелистики травянистые, кожистые или лепестковидные (у роталы, плеврофоры), их 4-6, у многих видов имеется наружная чашечка. Лепестки в числе 0-6, иногда как и чашелистики, между собой неравные, опадающие, прикрепляются на внутренней поверхности трубки. Тычинки в двух кругах, их в 2 раза больше, чем лепестков или один из кругов полностью или частично редуцирован, иногда в результате расщепления число их достигает 100 (у лагерстремии). Они равные или поочередно неравные, прикреплены ниже лепестков, иногда полностью заключены в цветочную трубку, в почкосложении прямые или согнутые в зависимости от их длины, у американского рода лафенсия тычинки достигают в длину 10 см, в почкосложении спирально скручены. Гинецей синкарпный, 2-6-гнездный (редко 1-гнездный) с полностью развитыми или неполными перегородками. Завязь у основания окружена диском, иногда имеющим форму чешуек (у роталы) и отделяющим нектар. У зивоморфных цветков диск развит только с верхней (адаксильной) стороны. Плод - сухая коробочка, заключенная в сохраняющуюся цветочную трубку. Семена в числе от 3 до многих, большей частью мелкие, но иногда достигают в длину 4-5 мм, часто с небольшим крылом.
Способы опыления у дербенниковых очень разнообразны. У аммании и у безлепестных видов роталы и несеи встречаются клейстогамные цветки. У аммании широколистной (Ammania latifolia) пыльцевые зерна прорастают уже в бутоне. Постоянная автогамия характерна и для видов бутерлака - лепестки и нектарники у его цветков отсутствуют, рыльца короткие и над ними нависают интрорзно растрескивающиеся пыльники.
Дербенниковые с зигоморфными цветками приспособлены к опылению птицами и длиннохоботковыми насекомыми. У видов куфеи, морфологически наиболее специализированных в семействе, цветочная трубка длинная и к нектару, отделяемому стенками шпорца у основания трубки ведет длинный и узкий проход. Этот проход создается благодаря тому, что дорсальные тычинки над вместилищем нектара не развиваются, две ближайшие к ним боковые тычинки укорочены и отклонены внутрь, ограничивая путь к нектару с боков, а ниже их располагаются 2 ряда волосков и дорсальный диск в основании завязи. Липкие желёзки на цветочной трубке отпугивают ползающих насекомых-вредителей.
Особым приспособлением к перекрестному опылению является гетероморфность цветков, наблюдающаяся у многих видов дербенниковых. Диморфные цветки известны у видов дербенника, роталы, пемфиса (Pemphis), несеи, а некоторые виды дербенника, несеи и декодона (Decodon) имеют триморфные цветки. Как и у всех видов с диморфными цветками, пыльники более длинных тычинок вырабатывают более крупную пыльцу, которая лучше всего прорастает и дает наиболее хорошие результаты на снабженных более крупными сосочками рыльцах длинных столбиков другой формы и, наоборот, мелкая пыльца коротких тычинок длинностолбчатых форм дает лучшие результаты на рыльцах короткостолбчатых форм. Диморфизм широко распространен у растений, триморфные же формы в пределах одного вида представляют довольно редкое явление в природе, так как для его возникновения необходимо наличие не менее чем двух кругов тычинок. Кроме дербенниковых, он отмечен только в семействах кисличных (Oxalidaceae) и понтедериевых (Pontederiaceae).
Явление гетероморфизма было подробно изучено Ч. Дарвином. С тонкой наблюдательностью в специальной работе он описал половые взаимоотношения триморфных форм дербенника иволистного (Lythrum salicaria, табл. 30). Три формы этого дербенника равно представлены в каждой популяции. Длинностолбчатая форма имеет средней длины тычинки эписепального круга и короткие эпипетального круга. Среднестолбчатая форма имеет длинные, превышающие столбик тычинки эписепального круга и короткие эпипетальные. Короткостолбчатая форма характеризуется длинными эписепальными тычинками и средней длины эпипетальными. Таким образом в цветке каждой формы два круга тычинок и столбик различаются между собой по длине, а в пределах разных форм размер длинных, средних и коротких тычинок совпадает с размером длинных, средних и коротких столбиков (рис. 104). Тычинки и столбики слегка наклонены в ту сторону, где расположен путь к нектару. Насекомые (главным образом пчелиные и двукрылые) садятся на выступающие тычинки или столбик и просовывают хоботки вдоль верхнего края цветочной трубки, чтобы добыть нектар. При этом пыльники длинных тычинок пачкают пыльцой брюшко и внутреннюю сторону задних ножек насекомого, средние тычинки оставляют пыльцу на нижней стороне груди и между передними ножками, а короткие тычинки трутся о хоботок и нижнюю губу. При посещении других цветков рыльца соответствующей длины касаются тех же частей тела насекомого, и перенос пыльцы осуществляется главным образом в пределах равнодлинных элементов, что способствует достижению наибольшей плодовитости при скрещивании. Такие скрещивания между женскими элементами определенной длины с двумя группами тычинок той же длины Дарвин обозначал как легитимные (законные), а 4 других возможных, но более или менее стерильных скрещивания женских элементов с четырьмя группами тычинок иной длины - как иллегитимные (незаконные).

Таблица 30. Непентовые, дербенниковые и гранатовые: 2 - дербенник иволистный (Lythrum salicaria), Московская область
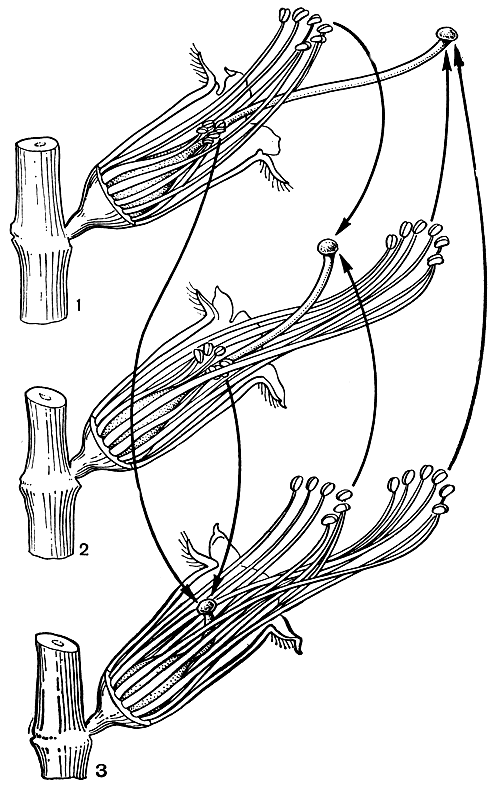
Рис. 104. Три формы цветков дербенника иволистного (Lythrum salicaria): 1 - длинностолбчатая форма; 2 - среднестолбчатая форма; 3 - короткостолбчатая форма. Лепестки и половина чашечки у всех форм удалены. Линии со стрелками показывают, какой пыльцой должны быть опылены рыльца, чтобы вызвать полное оплодотворение (по Ч. Дарвину, 1865)
Триморфизм по сравнению с диморфизмом дает больший процент плодовитых скрещиваний и, по наблюдению Дарвина, имеет, возможно, большое значение в жизни видов, ограниченных в своем распространении строгой экологической приуроченностью к определенным местообитаниям.
Коробочки дербенниковых с пленчатыми или кожистыми стенками, не вскрывающиеся или чаще вскрывающиеся по гнездам, по перегородкам, зубчиками на верхушке или неправильно разламывающиеся. Оригинальный способ вскрывания характерен для куфеи: локулицидная коробочка расщепляется по длине адаксиальной стенки, одновременно расщепляется с верхней стороны цветочная трубка, плацента резко отгибается назад, выступая из плода и выставляя на воздух еще незрелые семена, которые, дозревая, постепенно осыпаются (рис. 105). Крыло у семян дербенниковых служит как для распространения их ветром, так и как приспособление к плавучести, так как оно состоит из легкой губчатой ткани. Благодаря плавучести семян широко распространился по побережьям и коралловым рифам тропиков Старого Света кустарник (или небольшое деревце) пемфис кисловатый (Pemphis acidula). Семена его плавают как самостоятельно, так и используют доступный "транспорт" - их часто находят в кусках пемзы, вымываемой с вулканических островов, а морские птицы разносят семена пемфиса вместе с его ветками, которые они используют для строительства гнезд. Хорошей плавучестью отличаются легкие крылатые семена обильных по берегам рек видов лагерстремий: вода - дополнительный фактор в их распространении. Можно наблюдать, как постепенно завоевывает прибрежное пространство, продвигаясь по течению реки, дербенник иволистный. Вода разносит, однако, не его семена, которые быстро тонут, а проростки. Семена дербенника прорастают под водой, проростки всплывают на поверхность, плывя по течению, пока их не прибьет где-нибудь к берегу.
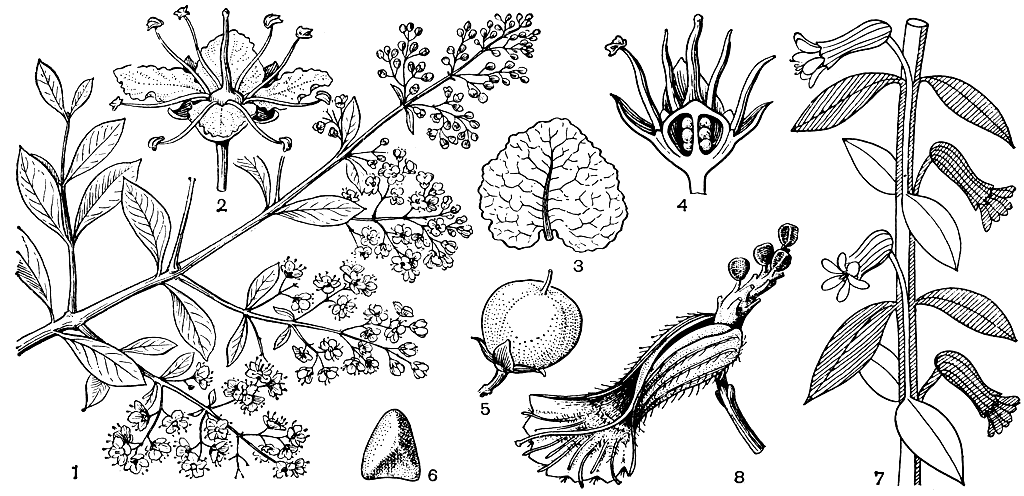
Рис. 105. Дербенниковые. Лосония невооруженная, хна (Lawsonia inermis): 1 - ветвь с цветками; 2 - цветок; 3 - лепесток; 4 - продольный разрез завязи; 5 - плод; 6 - семя. Куфея неприятная (Cuphea ingrata): 7 - часть стебля с внепазушными цветками; 8 - коробочка после растрескивания с отогнутой плацентой
Семена некоторых водных видов дербенниковых (куфеи, бутерлака, роталы, несеи) обнаруживают замечательную особенность: при попадании в воду их первоначально гладкая поверхность довольно быстро становится грубоволосистой. Волосовидные образования, имеющиеся в наружном слое эпидермы, у сухих семян обращенные внутрь клеток и свернутые, при увлажнении разворачиваются, пробивая эпидерму и выходя наружу, при этом они выделяют слизь, которая способствует прикреплению семян к проплывающим предметам.
Некоторые дербенниковые, как виды аммании и роталы, расширяют свой ареал в качестве сорняков рисовых посевов.
Для ряда видов характерно вегетативное размножение частями побегов (бутерлак) или путем укоренения дугообразных ветвей (у североамериканского монотипного рода декодон).
Некоторые дербенниковые известны как красильные растения, из них наибольшее значение имеет лосония невооруженная, или хна, хенна (Lawsonia inermis, см. рис. 105), растущая в тропиках Старого Света от Северной Австралии, Малайского архипелага и Индии до Аравии, Северной и Восточной Африки и Мадагаскара и культивируемая в Азии, Африке и тропической Америке. Из сухих листьев этого растения получают оранжево-красную краску, которая издавна используется главным образом на Востоке для окраски волос и ногтей. Считают, что культура этого растения возникла в Иране и близлежащих странах. Кристаллы красящих веществ из листьев лосонии применяют для окраски шерсти, шелка и пищевых продуктов; эфирное масло из ее пахнущих чайной розой цветков используют в парфюмерии. В СССР лосонию выращивают как двулетнюю культуру в Средней Азии и на Кавказе.
Древесные виды лагерстремии, лафенсии, физокалиммы (Physocallymma) дают полезную древесину. Отдельные виды локально используют как лекарственные растения и выращивают как декоративные. Особенно популярна в культуре в теплых странах лагерстремия индийская (Lagerstroemia indica), во время цветения покрывающаяся красивыми крупными метелками пурпурных, розовых или белых цветков. В СССР ее выращивают в садах и парках Черноморского побережья.
Листья бутерлака портулакового (Peplis portula) и пемфиса кисловатого (Pemphis acidula) употребляют на салат.
Семейство соннератиевые (Sonneratiaceae) (С. С. Морщихина)
Семейство соннератиевых включает всего 7 видов, относящихся к двум родам - соннератия (Sonneratia, рис. 106) и дуабанга (Duabanga), распространенных в тропиках восточного полушария.
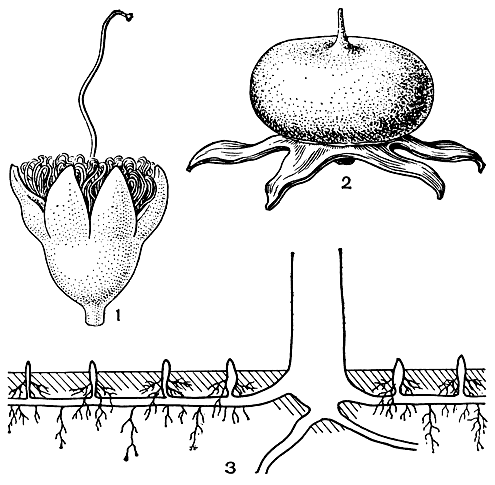
Рис. 106. Соннератия Гриффита (Sonneratia griffithii): 1 - цветок; 2 - плод с чашелистиками; 3 - схема корневой системы соннератии (по В. Троллю)
Виды соннератии произрастают в мангрове на морских побережьях, два вида дуабанги - во влажных тропических лесах. Соннератиевые - вечнозеленые деревья высотой 15-20 м, иногда больше. В знаменитом ботаническом саду Перадения на острове Шри-Ланка известны роскошные столетние экземпляры дуабанги молуккской (D. moluccana) высотой 35-40 м, ветвящиеся на высоте 2,5-3 м, с диаметром кроны около 30 м. Молодые веточки соннератиевых гибкие, повисающие. Листья супротивные, простые, цельные, с рудиментарными опадающими прилистниками. Во время сильных дождей соннератии поворачивают пластинки листа вертикально.
Соннератии, растущие на затопляемом приливами грунте, образуют на длинных горизонтальных, кабельных корнях многочисленные вертикальные, торчащие над поверхностью земли выросты, особенно мощные у соннератии сырной (S. caseolaris). Эти выросты способны к вторичному утолщению, покрыты рыхлой корой с обильными межклетниками и несут много мелких питающих корней, образующихся в верхней части заново по мере накопления ила и песка. Главное их значение в жизни дерева - вынос питающих корней в верхние плодородные слои почвы.
Цветки соннератиевых довольно крупные, актиноморфные, обоеполые, расположенные по 1-3 или в небольших щитках ветвей. Чашечка колокольчатая из 4-8 долей, сросшихся в короткую трубку, иногда ярко окрашенных в красный цвет с внутренней стороны, кожистая, неопадающая. Лепестки в том же числе, у некоторых видов соннератии отсутствуют или очень узкие, линейные, малозаметные; у дуабанги лепестки довольно крупные, гофрированные, белые или желтоватые, быстро опадающие. Тычинки большей частью многочисленные, расположенные в несколько рядов, у дуабанги молуккской их всего 12 в одном круге. Тычиночные нити белые, в нижней части расширенные, обычно длинные и свернутые в бутоне. Пыльники почковидно изогнутые, двугнездные, открываются продольно. Гинецей из 4-20 плодолистиков, завязь, сросшаяся с широким цветоложем, многогнездная, с многочисленными семязачатками на центральной плаценте. Столбик длинный, скрученный в бутоне, рыльце головчатое, крупное, у соннератии безлепестковой (S. apetala) диаметром до 7 мм, липкое, часто облепленное кварцевыми песчинками, принесенными ветром.
Цветение у соннератиевых ночное, кратковременное, на рассвете лепестки и тычинки опадают. Цветки издают неприятный запах, выделяют обильный нектар и посещаются вечером и на рассвете птицами-нектарницами и ночью многочисленными летучими мышами (Ван дер Пэйл, 1956).
Плоды у дуабанги - коробочки, раскрывающиеся по створкам, у соннератии же плоды - ягоды, сжато-шаровидные, с неопадающим столбиком и чашелистиками. Семязачатки у соннератиевых коленчато изогнуты и семена имеют характерно изогнутую форму, заполнены зародышем и лишены эндосперма. Семена дуабанги на обоих концах имеют узкие выросты семенной оболочки, способствующие распространению их ветром на большие расстояния. Проростки дуабанги очень светолюбивы, деревья встречаются в лесах рассеянно. В отличие от дуабанги виды соннератии образуют чистые стояния, но преимущественно открытые, несомкнутые или растут вместе с ризофоровыми. Плоды соннератии быстро сгнивают на земле, семена с легкой оболочкой разносятся водой в прилив и прорастают без периода покоя, оседая на пляжах и коралловых террасах.
Виды соннератии распространены от берегов Восточной Африки и Мадагаскара по побережьям тропической Азии до островов Хайнань и Рюкю (Нансей), Микронезии, на Больших и Малых Зондских островах, в Северной Австралии, на Новой Гвинее, Соломоновых островах и Новой Каледонии. Из пяти видов соннератии наиболее широкое распространение имеет соннератия белая (S. alba) и соннератия сырная.
В роде дуабанга вид дуабанга крупноцветковая (D. grandiflora) распространен в Юго-Восточной Азии, от южных склонов Восточных Гималаев до полуострова Индокитай и Малакка, а дуабанга молуккская встречается только на островах Малайского архипелага, на Филиппинских, Молуккских островах и Новой Гвинее.
Древесина соннератиевых находит применение в строительстве домов и лодок, однако большого экономического значения не имеет.
Семейство гранатовые (Punicaceae) (В. И. Трифонова)
С дербенниковыми очень тесно связано семейство гранатовых, представленное всего 1 родом с двумя видами, из которых один - гранат сокотранский (Punica protopunica) - эндемичен для острова Сокотра, а второй - гранат обыкновенный (P. granatum, рис. 107) - произрастает дико на Балканском полуострове и в Западной Азии, доходя на востоке до границ Северо-Западной Индии.
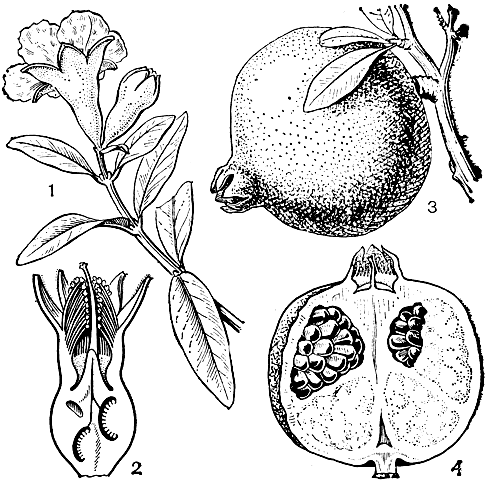
Рис. 107. Гранат обыкновенный (Punica granatum): 1 - ветвь с цветком и бутоном; 2 - разрез длинностолбчатого цветка; 3 - плод; 4 - разрез плода
Гранатовые - невысокие вечнозеленые деревья или листопадные кустарники с побегами, часто заканчивающимися колючками. Листья цельные, обычно супротивные, скученные на коротких стеблях, с рудиментарными прилистниками. Цветки обоеполые, актиноморфные, располагаются по одиночке или собраны в пучки на концах пазушных побегов. Окрашенная, толстая, 4-7-лопастная чашечка остается при плодах. Лепестков 4-7, ярко-пунцовых, реже белых или желтоватых, скомканных в бутоне наподобие лепестков мака. Тычинки многочисленные, расположенные в 3-4 круга, как и лепестки, прикрепленные к краю цветочной трубки. Гинецей из нескольких (до 7) плодолистиков, заканчивается тонким столбиком с головчатым рыльцем; завязь нижняя, многогнездная, с одним (гранат сокотранский) или двумя кругами гнезд. У граната обыкновенного благодаря разрастанию цветоложа происходит смещение наружного круга плодолистиков вверх, в результате чего образуются 2 яруса гнезд. Очень редко можно наблюдать 3-4 яруса. Такое своеобразное строение завязи-явление крайне редкое среди покрытосеменных и встречается только у граната обыкновенного. Плод - гранатина размером с крупное яблоко (табл. 30) - с кожистым пурпурно-красным или зеленоватым околоплодником-наполнен многочисленными семенами со стекловидной розово-красной пульпой (сочным наружным слоем семенной кожуры). Семена без эндосперма, с большими свернутыми одна на другую семядолями.

Таблица 30. Непентовые, дербенниковые и гранатовые: 3 - плоды граната обыкновенного (Punica granatum), Никитский ботанический сад
Дикорастущий гранат обыкновенный растет на сухих щебнистых и каменистых склонах, поднимаясь до высоты 1000 м над уровнем моря.
Нередко его можно встретить в подлеске алеппской сосны и литоральных дубов. Он образует чистые заросли или растет вместе с мушмулой, ежевикой и держи-деревом. Гранат сокотранский - невысокое (2-4,5 м) вечнозеленое дерево, довольно редко встречающееся в природе, растет на известняковых и каменистых плато на высоте 250-610 м над уровнем моря, нередко среди зарослей кротона (Croton). От граната обыкновенного он отличается более мелкими цветками, приподнятыми на цветоножках, строением завязи, плода и рядом других анатомических и морфологических признаков. Однако эти различия все же недостаточны для выделения граната сокотранского в самостоятельный род.
Гранатовые - перекрестноопыляемые растения. Днем, когда цветки их распускаются наиболее интенсивно, над растениями кружатся рои пчел. Более редкие гости - жуки, бабочки и муравьи. Наряду с перекрестным опылением может происходить и самоопыление, чему способствует особое положение молодого рыльца, загнутого таким образом, что оно соприкасается с нижним рядом созревших пыльников. Цветок остается раскрытым 3-4 дня, на второй день начинают созревать и лопаться пыльники. Несмотря на недолговечность отдельных цветков, цветение продолжается долго: растение стоит покрытое цветками от 1 до 3 месяцев. После опыления цветки с короткими столбиками, функционирующие как мужские, опадают так же, как и цветки промежуточного типа, а из длинностолбчатых цветков развиваются плоды. Как было подсчитано, процент таких цветков невелик и колеблется от 5 до 20% от общего числа цветков на растении и зависит от сорта и возраста растений.

Таблица 30. Непентовые, дербенниковые и гранатовые: 4 - цветок граната протопуники (Punica protopunica), Сокотра
Гранат обыкновенный обладает многими полезными свойствами. Это одна из древнейших культур Средиземноморья, о чем говорят его находки в египетских захоронениях и изображения на древневизантийских тканях. Плоды его нередко служили образцом для орнаментов у древних греков, арабов, египтян и даже ассирийцев. В настоящее время его культивируют во многих странах мира. В нашей стране гранат разводят в Закавказье, в Крыму и Средней Азии. Сочные семена граната очень вкусные. Сок, получаемый из его семян, содержащий до 14% витамина С, является прекрасным освежающим напитком, из него готовят мягкие сухие вина и шербеты, напиток гренадин и пунши. Кора ветвей и околоплодника богата дубильными веществами; ее используют при дублении и окраске в черный цвет особо тонких сортов кожи типа сафьяна. Настой из цветков, коры ветвей и особенно корней обладает лекарственными свойствами. Листья служат заменой чая. Горьковато-деревянистые на вкус европейца плоды граната сокотранского считаются довольно приятными у местных жителей.
Гранат обыкновенный - прекрасное декоративное растение; особенно хорошо оно во время цветения, сплошь покрытое массой довольно крупных пунцово-красных цветков. Его используют для живых изгородей и одиночных посадок.

Таблица 30. Непентовые, дербенниковые и гранатовые: 5 - плоды граната протопуники
Семейство меластомовые (Melastomataceae) (Н. Н. Имханицкая)
В семействе меластомовых до 240 родов и не менее 4500 видов (а по мнению некоторых авторов, даже более 5000), свойственных тропическим и частично субтропическим областям обоих полушарий. Лишь немногие его представители, как род рексия (Rhexia), заходят в умеренную зону. Более половины всех родов и две трети видов меластомовых сосредоточены в тропиках Нового Света. Особенно богата ими Южная Америка. Поразительное разнообразие меластомовых, встречающихся в самых различных местообитаниях - от морских побережий до 4000 м над уровнем моря в Андах,- составляет одну из особенностей растительного мира американских тропиков. Меластомовые широка распространены также в тропической Азии, на островах Океании, в Африке и на Мадагаскаре. В Австралии известно всего 5 родов (с 9 видами).

Рис. 108. Меластомовые. Мединилла великолепная (Medinilla magnifica): 1 - ветвь с цветками; 2 - цветок; 3 - тычинка (виден придаток связника); 4 - плод. Тибухина пятитычинковая (Tibouchina semidecandra): 5 - цветок; в - диморфные тычинки. Рексия вирджинская (Rhexia virginica): 7 - цветок; 8 - тычинка; 9 - плод
Меластомовые - многолетние или однолетние травы, кустарнички, кустарники, реже деревья, иногда листопадные, лианы и эпифиты. Среди них есть болотные и водные растения, галофиты и ксерофиты. Стебли меластомовых часто 4-угольные в поперечном сечении; листья супротивные, обычно крест-накрест расположенные, редко мутовчатые. Они часто крупные, кожистые или мясистые, разнообразные по форме, цельные, пильчатые или зубчатые по краю, нередко покрыты, как и другие части растения, чешуями, разнообразными волосками, щетинками или сосочками, лишены прилистников. Часто листья очень красиво окрашены, нередко пестрые, с пятнами или точками. Они снабжены 3-9(11) рельефно выступающими продольными жилками, дуговидно восходящими от основания листа к верхушке; они соединяются друг с другом и с главной жилкой многочисленными поперечными, часто почти параллельными жилочками, редко листья перистонервные.
Цветки меластомовых отличаются яркими пурпурными, фиолетовыми, розовыми, реже ярко-красными, белыми или желтыми лепестками, розовыми, голубыми, белыми тычиночными нитями и крупными желтыми, розовыми, пурпурными или фиолетовыми пыльниками (табл. 31). Обычно цветки крупные (у некоторых бразильских видов рода тибухина (Tibouchina, рис. 108), диаметром до 20 см, реже мелкие или даже совсем крошечные. Они собраны в разнообразные верхушечные или пазушные верхоцветные соцветия или одиночные, нередко с окрашенными прицветниками. Цветки обоеполые, редко однополые, обычно 4-5(6)-членные, актиноморфные или слегка зигоморфные из-за расположения тычинок. Цветочная трубка свободная или частично или полностью приросшая к завязи, иногда соединенная с ней лишь продольными перегородками, часто ярко окрашенная. Лопасти чашечки опадающие или остающиеся, створчатые, иногда они редуцированы до простого ободка или срастаются в колпачок (калиптру), отваливающийся при раскрывании цветка. Лепестки обычно свободные, в бутоне скрученные вправо. Хотя число тычинок сильно варьирует у различных представителей семейства, обычно их вдвое больше, чем лепестков, и расположены они в два круга (рис. 109). Тычинки наружного и внутреннего кругов равные или чаще различаются величиной, окраской пыльников и придатками связника. В бутоне тычинки согнуты вниз, и пыльник, обращенный своей верхушкой к основанию цветка, заполняет все пространство между завязью и цветочной трубкой. При раскрывании цветка тычиночная нить выпрямляется и выносит пыльник вверх. В раскрытом цветке тычинки располагаются симметрично или нередко они наклоняются к одной стороне цветка. Пыльники вскрываются обычно единственной верхушечной порой, редко 2 порами или 2 продольными щелями. Связник часто продлен ниже пыльцевых гнезд и обычно снабжен в месте соединения с тычиночной нитью разнообразными по величине и форме придатками. У более примитивных родов тычинки не имеют развитых придатков. У растений с диморфными тычинками пыльники крупных тычинок (обычно из наружного круга) по своей окраске почти неотличимы от лепестков, тогда как пыльники коротких тычинок (как правило, внутреннего круга) ярко окрашены в желтый цвет и вместе с желтыми придатками крупных тычинок, расположенными непосредственно под ними, привлекают насекомых. Желтые пыльники коротких тычинок служат в основном лишь для привлечения насекомых, собирающих пыльцу. Крупные же пыльники длинных тычинок, сливающиеся по цвету с лепестками и потому малозаметные для насекомых, предназначены только для опыления.
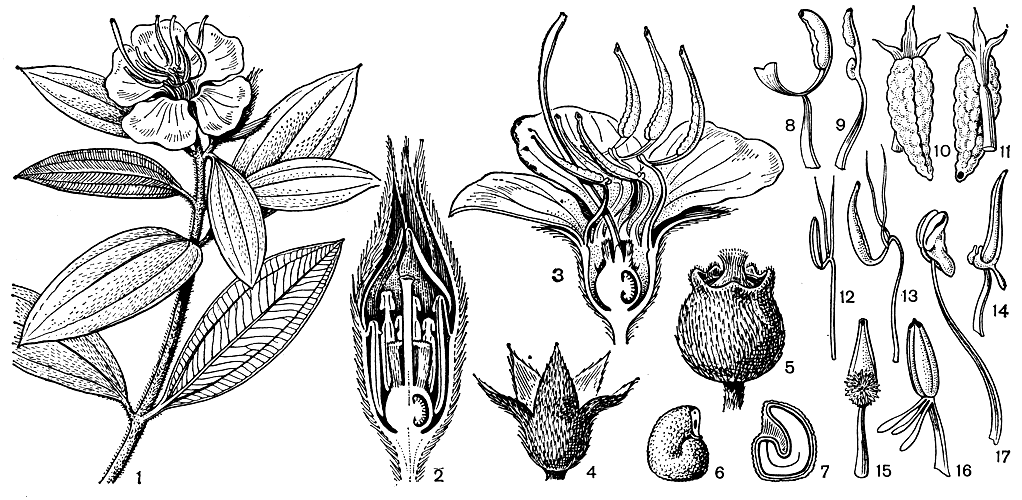
Рис. 109. Меластомовые. Меластома малабарская (Melastoma malabathricum): 1 - ветвь с цветком; 2 - продольный разрез бутона (тычинки согнуты вниз, пыльник заполняет пространство между завязью и цветочной трубкой); 3 - продольный разрез цветка (видны диморфные тычинки); 4 - чашечка; 5 - плод; 6 - семя; 7 - продольный разрез семени. Центрадения неравнобокая (Gentradenia inaequilateralis): 8, 9 - диморфные тычинки. Омфалопус обманчивый (Omphalopus fallax): lo - тычинка (вид спереди); 11 - тычинка (вид сзади). Аппендикулярия тимьянолистная (Арреп-dicularia thymifolia): 12, 13 - диморфные тычинки. Мединилла яванская (Medinilla javanensis): 14 - тычинка. Погонантера отвернутая (Pogonanthera reflexa): 15 - тычинка. Триолена скорпионовидная (Triolena scorpioides): 16 - тычинка. Мурири пуса (Mouriri pusa): 17 - пыльники вскрываются продольной щелью

Таблица 31. Меластомовые: 1 - амфиблема цимозная (Amphiblemma cymosum), оранжерея Ботанического института АН СССР в Ленинграде; 2 - триолена жёстковолосистая (Triolena hirsute), оранжерея Ботанического института АН СССР в Ленинграде
У большинства меластомовых завязь нижняя, реже верхняя, 2-15-гнездная, иногда 1-гнездная, как у некоторых видов рода мурири (Mouriri), обычно с многочисленными семязачатками в каждом гнезде на угловой плаценте, редко на париетальной или свободной центральной, или базальной. Столбик нитевидный, прямой или сильно наклоненный на одну сторону цветка, заканчивается простым рыльцем. Плод - многосемянная или односемянная ягода с сочным или кожистым околоплодником (табл. 31) или многосемянная коробочка, включенная в цветочную трубку. Семена обычно многочисленные, мелкие, прямые или согнутые, улитковидные, клиновидные, булавовидные, неправильно угловатые или шаровидные, иногда крылатые, без эндосперма, с очень маленьким зародышем с мясистыми семядолями, из которых одна может быть крупнее другой.
Меластомовые разделяют на три подсемейства: собственно меластомовые (Melastomatoideae), астрониевые (Astronioideae) и мемецилоновые (Memecyloideae). У видов двух первых подсемейств, различающихся типом плацентации, плоды многосемянные и очень мелкие, семена с маленьким зародышем, тогда как представители третьего подсемейства имеют 1-2-семянные плоды и семена с крупным зародышем. В большинстве родов подсемейства меластомовых пыльники вскрываются верхушечными порами, а у представителей двух других подсемейств - продольными щелями.
Подавляющее большинство родов относится к подсемейству меластомовых. Важнейшие роды этого подсемейства - микония (Miconia), мединилла (Medinilla, см. рис. 108), тибухина, леандра (Leandra), клидемия (Clidemia) и др. Род микония - самый богатый видами род семейства. Он объединяет более 700 (до 900) видов деревьев и кустарников тропической Америки, встречающихся от уровня моря до альпийских парамо.
Подсемейство астрониевых ограничено Малезией и Полинезией. Род астрония (Astronia) насчитывает около 70 видов кустарников или среднего размера деревьев, произрастающих в подлеске низинного и горного влажного леса от уровня моря до 2500 м над уровнем моря. Несколько видов астронии являются обычной составной частью лесов на многих тихоокеанских островах. Центр видового разнообразия астронии - Филиппины. Шаровидная коробочка (диаметром до 1 см) этих растений содержит многочисленные линейные семена, которые могут переноситься ветром.
Пантропическое подсемейство мемецилоновые, которому иногда придают ранг семейства, связывает меластомовые с миртовыми. Ареал его охватывает тропическую Америку и тропики Старого Света. Род мемецилон (Memecylon) включает около 300 видов деревьев (иногда высотой до 25 м) и кустарников, встречающихся в подлеске низинных и горных лесов, по берегам рек, на побережье в тропических областях Африки, Азии и Океании. Род мурири объединяет около 50 видов кустарников и деревьев, которые особенно разнообразны в долине реки Амазонки. Бразильская мурири стволоцветковая (M. trunciflora) образует крупные, длиной до 10 см, ароматные, сочные, грушевидные плоды, приятные на вкус, сверху покрытые слоем клейкого вещества.
Без многочисленных растений из семейства меластомовых трудно представить тропический дождевой лес. Это разнообразные по облику кустарники и невысокие деревца, входящие в состав подлеска, лианы, прикрепляющиеся к стволам деревьев придаточными корнями, эпифиты, обильно покрывающие крупные деревья, или полуэпифитные растения, еще не утратившие окончательно связи с почвой. На влажных, затененных местах под пологом леса, вблизи воды, на покрытых мхом влажных скалах можно встретить меластомовые - травы с нежными или сочными листьями, восходящие, вьющиеся или распростертые, ползучие и укореняющиеся или даже бесстебельные, как, например, гравесия удивительная (Gravesia mirabilis) во влажных лесах Мадагаскара с розеткой красиво окрашенных листьев. Число меластомовых, ведущих эпифитный образ жизни, довольно велико. Они поселяются на стволах и крупных ветвях высоких деревьев, на мертвых растениях или на скалах, используя их лишь в качестве опоры. Мощная корневая система, прочно прикрепляющая растение к опоре, нередко снабжена разнообразной формы и величины клубнями. Продолговатые, мясистые, сладковатые на вкус клубни мединиллы серповидной (M. falcata) скучены в крупную массу, вес которой может достигать нескольких килограммов.
Многие представители меластомовых встречаются в заболоченных лесах, на болотах. У растений, приспособленных к жизни в воде, в погруженных в воду частях стебля и корнях развита воздухоносная ткань.
Меластомовых можно встретить также на морских побережьях, на коралловых рифах, в мангровых болотах. Немало растений этого семейства встречается в засушливых условиях - на сухих и солнечных местах, на солончаках, в саваннах, на скалах. Некоторые виды растут в кратерах вулканов, на старых потоках лавы, вблизи горячих источников. На сухих плоскогорьях внутренних районов Бразилии в кампосе обильны невысокие сильно опушенные жестколистные кустарники или кустарнички, имеющие нередко эрикоидный облик, или травы с мелкими чешуевидными, часто черепитчато расположенными на стеблях листьями. Из растений африканских саванн особенно замечательны виды рода диссотис (Dissotis), насчитывающего около 140 видов тропической и Южной Африки. Это главным образом травянистые растения, а также кустарнички или кустарники, обычно волосистые, с пурпурными или фиолетовыми цветками. Одни виды диссотиса - эфемеры, небольшие однолетние растения, которые успевают пройти весь цикл развития во влажное время года до наступления сухого сезона. Другие виды являются эфемероидами. Некоторые виды диссотиса - клубнеобразующие многолетние травы. Во время сухого периода надземные органы эфемероидов полностью отмирают, живыми остаются лишь снабженные почками клубни или корневища.
В лесах тропической Южной Америки, особенно в долине реки Амазонки, известно немало меластомовых, на листьях которых имеются крупные мешковидные вздутия. Они выдаются на верхней поверхности листа в виде двойных, разделенных продольными перегородками пузырей, открываясь наружу небольшими отверстиями в пазухах базальных жилок на нижней стороне листа. В этих полых вздутых мешках поселяются маленькие, больно жалящие муравьи, которые, очевидно, охраняют растение от крупных муравьев-листорезов, приносящих ему большой вред. Отсюда название этих вздутий - формикарии (от латинского названия рода муравьев - Formica). Туземцы всегда избегают такие растения. Стоит лишь чуть-чуть потрясти листья, как тотчас муравьи выбираются из своих жилищ и нападают на человека. Формикарии разнообразны по форме, от узкоовальных до шаровидных, иногда довольно крупные. Чаще всего возникают они в основании листовой пластинки или на черешке, реже на стебле непосредственно под черешком. У многих растений с такими мешками листья анизофильные. На ветвях тококи гвианской (Tococa guianensis) нередко можно видеть висячие "муравьиные сады". Затопление почвы высокой водой делает невозможным строительство муравейников на земле, вынуждая муравьев сооружать свои жилища на низких деревьях или кустарниках, куда они сносят вместе с землей семена и плоды различных растений. В муравейниках развиваются своеобразные сообщества эпифитных растений, так называемых "муравьиных эпифитов", многие из которых приурочены к "муравьиным садам". Плоды тококи, приятные на вкус, едят не только местные жители, но и обезьяны. Вот почему это растение называют также "деревом макак".
В Перуанских Андах встречается птерокладон Спруса (Pterocladon sprucei) из монотипного рода со вздутыми полыми конечными ветвями с зонтиковидными соцветиями, которые постоянно населены муравьями. К числу мирмекофильных растений принадлежат и эпифитные виды малайского рода пахицентрия (Pachycentria); муравьи селятся в корневых клубнях этих растений.
Цветки меластомовых опыляют главным образом собирающие пыльцу крупные пчелы и шмели, а также осы, маленькие мухи, муравьи и жуки. Цветки некоторых меластомовых опыляют колибри и летучие мыши. Кроме яркой окраски лепестков и тычинок, запаха цветков и нектара, насекомых привлекают также окрашенные прицветники либо красные цветоножки и оси соцветия. Своеобразные, вздутые в основании пыльники ряда меластомовых по своему действию при выбрасывании пыльцы напоминают воздуходувные мехи. Впервые такие пыльники были описаны у рексии вирджинской, или луговой красавицы, или оленьей травы (Rhexia virginica, см. рис. 108). Это травянистое растение с клубневидно утолщенными корнями и с крупными красивыми пурпурными 4-членными цветками украшает влажные песчаные луга и болота от штата Нью-Йорк до Каролины и Флориды. Самая нижняя часть ярко-желтого удлиненного и изогнутого одногнездного пыльника имеет форму пузыревидного мешка, в основании которого - шпорцевидный придаток связника. Столбик расположен значительно ниже и позади пыльников. К его липкому рыльцу тотчас пристает пыльца от брюшка пчелы. Набухший мешок в основании пыльника при нажимании на него лапками пчелы действует как воздуходувные мехи, вызывая струю пыльцы, с силой выбрасываемую из единственной мельчайшей верхушечной поры на тело насекомого, в сторону, противоположную от рыльца. При этом пыльник сначала подпрыгивает вперед, а затем возвращается в исходное положение. Аналогичным образом выбрасывается пыльца у некоторых других меластомовых. При опылении гетероцентрона (Heterocentron) и меластомы (Melastoma) существенную роль играют придатки связника, которые под тяжестью насекомого или от толчка в момент его отлета приводят в движение крупные пыльники, вызывая выбрасывание пыльцы из верхушечных пор на тело насекомого. Механизм опыления этих растений, очевидно, однотипен. Цветки меластомы опыляются крупными пчелами и шмелями, собирающими пыльцу. Привлекаемая желтыми пыльниками, пчела приземляется на сильные нити тычинок, при этом надавливая на вильчатые придатки связника крупных тычинок, находящиеся непосредственно под желтыми пыльниками. Крупные фиолетовые пыльники, по окраске не отличающиеся от лепестков, тотчас опускаются вниз, в сторону от тела насекомого. По мере того как пчела "доит" "пищевые" пыльники сильными движениями своих челюстей, "пищевая" пыльца набрасывается на ее брюшко. Пчела издает жужжащий звук, вибрируя своими крыльями, и вызывает целое облако пыльцы из фиолетовых "опыляющих" пыльников, которые действуют как катапульты. Связник способствует катапультированию пыльцы. Пыльца выбрасывается на спинку улетающей пчелы не только в результате вибрации, но и при поднимании пыльников в момент отлета пчелы. После нескольких посещений спинка пчелы полностью припудрена пыльцой. Рыльце касается спинки насекомого, не наблюдается контакта брюшка с "опыляющими" пыльниками и рыльцем. Два вида пыльцы иногда переносятся пчелой раздельно. Крупные размеры меластомовых делают невозможным их опыление маленькими пчелами, которые совершают набег на "пищевые" пыльники, но не осуществляют опыление.
Наиболее полно опыление было изучено у ряда родов трибы тибухиновых (Tibouchineae) - тибухины, итатиайи (Itatiaia) и брахиотума (Brachyotum). Неотропический род тибухина объединяет свыше 200 видов деревьев, кустарников, которые особенно обильны в Бразилии и в Андах. Цветки у большинства видов крупные, иногда окружены прицветниками, часто срастающимися и закрывающими бутон. Тычинки с короткими и длинными изогнутыми пыльниками, с сильно продолженными связниками с 2 маленькими бугорками, или шпорцами придают цветку вид паука (отсюда и название рода - паук-цветок). Цветки большинства видов тибухин опыляют крупные пчелы и шмели, собирающие пыльцу. В цветках некоторых тибухин с выраженной протандрией и с обильно выделяющимся нектаром были обнаружены муравьи. Маленькие и крошечные осы и даже пчелы нередко прогрызают сморщенную верхушку пыльника в цветках тибухин и таким образом собирают пыльцу. Другая группа насекомых (крупные осы) съедает частично или иногда уничтожает полностью тычинки. Защитой от непрошеных гостей, особенно муравьев, у тибухин служат железистые волоски на связнике и тычиночных нитях. Один из видов этого рода - тибухина толстая (T. grossa) - опыляется летучими мышами, питающимися обильно выделяющимся нектаром цветков. Это растение встречается в горах Колумбии, Эквадора и Перу. Темно-красные цветки издают сильный, особенно ночью, капустный запах. Плотные мясистые лепестки длиной 2,5-3 см с сальной верхней поверхностью образуют колокольчатый венчик. Опыление цветков осуществляется летучими мышами.
Маленький кустарник брахиотум багульниколистный (Brachyotum ledifolium), встречающийся в Андах, Эквадоре и Перу, замечателен тем, что его цветки опыляются колибри. Вздутое основание пыльников несет каплю нектара, который выделяется на верхушке тычиночных нитей. Когда колибри всовывает клюв внутрь цветка, то клюв ударяет по набухшему основанию пыльников, и тотчас из крошечных верхушечных пор выбрызгивается струя пыльцы и попадает на мелкие перья в основании клюва. При посещении других цветков перышки головы колибри с прилипшей пыльцой касаются выступающего рыльца. При давлении на эластичную стенку пыльника спичкой струя пыльцы может быть выброшена на расстояние 3 см. Очевидно, другие виды этого рода также являются орнитофильными. Цветки же брахиотума Бентама (B. benthamianum), эндемичного для Эквадора и Перу, приспособлены к опылению пчелами и шмелями. У некоторых меластомовых наблюдается каулифлория. У ряда видов отмечено самоопыление.
Сочные яркие ягоды меластомовых привлекают плодоядных птиц и других животных (обезьян, белок), которые, охотно поедая плоды, распространяют семена. Плодами меластомовых питаются бульбули, голуби, вороны, индийская птица мина, бразильская сабиа и другие плодоядные птицы.
Мелкие семена из сухих открытых коробочек у различных представителей семейства могут переноситься ветром. К анемохорным растениям, очевидно, принадлежат виды меластомовых с крылатыми семенами.
Хозяйственное значение семейства невелико. Меластомовые известны главным образом как декоративные растения. В листьях, плодах и коре многих видов содержатся красящие вещества. Получаемые из них краски местное население использует для окрашивания тканей, циновок или в качестве чернил (например, из ягод миконии). Из коры некоторых тибухин получают черную краску. Желтое красящее вещество, в обилии содержащееся в листьях ряда видов мемецилона, заменяет на Шри-Ланке шафран, и потому этот род называют "шафрановым деревом". Сочные плоды многих меластомовых приятны на вкус и употребляются в пищу или для приготовления освежающих напитков, мармелада, варенья. Виды американского рода беллуция (Bellucia) - плодовые растения. Плоды мурири - продукт питания местного населения в Бразилии. Листья, кора, ягоды многих меластомовых имеют лечебные свойства и применяются в народной медицине. Листья меластомы малабарской (M. malabathricum) - источник питания шелкопряда, производящего высококачественный шелк. Плоды этого растения съедобны, из них получают черную или пурпурную краску, из которой приготавливают чернила. Древесина ряда видов находит применение в строительстве домов, для изготовления мебели, инструментов, для получения древесного угля хорошего качества. Твердая и тяжелая ("железная") древесина мемецилона съедобного (Memecylon edule) используется для стропил и свай при строительстве домов, известна в Индии как заменитель самшита. Плоды этого растения съедобны.
Семейство миртовые (Myrtaceae) (Е. Н. Немирович-Данченко)
Миртовые - одно из самых крупных семейств в порядке миртовых. Оно включает около 140 родов и, вероятно, не менее 3000 видов, обитающих главным образом в тропических странах и особенно многочисленных в Австралии и тропической Америке. Из всех представителей семейства дальше всех на север продвинут мирт обыкновенный (Myrtus communis, рис. 110), достигающий Азорских островов, Европы и Западной Азии, а в южном полушарии - метросидерос зонтичный (Metrosideros umbellatus), достигающий острова Кэмпбелл.
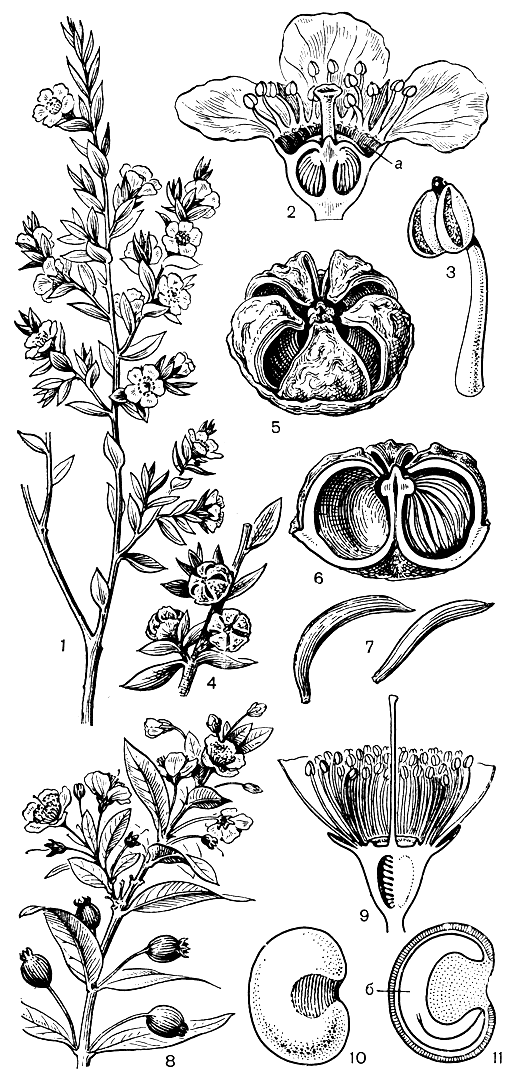
Рис. 110. Миртовые. Лептоспермум метловидный (Leptospermum scoparium): 1 - ветвь с цветками; 2 - продольный разрез цветка (а - нектарник); 3 - тычинка; 4 - ветвь с плодами; 5 - зрелый плод; 6 - продольный разрез плода; 7 - семя. Мирт обыкновенный (Myrtus communis): 8 - ветвь с цветками и плодами; 9 - продольный разрез цветка; 10 - семя; 11 - продольный разрез семени (б - зародыш)
Миртовые входят в состав самых разных растительных сообществ. Их можно встретить в дождевых лесах и пустынях, на каменистых горных склонах и пустошах, высоко в горах и на затопляемом морском берегу. Они образуют чистые леса и кустарниковые заросли, могут быть преобладающей породой в смешанном лесу, растут группами или единичными экземплярами в различных растительных сообществах. Монотипный палеотропический род осборния (Osbornia octodonta) является одним из специализированных мангровых растений и снабжен пневматофорами.
Миртовые - деревья и кустарники, среди которых имеются все переходы от гигантских деревьев, как некоторые виды эвкалипта (Eucalyptus, рис. 111) до мелких ползучих кустарничков. Листья супротивные или редко очередные, простые, обычно цельные. Они бывают округлыми, продолговатыми, ланцетными или игольчатыми. Размеры листьев варьируют от 1-2 мм у видов рода беккея (Baeckea) до 50 см у некоторых видов эвкалипта. Большинство миртовых имеет в молодом возрасте четырехгранные стебли.
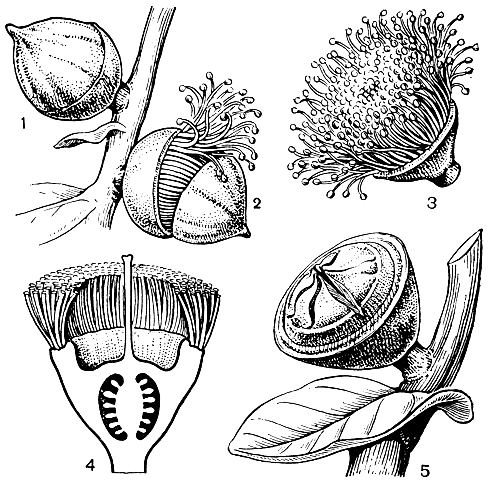
Рис. 111. Эвкалипт крупноплодный (Eucalyptus macrocarpa): 1 - бутон; 2 - раскрывающийся бутон и опадение крышечки; 3 - цветок; 4 - продольный разрез цветка; 5 - плод
Цветки миртовых собраны в разного рода верхоцветные или реже бокоцветные соцветия (табл. 32). В некоторых линиях эволюции миртовых соцветия подвергаются самым различным, иногда очень своеобразным видоизменениям, связанным с биологией опыления. Очень своеобразно, например, чрезвычайно похожее на корзинку сложноцветных щитковидное соцветие западноавстралийского монотипного рода актинодиум (Actinodium), у которого роль "язычковых" цветков играет несколько родов видоизмененных стерильных цветков. Цветки актиноморфные или очень редко зигоморфные, обоеполые или редко однополые, обычно с двумя прицветничками у основания. Чашелистиков и лепестков по 4-5, свободных или более или менее сросшихся; в некоторых случаях чашелистики сильно редуцированы и незаметны, а лепестки иногда отсутствуют. Весь околоцветник, или только чашечка, или венчик срастаются, образуя так называемую калиптру (или колпачок), опадающую при раскрывании цветка. У некоторых родов, как, например, плиния (Plinia) и каликоректус (Calycorectus), доли чашечки более или менее срастаются, но при раскрывании цветка расщепляются. Иногда, как у некоторых видов марлиереи (Marlierea), одна увеличенная доля чашечки становится колпачковидной. У некоторых видов сизигиума (Syzygium) доли околоцветника соединяются, образуя колпачок, который при раскрывании цветка опадает как целое. Хорошо развитая калиптра возникла в нескольких разных линиях эволюции миртовых и обычно связана с хорошо развитыми тычинками, как, например, у эвкалипта. Чаще всего калиптра образуется чашелистиками, но, как показал ряд австралийских ботаников (Л. Д. Прайор и Л. А. С. Джонсон, 1971; Л. А. С. Джонсон, 1972, 1976), у эвкалипта и близких родов калиптра образуется из венчика (при наличии свободных чашелистиков), из чашечки (при отсутствии венчика или при наличии более или менее свободных лепестков, опадающих вместе с калиптрой) и из сросшихся зачатков чашечки и венчика. Наконец, у целого ряда видов эвкалипта имеются две калиптры, образованные одна из чашечки, а другая - из венчика, причем обычно первая опадает раньше. Интересно также, что у некоторых родов сросшиеся или слипшиеся прицветнички могут образовать подобие калиптры. Все это показывает, что одна и та же биологическая задача может решаться разными путями с использованием разных морфологических структур. Тычинки обычно многочисленные, свободные или в пучках (табл. 32). Цветки, у которых лепестки сохраняются при раскрывании, имеют обычно небольшое число тычинок, равное или в два раза превышающее число лепестков. У цветков с опадающим при раскрывании околоцветником тычинок много и они часто образуют пучки разного вида; у одних эти пучки напоминают перо, у других пучок тычинок ветвится как ствол дерева (например, у тристании - Tristania), у третьих образует веер. В цветке может быть до 5 пучков с 20 и более тычинками в каждом пучке. Многочисленные тычинки имеют яркую, часто контрастную окраску тычиночных нитей и пыльников, которые и образуют привлекающий аппарат цветка. Гинецей вторично-синкарпный, редко паракарпный, состоит из 2-3 (редко до 16) плодолистиков. Столбик длинный, тонкий, с небольшим головчатым рыльцем. Завязь обычно полунижняя (только у псилоксилона - Psiloxylon - монотипного рода, эндемичного для островов Маврикий и Реюньон, завязь вполне верхняя, но этот род нередко выделяется в отдельное семейство), 1-16-гнездная, с 2 - многими семязачатками на каждой плаценте.
Миртовые почти исключительно перекрестноопыляемые растения, хотя есть сведения, что у некоторых видов австралийского рода дарвиния (Darwinia) наблюдается самоопыление. Цветки миртовых протандричны. Опылителями являются как насекомые, так и птицы и млекопитающие. Насекомые, как правило, опыляют мелкие цветки с заметным околоцветником и небольшим числом тычинок, но они охотно посещают также виды эвкалипта, метросидероса (Metrosideros), мелалеуки (Melaleuca) и другие, у которых только многочисленные и яркие тычинки служат для привлечения опылителей. Особенно много насекомоопыляемых миртовых в склерофильных верещатниках и скрэбах. Цветки, окрашенные в красный цвет, особенно цветки с многочисленными и умеренно длинными тычинками, обычно опыляются птицами. Особенно велика роль птиц в опылении миртовых в Австралии и Новой Зеландии, где число специализированных насекомых-опылителей, извлекающих свою пищу из цветков, невелико, особенно число высших пчелиных. Поэтому большую роль в опылении миртовых играют здесь попугаи лори (Psittacidae из подсемейства Loriinae), цветоносовые (Dicaeidae), нектарницевые (Nectariniidae), белоглазковые (Zosteropidae) и медососовые (Meliphagidae). Как указывает австралийской ботаник Дж. А. Армстронг (1979), орнитофилия наблюдается во всех экологических условиях, даже в пустыне. Интересно, что в Южной Африке, где эвкалипты прекрасно растут, постоянными опылителями стали черноголовые иволги и желтые ткачи, которым нектар эвкалиптов пришелся по вкусу. Не менее интересно, что в Ленинграде, в оранжереях Ботанического института Академии наук СССР нектар каллистемона пьют воробьи, производя опыление. К постоянным опылителям миртовых принадлежат также различные мелкие нелетающие млекопитающие (в Австралии особенно сумчатые). Некоторые из них имеют хоботковидно вытянутую мордочку, которую они погружают в цветок, поедая его сладкие части и нектар. Таков обитающий на юго-западе Австралии хоботноголовый кускус, или медовый поссум (Tarsipes spencerae), опыляющий цветки различных миртовых, особенно бофортии тычинковой (Beaufortia anisandra) и калотамнуса изящного (Calothamnus gracilis). Обитающий на юго-востоке и в центральных районах Австралии тонкохвостый кускус, или карликовый поссум (Cercaertus concinnus), опыляет цветки ряда видов эвкалипта и бофортии. Миртовые, приспособленные к опылению не летающими, млекопитающими, характеризуются крупными цветками и соцветиями с сильно выраженным запахом и обильным выделением нектара. Активное участие в опылении миртовых принимают также представители отряда рукокрылых - крыланы и летучие мыши. В Австралии виды крыланов (семейство Pteropidae), особенно виды летучих лисиц (Pteropus), добывают нектар из многих видов эвкалиптов, из видов ангофоры, мелалеуки, каллистемона и некоторых других миртовых. Цветки далеко не всегда бывают узкоприспо-соблены только к определенному типу опылителей. Так, в Австралии цветки мелалеуки пятинервной (M. qninquenervia) в течение дня регулярно посещаются насекомыми и птицами, а ночью сосущим нектар длинноязычным австралийским крыланом (Synonycteris australis). Этот крошечный крылан, длина тела которого всего лишь 5-6 см, с длинным подвижным языком питается почти исключительно цветками и нектаром. Он очень охотно посещает цветки мелалеуки, а также цветки каллистемона.

Таблица 32. Миртовые: 4 - каллистемон жестколистный (Callistemon rigidus), Батумский ботанический сад
Плоды миртовых очень разнообразны по форме и размерам (табл. 32). Сочные плоды отличаются яркостью окраски: они могут быть красные, оранжевые, желтые, голубые и черные за счет цвета экзокарпия. Эти плоды распространяются в основном приматами и птицами. У коробочек и орехов покровы становятся кожистыми или одревесневшими. Предполагают, что появление этого признака связано с обитанием в условиях частых низовых пожаров и засухи. С другой стороны, пожары способствуют прорастанию семян, высвобождая их из одревесневших плодовых оболочек. Созревание плодов и развитие семян длится от нескольких недель до нескольких лет. Коробочки растрескиваются обычно сверху вниз на разную глубину, причем механизм разбрасывания семян очень варьирует. У мангрового рода осборния плоды утрачивают сочный околоплодник, но остаются нераскрывающимися. Они распространяются по воде. Сухая ягода с ложными перегородками образуется при разделении гнезд завязи на многочисленные перегородки (родомиртус - Rhodomyrtus). Сухие плоды распространяются чаще всего птицами. Размеры семян от 0,5 до 22 мм и более. Семена бывают палочковидные (мелалеука, эквалипт), слегка изогнутые (лептоспермум), овальные или почковидные (мирт), иногда крылатые (пурпуреостемон - Purpureostemon), с гладкой, шершавой или ребристой поверхностью. Зародыш прямой, изогнутый, звездчатый, с узкими тонкими или толстыми семядолями и слабо развитым эндоспермом. По строению зародыша многие миртовые хорошо различаются.

Таблица 32. Миртовые: 5 - мелалеука зверобоелистная (Melaleuca hypericifolia), оранжереи Ботанического института АН СССР в Ленинграде
Представители миртовых резко различаются по характеру плода: одни имеют сухие плоды, другие - сочные. Сухие плоды - орехи или коробочки - характерны для около 80 родов, объединяемых в подсемейство лептоспермовые (Leptospermoideae). Ареал этого подсемейства ограничен Австралией, отдельные роды достигают Малезии и Южной Азии и лишь единственный род тепуалия (Tepualia) встречается в Южной Америке (Чили). Сочноплодные миртовые объединяют более 60 родов в подсемейство миртовые (Myrtoideae), у которых очень разорванный ареал: тяготея к тропической Америке, они растут также в Африке, Южной и Юго-Восточной Азии, Северной Австралии и являются более мезофильными растениями.

Таблица 32. Миртовые: 3 - лептоспермум прутьевидный (Leptospermum scoparium), Батумский ботанический сад
В подсемействе лептоспермовых наиболее крупный род - эвкалипт, включающий около 525 видов, распространенных в основном в Австралии и Тасмании с немногими видами в Малезийской флористической области. Эвкалипты - самые высокие растения среди цветковых. Были описаны экземпляры высотой до 155 м при диаметре ствола 25 м. Теперь таких гигантов уже нет, но 100-метровые деревья в возрасте 350-400 лет еще недавно украшали влажные леса Австралии. Стройные, высокие, прямоствольные виды эвкалипт царственный (E. regnans), эвкалипт гигантский (E. gigantea) растут во влажном климате, образуя верхний ярус в смешанном лесу. После пожаров возникает чистый эвкалиптовый лес, но без эвкалипта царственного, так как он не выносит пожаров. Эвкалипты опережают по скорости роста все породы: к концу первого года высота сеянцев достигает 2,5 м, а к 10 годам - 25-30 м. В сухом жарком климате растут более низкорослые виды, а в пустынях Центральной Австралии карликовые эвкалипты (их более 130 видов), едва достигающие 2-3 м в высоту. Они образуют вместе с акациями густые заросли малли. Из кустарниковых эвкалиптов состоит преимущественно и другой характерный тип растительности Австралии - скрэбы. Корявым деревцем растет высоко в горах, где выпадает снег, эвкалипт снеголюбивый (E. niphophila), на каменистопесчаных почвах образует плотные заросли эвкалипт камеденосный (E. gummifera). Эвкалипты своеобразно приспособились к жизни и размножению в условиях частой засухи, резкой смены температур, плохого корневого питания и пожаров. У молодых растений на гипокотиле образуются вздутия - древесные клубни (лигнотуберы), которые с возрастом увеличиваются в размерах. Из лигнотубер возникают многочисленные побеги. У эвкалипта камеденосного обнаружен клубень, занимающий площадь 75 м2, на нем насчитали 301 живой побег и 70 мертвых. Большую часть клубня занимает сердцевина, содержащая крахмал и неорганические вещества, имеются также камбий, флоэма, корка, луб. Эвкалипты - вечнозеленые жестколистные растения. Только 10 видов теряют листья в сухой жаркий период - это обитатели северных областей Австралии.

Таблица 32. Миртовые: 1 - эвкалипт Джили (Eucalyptus gillii), оранжереи Ботанического института АН СССР в Ленинграде
Листья эвкалипта цельные, у молодых растений супротивные, сидячие, стеблеобъемлющие. У взрослых листорасположение очередное, листовая пластинка на длинных черешках повертывается всегда параллельно падающим солнечным лучам. Именно поэтому эвкалиптовые леса являются светлыми, с хорошо развитым травянистым покровом. Палисадная ткань расположена на обеих сторонах листа. Листья варьируют по форме от круглых до саблевидных. Они содержат множество эфирномасляных желёзок. Корковый слой, покрывающий ствол, может быть гладким, глубокобороздчатым или чешуйчатым. У некоторых эвкалиптов в период сильного роста корковый слой ежегодно отделяется, что придает стволу очень своеобразный вид: корка висит в виде больших лоскутов или мелких лохмотьев. Раскрывшийся цветок эвкалипта не имеет околоцветника, так как тычинки, распрямляющиеся при раскрывании бутона, сбрасывают калиптру. Тычинок много, с длинными тычиночными нитями, белыми, реже желтыми, розовыми или красными, прикрепленными по краю вогнутого цветоложа. Завязь 2-7-гнездная с многочисленными семязачатками. У ряда видов описаны функционально однополые цветки. Цветение наступает у эвкалиптов через 12-18 месяцев после заложения бутонов, что оправдывает необходимость калиптры как защиты. Плоды овальные или шаровидные, с гладкой или ребристо-бугорчатой поверхностью, диаметром от нескольких миллиметров до нескольких сантиметров. Они созревают около года, но остаются на материнском растении по нескольку лет, после чего коробочки вскрываются. Деревянистыми плодами эвкалиптов питаются попугаи, способствуя их расселению.
Эвкалипт оказался одним из полезнейших родов семейства миртовых. Он отличается ценной древесиной; благодаря способности к быстрому росту его разводят почти во всех тропических и субтропических странах. Кроме того, практическое значение имеет кора эвкалиптов, содержащая дубильные вещества, а также листья, из которых извлекают ценное эфирное масло, широко употребляемое в медицине, парфюмерии и промышленности. В СССР культивируют около 30 видов эвкалипта, главным образом на Черноморском побережье Кавказа.
Интересен род метросидерос (около 60 видов), распространенный от Южной Африки до Полинезии и Новой Зеландии. Вечнозеленые кустарники, деревья и деревянистые лианы. Есть эпифитные формы. Цветки с многочисленными тычинками, у которых широкий связник с несколькими масляными желёзками. Древесина некоторых крупных деревьев не уступает по качеству железному дереву.
Австралийско-новокаледонский род каллис-темон включает около 25 видов, из которых большинство произрастает в Австралии. Яркие тычинки, составляющие основную часть цветков, собраны в плотные соцветия длиной 5-12 см и шириной 3-6 см. В зависимости от окраски тычиночных нитей соцветия бывают густокрасными, желтыми или кремовыми. В роде встречаются как деревца, так и кустарники, с торчащими ветвями и ланцетовидными листьями.
Пучками ярких тычинок образованы также розовато-лиловые цветки другого преимущественно австралийского рода - мелалеуки, включающего более 100 видов. Подавляющее большинство видов ограничено Западной Австралией, где образуют великолепно цветущие заросли. Это кустарники, реже деревья высотой до 9 м. Кроны деревьев очень плотные и тенистые, так что под их пологом нет растительности. Листья от продолговатых до узколинейных, почти нитевидных. В листьях образуются ароматические эфирные масла. Цветки собраны плотной головкой или "ершиком". Репродуктивные побеги мелалеуки и каллистемона прорастают в вегетативную ветвь, на которой опять появляются цветки. Плоды одревесневающие, остающиеся на ветвях по нескольку лет. В Восточной Австралии, Юго-Восточной Новой Гвинее и Новой Каледонии обитает так называемое каепутовое дерево (Melaleuca quinquenervia, часто неправильно называемое M. leucadendron). Этот вид часто культивируется как эфиромасличное растение. Из листьев и молодых ветвей получают эфирное масло, применяемое в медицине и как инсектицид. Культивируется также для осушки болот и как декоративное растение.
Из представителей подсемейства лептоспермовых назовем также род лептоспермум (Leptospermum, табл. 32). В нем более 40 видов вечнозеленых кустарников и деревьев, обитающих главным образом в Австралии, и только 1-2 вида встречаются на Малайском архипелаге и 3 вида в Новой Зеландии. Лептоспермум сглаженный (L. laevigatum) широко используется в Австралии для закрепления песков. Некоторые виды имеют декоративное значение.
Широкой известностью пользуется мирт, по имени которого названо семейство и подсемейство миртовые. В современном более узком его понимании род мирт заключает не более 16 видов, один из которых встречается, как уже было сказано, на Азорских островах, в Европе, Северной Африке и Западной Азии, другой - в Африке, а остальные - в Вест-Индии и Флориде. Однако, по мнению известного американского ботаника Р. Мак Во (1968), вест-индские виды стоят ближе к некоторым американским родам, чем к мирту обыкновенному. Поэтому не исключено, что род мирт сведется к двум афро-евразиатским видам. Это небольшие вечнозеленые деревья и кустарники. Цветки с 4 лепестками и многими тычинками. Плод - ягода, увенчанная остающимися чашелистиками. Мирт обыкновенный является одним из характерных элементов средиземноморского маквиса. С древнейших времен мирт обыкновенный культивируется главным образом ради эфирных масел, содержащихся в листьях и других частях. Зеленые и сухие плоды используются как приправа в кулинарии. В СССР его выращивают в Крыму и на Черноморском побережье Кавказа.
К мирту, особенно к вест-индским видам, близок большой род псидиум (Psidium, около 100 видов в Вест-Индии и в тропической Южной Америке). Вечнозеленые деревья или кустарники обычно с крупными белыми цветками с многочисленными тычинками и шарообразными или грушевидными плодами-ягодами, которые иногда бывают крупными и съедобными. Некоторые виды псидиума культивируются как плодовые деревья. Наиболее известна гуава, или "гуайава" (Psidium guajava), широко культивируемая в тропических и субтропических странах из-за богатых витаминами ароматных, кисло-сладких, сочных плодов, обладающих пищевыми качествами. Плоды гуавы используют для приготовления джемов, приправ, соков, пунша и особенно желе. Его семена широко распространяются птицами и другими животными и во многих местах он легко натурализуется. В СССР гуава имеется в коллекционных насаждениях Черноморского побережья Кавказа.
С миртом сближается также род родомиртус (Rhodomyrtus), включающий около 20 видов, распространенных от тропической Азии до Новой Каледонии и Австралии. Индо-малезийский вид родомиртус опушенный (R. tomentosa) культивируют ради съедобных плодов.
Почти половина всех видов семейства миртовых относится к двум родственным родам - евгении (Eugenia) и сизигиуму (Syzygium). Род евгения заключает около 1000 видов, произрастающих главным образом в тропической Америке. Лишь немногие виды этого рода встречаются в тропиках Старого Света. Вечнозеленые деревья и кустарники с простыми листьями. Чашечка 4-лопастная; лепестков 4, белых; завязь 2-гнездная; плод - ягода. Некоторые виды, например "суринамскую вишню" или евгению одноцветковую (E. uniflora), культивируют ради съедобных плодов. Палеотропический род сизигиум, насчитывающий от 400 до 500 видов, отличается от евгении характером соцветия, трубкой чашечки, продолженной выше завязи, свободными (не сросшимися) семядолями и семенной кожурой. Виды сизигиума - вечнозеленые деревья с простыми листьями. Лепестки свободные или сросшиеся в калиптру. Самым известным представителем рода является гвоздичное дерево, или сизигиум ароматный (S. aromaticum). Все части этого растения содержат желёзки с эфирным маслом, известным под названием гвоздичного масла. Ценность гвоздичного масла определяется наличием в его составе эвгенола. Его широко применяют как сырье для производства ванилина, используемого в парфюмерной промышленности и для приготовления некоторых лекарственных препаратов. Высушенные бутоны гвоздичного дерева, известные под названием гвоздики, используют как пряность. Гвоздичное дерево родом с Молуккских островов широко культивируют во многих тропических странах. В культуре известны и некоторые другие виды сизигиума, в частности юго-восточноазиатский вид, известный под названием сигизиум ямбоза (S. jambos). В тропиках широко культивируют как декоративное плодовое растение малайский вид сизигиум малаккский (S.malaccense) - одно из наиболее красивых тропических деревьев.

Таблица 32. Миртовые: 2 - сизигиум метельчатый (Syzygium paniculatum), оранжереи Ботанического института АН СССР в Ленинграде
Наконец, скажем несколько слов о небольшом южноамериканском роде фейхоа (Feijoa, табл. 32), состоящем всего из 2 видов. В последнее время некоторые ботаники объединяют фейхоа с другим южноамериканским видом, описанным на два года раньше фейхоа под названием акка (Асса). Однако не все ботаники с этим согласны, и австралийские ботаники Б. Бриггс и Л. А. С. Джонсон (1979) продолжают различать эти два рода. Наиболее важным представителем рода фейхоа является фейхоа Селлова (F. sellowiana). Это вечнозеленое деревце или кустарник с раскидистой густой кроной и серебристо опушенными листьями, обитающий дико во влажных субтропических и умеренно теплых частях Южной Бразилии, Парагвая, Уругвая и Северной Аргентины. Взрослые растения выдерживают температуру до -12 °С и устойчивы против засухи. Многочисленные темно-красные тычинки более длинные, чем мясистые лепестки, привлекают птиц. Плод фейхоа - зеленая с красноватым оттенком ягода. Мякоть зрелых плодов плотная, сочная, кисло-сладкая с приятным вкусом, напоминающим вкус гуави. Плоды используют в свежем и переработанном виде. Фейхоа культивируют как плодовое и декоративное растение во многих странах, в том числе на Черноморском побережье Кавказа и на Южном берегу Крыма. Имеются урожайные высококачественные сорта, некоторые из них выведены в СССР.

Таблица 32. Миртовые: 6 - фейхоа Селлова (Feijoa sellowiana), Адлер
Для человека имеют значение также другие представители семейства миртовых. Незрелые плоды многих миртовых употребляют как пряности. Таковы плоды американской пименты двудомной, или лекарственной (Pimenta dioica, или P. officinalis), называемые "ямайский душистый перец". Листья, цветки, бутоны, семена и кора многих миртовых находят применение в медицине. Отвары листьев и бутонов видов беккеи и сизигиума употребляют как тонизирующие напитки вместо чая. Некоторые миртовые являются ценными медоносными растениями. Особенно популярен у пчеловодов нектар из цветков эвкалипта разноцветного (E. diversiocolor). Но наибольшую ценность имеет древесина ряда миртовых. Особенно ценится на мировом рынке древесина эвкалиптов, являющаяся строительным материалом, сырьем для бумажно-целлюлозной промышленности, для производства лаков, клея и других веществ. Ряд видов имеет очень тяжелую и твердую древесину, устойчивую к гниению за счет высокого содержания дубильных веществ. Особенно твердую древесину имеет эвкалипт железнодревесный (E. syderoxylon), содержащий в клеточных стенках сосудов кремний.
Отдельные виды культивируют как декоративные растения из-за красивых цветков и плодов или свежей зелени листвы. Наибольшей известностью пользуется мирт, с которым связаны многие народные обряды и праздники. В горшечной культуре и закрытом грунте часто выращивают виды лептоспермума, мелалеуки, каллистемона.
Семейство комбретовые (Combbemaceae) (С. С. Морщихина)
Семейство комбретовых охватывает около 600 видов, относящихся к 20 родам. Это деревья, кустарники и деревянистые лианы, являющиеся существенным элементом во многих древесных формациях тропических стран обоих полушарий. Наибольшее число видов комбретовых во флоре тропической Африки. Наименее богата комбретовыми флора Австралии, в которой представлены только три рода: терминалия (Terminalia), люмнитцера (Lumnitzera) и макроптерантес (Macropteranthes). Самые крупные роды - комбретум (Combretum) и терминалия, объединяющие примерно по 250 видов каждый, широко распространены в тропиках обоих полушарий (но комбретум отсутствует в Австралии) и местами выходят за пределы тропиков.
В семействе комбретовых преобладают деревья средней величины и кустарники, обитающие в листопадных лесах, в саваннах и кустарниковых зарослях. Благодаря широкому экологическому диапазону в семействе есть вечнозеленые и в большей или меньшей степени листопадные виды. У некоторых видов, например у терминалии катаппа (T. catappa) листья перед опадением краснеют. Терминалия катаппа, широко распространенная в прибрежных лесах от Восточной Африки и Мадагаскара до Полинезии, сбрасывает листья дважды в году.
Среди комбретовых есть и могучие, высотой до 40-50 м, деревья с досковидными корнями, входящие в первый ярус тропических дождевых и муссонных лесов. Терминалии часто можно узнать по характерному облику кроны. Вершина дерева нарастает моноподиально, боковые ветви расположены почти горизонтально и несут на концах большую массу листьев. Известный английский ботаник Э. Дж. Корнер, долгие годы работавший в тропиках, метко назвал терминалии за форму их кроны деревьями-пагодами.
Многочисленны в семействе вьющиеся и лазящие лианы, многие из них закрепляются за опору колючками, которые представляют собой основание черешка опавшего листа. Комбретум метельчатый (C. paniculatum), комбретум кистевидный (C. racemosum) - лианы, широко распространенные в лесах тропической Западной Африки, поднимаются по деревьям на 20-метровую высоту.
Виды двух родов - лагункулария (Laguncularia) и люмнитцера (Lumnitzera) - произрастают в мангрове. Лагункулария встречается в мангрове бассейна Атлантического океана, т. е. в Центральной и Южной Америке и Западной Африке. Люмнитцера распространена от берегов Восточной Африки и Мадагаскара к востоку по побережьям тропической Азии, Северной Австралии до Полинезии. Лагункулария и люмнитцера - небольшие вечнозеленые деревья высотой около 6-10 м. Произрастают они на регулярно затопляемом приливами грунте, защищенные от волн открытого моря полосой зарослей ризофоры или авиценнии. Лагункулария имеет короткие ходульные корни, закрепляющие дерево в илистом грунте, и вертикальные спаржевидные "дыхательные" выросты корней с отрицательным геотропизмом. Газообмен, происходящий в их тканях, восполняет кислородный дефицит, который испытывает корневая система в затопляемом грунте. Такую же "дыхательную" функцию выполняют вертикальные коленчатые выросты, образующиеся на горизонтальных корнях люмнитцеры.
На побережьях Атлантического океана, во внутренней части мангровых зарослей и на открытых скалистых берегах встречается конокарпус прямостоящий (Conocarpus erectus, табл. 33) - небольшое вечнозеленое дерево, легко образующее придаточные корни в нижней части ствола по мере нарастания намываемого приливами грунта. Во Флориде встречается его северная форма с серебристым опушением, а на известняковых берегах Карибского моря - наскальная.

Таблица 33. Комбретовые, кипрейные и ризофоровые: 1 - конокарпус прямостоящий (Conocarpus erectus), остров Куба
Комбретовые имеют цельные листья, расположенные супротивно, мутовчато или спирально; прилистники отсутствуют. Однако Ф. Веберлинг (1960) установил наличие рудиментарных шиловидных прилистников на ранних стадиях развития у видов терминалии, бухенавии (Buchenavia) и аногейссуса (Anogeissus).
Для комбретовых характерно наличие желёзок на черешке листа или на пластинке у основания, по сторонам главной жилки или у края. У некоторых видов вся пластинка усеяна мелкими желёзками, отчего лист выглядит точечно-прозрачным. Не менее характерно для комбретовых опушение из простых или многоклеточных волосков, выделяющих оксалаты кальция. Строение волосков разнообразно и является систематическим признаком (рис. 112). Наличие многоклеточных волосков в виде щитовидных чешуек на ножке, присущее видам комбретума и тилоа (Thiloa), позволяет определять даже фрагменты их ископаемых остатков. Устьица у комбретовых аномоцитные. Членики сосудов с простой перфорацией.
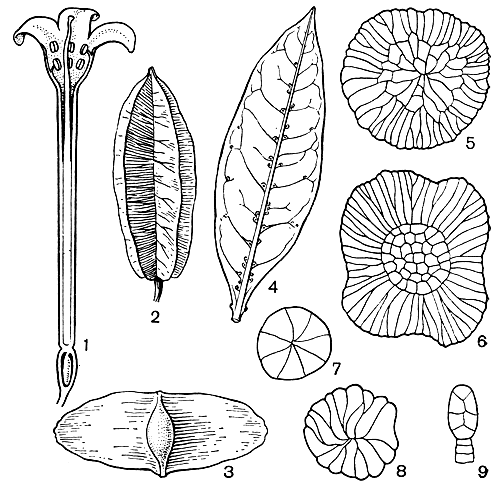
Рис. 112. Комбретовые. Квисквалис индийский (Quisqualis indica): l - продольный разрез цветка; 2 - плод. Терминалия великолепная (Terminalia superba): 3 - плод. Конокарпус прямостоящий (Conocarpus erectus): 4 - лист с желёзками. Различные типы волосков комбретума: 5 - комбретум кустарниковый (Combretum fruticosum); 6 - комбретум Илера (C. hilarianum); 7 - комбретум наскальный (C. rupicola); 8 - комбретум ланцетовидный (C. lanceolatum); 9 - комбретум десятитычинковый (C. decandrum)
Цветки комбретовых обычно довольно мелкие, невзрачные, белые, желтоватые или зеленоватые, иногда ярко-красные, у многих видов душистые, собраны в пазушные или верхушечные соцветия. Тип соцветий варьирует, преобладают кистевидные и колосовидные соцветия с удлиненной осью, одиночные или собранные в метелку. У видов конокарпуса и птелеопсиса (Pteleopsis) ось соцветия сильно укорочена, так что соцветие почти головчатого типа.
Для цветков комбретовых характерно наличие цветочной трубки, которую образует околоцветник, сросшийся в нижней части с завязью. Длина цветочной трубки варьирует, достигая у квисквалиса индийского (Quisqualis indica, табл. 33) 6-8 см (рис. 112). Цветки, как правило, актиноморфные (или слабозигоморфные), чашелистики и лепестки в равном числе, по 4-5 (редко 6-8). Цветки терминалии и близких к ней родов, а также тилоа (Thiloa), каликоптериса (Galycopteris) и немногих видов комбретума лишены лепестков. Тычинок обычно вдвое больше, чем долей околоцветника. У видов лагункуларии, люмнитцеры, конокарпуса и некоторых других родов цветки имеют лишь один круг тычинок. Тычинки с сильна развитым связником, который переходит в короткое острие.

Таблица 33. Комбретовые, кипрейные и ризофоровые: 2 - квисквалис индийский (Quisqualis indica), оранжереи Ботанического института АН СССР в Ленинграде
В цветках комбретовых хорошо развит нектарный диск, круглый или 5-10-лопастный, который обильно выделяет нектар, привлекающий насекомых, а у некоторых видов комбретума также и птиц. Цветки протогиничны, нередко рыльца торчат из еще не раскрывшихся бутонов. Гинецей из 4-5 плодолистиков, паракарпный, с длинным нитевидным столбиком. Завязь нижняя (только у видов стрефонемы - Strephonema - полунижняя), одногнездная, обычно с 2 (редко с 6 ) анатропными, свисающими с верхушки завязи на длинных фуникулусах семязачатками, из которых лишь один развивается в семя.
Плоды всегда односемянные, обычно нераскрывающиеся, большей частью сухие, ребристые, с 2-5 выступающими кантами или крыловидными выростами, которые способствуют распространению ветром. Такой тип плодов преобладает у видов комбретума (рис. 112), встречается у многих видов терминалии. Плоды прибрежных видов комбретума, лагункуларии, люмнитцеры, некоторых терминалии и др. обладают легким пористым пробкообразным околоплодником, который придает им плавучесть и обеспечивает распространение водой. Некоторые виды терминалии имеют крупные плоды с сочным околоплодником и твердым эндокарпием. Такой костянковидный тип плода встречается у видов терминалии, растущих по морским побережьям и в лесах вдали от моря. Самые крупные плоды имеет терминалия Кернбаха (T. kaernbachii) из лесов Малезии. Ее красные эллиптические плоды достигают 17 см в длину при диаметре около 6 см. Сочный околоплодник привлекает летучих мышей, которые участвуют в распространении плодов многих видов терминалии.
Семена комбретовых без эндосперма. Зародыш обычно со спирально свернутыми семядолями. Терминалия Кернбаха и некоторые другие виды с сочными плодами имеют круглый зародыш с 3 или 4 мясистыми семядолями (см. рис. 112), которые в семени сложены, как сегменты шара, и напоминают семядоли идиоспермума австралийского из семейства каликантовых. Для комбретовых характерно надземное прорастание семян, но у некоторых видов комбретума из саванн Западной Африки семена, прорастающие на поверхности, затягиваются первичным корнем в глубь почвы, что, очевидно, помогает проросткам выжить во время пожаров.
Комбретовым присуще наличие дубильных веществ во всех частях растений, что позволяет многие из них использовать в кожевенном производстве, в медицине, для получения красителей. Плоды некоторых видов терминалии из Южной Азии, известные под названием миробалан, имеют промышленное значение и являются предметом экспорта. Наибольшее значение в промышленности и медицине имеют терминалия хебула (T. chebula), плоды которой содержат 43-53% танина, и терминалия беллирийская (T. bellirica). Некоторые терминалии имеют съедобные плоды. Терминалию катаппа культивируют во всех тропических странах под названием "индийский миндаль". Ее семена содержат до 50% пищевого масла и используются в пищу как орехи. Многие представители семейства дают ценную строительную древесину. Особенно ценится древесина терминалии войлочной (T. tomentosa) - обычного спутника тикового дерева в муссонных лесах Индии. Для мебели и художественных паркетов используют древесину так называемого "желтого махагони", терминалии великолепной (T. superba) из лесов Западной Африки. Во многих странах выращивают в садах декоративный лазящий кустарник квисквалис индийский, длинные трубчатые цветки которого белые при распускании меняют окраску и становятся ярко-красными.
Семейство кипрейные (Onagraceae) (Л. И. Иванина)
Семейство заключает около 25 родов и до 700 видов. Большинство кипрейных сосредоточены в Новом Свете, в основном в западных областях Северной Америки и Мексики. Лишь представители немногих родов распространены по всему свету или только в тропиках.
Кипрейные - однолетние и многолетние травы, редко кустарники и небольшие деревья. Сравнительно высокие (до 30 м) деревья встречаются среди видов центрально-североамериканского рода хауя (Hauya). Листья кипрейных очередные или супротивные, реже мутовчатые, простые, цельные, преимущественно ланцетные и продолговато-яйцевидные средних размеров, цельнокрайные или зубчатые, без прилистников или реже с очень небольшими, обычно опадающими прилистниками. В семействе преобладают сравнительно крупные, красные, фиолетовые, розовые или желтые и белые обоеполые (редко однополые) цветки. Они расположены по одному (реже по два) в пазухах листьев или собраны в бокоцветные соцветия - олиственные кисти, колосья или метелки. Прицветнички отсутствуют, за исключением некоторых людвигий (Ludwigia, рис. 113), имеющих цветки с 2 прицветниками. У большинства кипрейных цветки актиноморфные, реже слегка зигоморфные, но у центральноамериканских лопесий (Lopezia, рис. 114) ясно зигоморфные. Они обычно 4-членные, иногда бывают 3-7-членные, а у двулепестника, или цирцеи (Circaea, рис. 115), цветки 2-членные. Цветочная трубка, приросшая к завязи, нередко продолжена над завязью. Чашечка в почкосложении створчатая, по отцветании обычно опадающая. Венчик в почкосложении скрученный, большей частью с яйцевидно-округлыми лепестками, часто суженными при основании в ноготок. У мексиканского монотипного рода ризенбахия (Riesenbachia) и у некоторых видов фуксий (Fuchsia) венчика нет вовсе, а его функцию при опылении выполняет обычно ярко окрашенная чашечка (табл. 33). Тычинки в том же числе, что и лопасти чашечки, или чаще в двойном против долей венчика и преимущественно расположены в двух кругах, редко тычинок две или одна. Пыльцевые зерна фуксий, энотеры (Oenothera, см. рис. 115) около полюса снабжены длинной висциновой нитью, которая, очевидно, способствует переносу пыльцы опылителями. Гинецей вторичносинкарпный, состоит обычно из 4 плодолистиков. Столбик простой, головчатый или с 4-лучевым рыльцем. Завязь всегда нижняя. Число гнезд завязи обычно соответствует числу чашелистиков.

Таблица 33. Комбретовые, кипрейные и ризофоровые: 3 - фуксия магелланскас (Fuchsia magellanica), оранжереи Главного ботанического сада в Москве
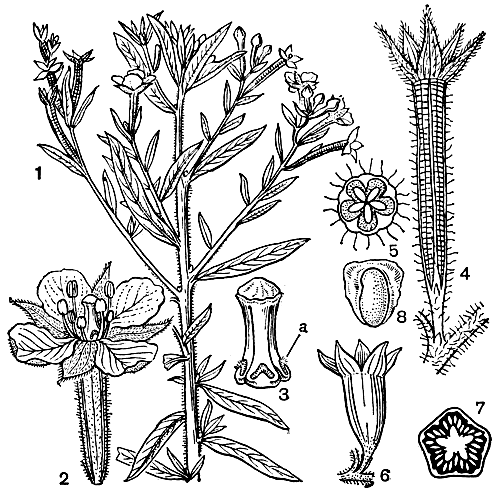
Рис. 113. Кипрейные. Людвигия узкошовная (Ludwigia stenoraphe): 1 - ветвь с цветками и плодами; 2 - цветок; 3 - столбик и рыльце (а - нектарники). Людвигия тонкоплодная (L. leptocarpa): 4 - плод; 5 - поперечный разрез плода. Людвигия многолетняя (L. perennis): 6 - плод; 7 - поперечный разрез плода; 8 - семя
Кипрейные - перекрестноопыляемые и реже самоопыляемые растения. Их опылители - насекомые (преобладают среди них пчелиные и чешуекрылые), а для фуксий главным образом колибри. У рода людвигия (75 видов, обитающих в обоих полушариях, преимущественно в субтропических и тропических областях) число чашелистиков от 3 до 7, чашечка не опадающая после цветения. Лепестки их желтые или белые, в том же числе, что и чашелистики, или отсутствуют. Трубка венчика не продолжена над завязью, столбик короткий, с головчатым или полушаровидным рыльцем, окруженный при основании нектарными желёзками. По данным американских исследователей Дж. Эстеса и Р. Торна (1974), цветки людвигии пеплоидной (L. peploides), произрастающей в небольших водоемах в штатах Оклахома и Техас, посещают около 30 видов насекомых. Для этого вида более характерна гейтоногамия (перенос пыльцы с других цветков того же растения). Основные опылители - перепончатокрылые. По поведению на цветке они распределяются на три группы. К первой группе относятся насекомые, садящиеся на рыльце, чтобы собрать пыльцу и нектар. Это самые крупные насекомые из пчелиных. Во вторую группу входят насекомые (в основном галиктовые - Halictidae), приземляющиеся на пыльники, чтобы собрать пыльцу. Двигаясь от пыльника к пыльнику, они пересекают рыльце и, также как насекомые первой группы, обычно участвуют в перекрестном опылении. Третья группа - это самые маленькие насекомые из пчелиных. Они собирают пыльцу, двигаясь вокруг пыльников, и обычно в контакт с рыльцем не вступают, т. е. при опылении они не, эффективны.
Цветки кипрея узколистного, или иван-чая (Epilobium angustifolium), некоторыми ботаниками относимого к особому роду хамерион (Chamerion), обычно служат классическим примером протандрии. Именно при наблюдении цветения этого вида немецким ботаником X. К. Шпренгелем была открыта дихогамия растений и описана в его сочинении, изданном в 1793 г., "Разоблаченная тайна природы в строении и оплодотворении цветков". Основные опылители иван-чая - пчелы, но нередко цветки посещают шмели, бабочки и другие насекомые. Темно-розовые высокие соцветия иван-чая насекомые замечают издали. Растения отличаются большой продолжительностью цветения (1-2 месяца), а цветки - обилием нектара. За время цветения один цветок выделяет от 0,5 до 12 мг (а иногда до 26) нектара. Нектарники расположены между расширенным основанием тычиночных нитей и основанием столбика и представляют собой темно-зеленое кольцо. Они возникают из меристематических клеток верхней части завязи и основания тычиночных нитей. В открытом цветке иван-чая легко заметить 4 ланцентных сиренево окрашенных чашелистика, столько же обратно-яйцевидных, слегка суженных в ноготок лепестков, 8 тычинок (у 4 тычинок нити немного длиннее) и 1 пестик. Первыми в цветке созревают розово окрашенные пыльники на сравнительно длинных и тонких нитях. В это время недлинный столбик с нераскрытым рыльцем изогнут книзу. После того как пыльники отпылили, столбик удлиняется почти вдвое, а рыльце раскрывается четырьмя растопыренными лопастями. Пчелы или другие насекомые, обычно вымазавшиеся пыльцой в более молодых цветках, при взятии нектара касаются рыльца, и так происходит перекрестное опыление.
У лопесий (Lopezia, 17 видов, обитающих в Мексике и Северной Америке) цветки зигоморфные с сильно измененными частями в связи с приспособлением к опылению насекомыми. Так, у лопесий увенчанной (L. coronata) чашелистики располагаются так, что один направлен вперед, а 3 других назад (см. рис. 114); лепестков тоже 4, но задние уже двух передних и согнуты коленом, с медовой желёзкой (медовиком) в изгибе. Из двух тычинок только одна плодущая, а вторая лепестковидная, сложенная лодочкой. В ранней стадии открытого цветка пестик не развит и его столбик вместе с одной плодущей тычинкой заключен в стаминодий (лепестковидную тычинку). В более раскрытом цветке тычинки занимают середину, цветка. Когда насекомое опускается на такой цветок за нектаром, то оно наталкивается на зрелый пыльник и вымазывается пыльцой. В зрелом цветке столбик занимает место тычинок, и насекомое, которое уже посетило молодой цветок и вымазалось пыльцой, подлетая к цветку, соприкасается с рыльцем и опыляет цветок.
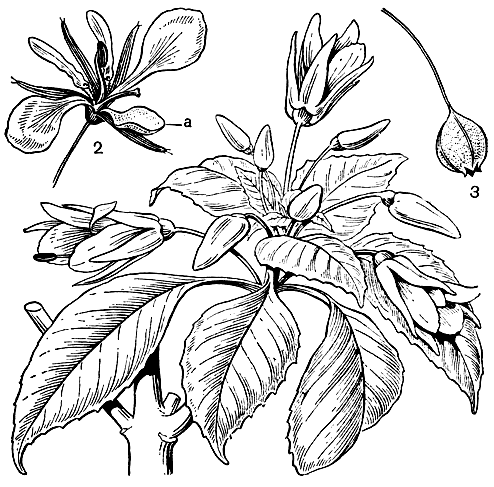
Рис. 114. Кипрейные. Лопесия крупнолистная (Lopezia macrophylla) 1 - ветвь с цветками. Лопесия увенчанная (L. coronata): 2 - цветок (а - лепестковидный стаминодий); 3 - плод
Цветки энотеры двулетней (Oenothera biennis), раскрывающиеся только перед заходом солнца, приспособлены к опылению ночными бабочками и пчелиными. Их крупные желтые венчики с длинной трубкой сравнительно хорошо заметны в сумерках. Цветки протандричные. Сначала развиваются тычинки и стоят в первый вечер с раскрытыми пыльниками. Бабочки, опустив хоботок в трубку венчика, вымазываются пыльцой. Интересные наблюдения над опылением пчелиными энотеры были проведены в различных районах Северной Америки П. Рейвеном с сотрудниками (1963). В пустыне Колорадо 9 видов энотер опыляются 21 видом пчелиных, а в Большом Бассейне 10 видов энотер опыляются 61 видом пчелиных. В Европе большинство видов энотеры - самоопыляющиеся растения. Мелкие цветки цирцей, или двулепестника, опыляются преимущественно мухами. В этом роде 12 видов, распространенных в основном в умеренной зоне. Название рода происходит от имени греческой мифологической волшебницы, упоминаемой Гомером в "Одиссее". Цветки цирцей парижской (С. lutetiana) приманивают маленьких мух. Если мухи уже с пыльцой (принесенной с другого цветка), то при посадке на цветок они обязательно коснутся рыльца. Передними ногами мухи обычно охватывают основания тычинок и подгибают пыльники под свое тело. Таким образом они опять нагружаются пыльцой и улетают с ней на другой цветок. У кипрея малоцветкового (Epilobium parviflorum), распространенного в Евразии от Атлантической Европы до Гималаев, цветки небольшие, светло-розовые, почти белые. 4 тычинки этого вида окружают рыльце, и пыльца собственного цветка попадает на него - происходит самоопыление.
Среди орнитофильных растений фуксии, как указывает ряд авторов, принадлежат к одной из самых древних групп. Цветки их относятся к трубчатому типу. Они ярко окрашенные (обычно красные), с окрашенной чашечкой, длинными 8 неравными тычинками, богатые нектаром и пыльцой. Некоторые виды фуксии опыляются ветром и насекомыми. Фуксии обитают в Центральной и Южной Америке (а несколько видов встречаются на островах Новой Зеландии и Таити).
У большинства кипрейных плод - многосемянная вскрывающаяся коробочка, реже ягода (фуксия) или ореховидный (цирцея).
Распространение семян и плодов происходит преимущественно анемохорно и зоохорно. Семена кипреев (Epilobium) снабжены длинным хохолком из волосовидных придатков - парашютом. Такие растения-анемохоры имеют некоторое превосходство при захвате свободных территорий. Кипрей, очевидно, способны расселяться как мигрирующие виды и долго сохранять всхожесть в почве. Этим, по-видимому, объясняется тот факт, что большие гари в лесу заселяются иван-чаем. Ветром разносятся коробочки и семена у ряда других кипрейных. У энотер коробочки иногда имеют значительные крылатые ребра, а семена снабжены кожистой оторочкой (см. рис. 115). Семена древесных видов рода хауя представляют собой как бы удлиненное ушкообразное крыло, которое легко может подхватываться ветром. Плоды у цирцей более или менее покрыты прочными крючковидно загнутыми волосками - выростами эпидермальных клеток. Такие плоды приспособлены к пассивному переносу их на поверхности тела животных (эпизоохория). Распространение семян у ягодообразных плодов фуксий производят в основном птицы.
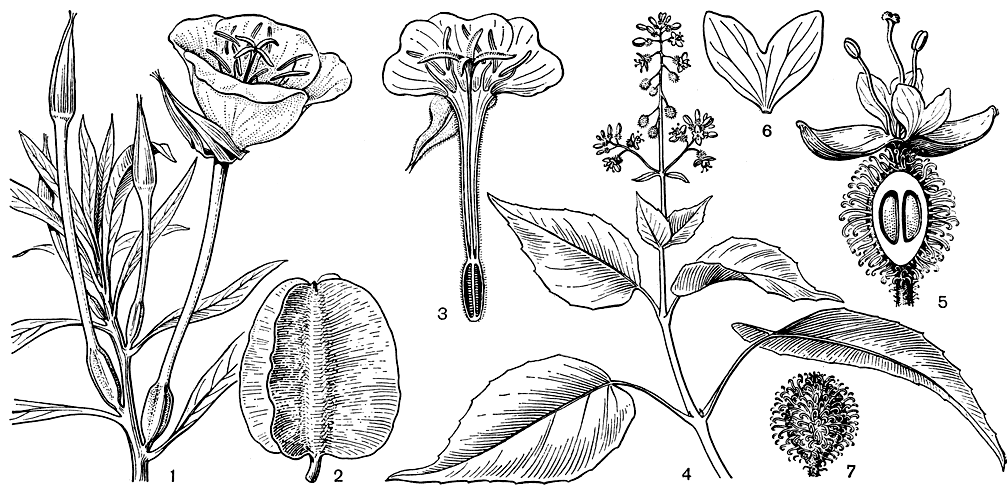
Рис. 115. Кипрейные. Энотера миссурийская (Oenothera missouriensis): 1 - ветвь с цветком; 2 - плод. Энотера двулетняя (O. biennis): 3 - разрез цветка. Цирцея сердцевидная (Circaea cordata): 4 - верхняя часть растения; 5 - цветок; 6 - лепесток; 7 - плод
Кипрейные по своей экологии (а также и морфологии и анатомии) преимущественно мезофиты или гигрофиты. Они растут главным образом по берегам рек и ключей, у канав, на пойменных лугах, во влажных лесах, на ключевых болотах, на вырубках и по гарям. Многие виды встречаются в горах от верхнего лесного до альпийского пояса. Так, в альпийском поясе Закавказья и в Иране можно встретить кипрей холодный (E. algidum), широко распространен в Евразии арктоальпийский кипрей альпийский (E. alpinum), на Памире около горячих ключей растет кипрей теплолюбивый (E. thermophilum). Энотеры и гауры (Gaura) обитают в прериях и в пустынях Северной Америки. В горных южноамериканских лесах (в Боливии до 3500 м над уровнем моря) произрастают многие виды фуксий. Среди людвигий встречаются виды, приспособленные к водно-болотному образу жизни. Наряду с обычными корнями (положительно геотропичными), погруженными в ил, у них имеются прямостоячие, вальковатые дыхательные корни (отрицательно геотропичные), в которых развита воздухоносная ткань аэренхима. В ее пустотах всегда находится воздух. Это запас кислорода, необходимый для тех частей растения, которые остаются погруженными в воду. Кроме того, растения снабжены еще и плавательными корнями.
Наиболее крупные роды в семействе - кипрей (около 200 видов), энотера (около 120 видов) и фуксия (около 100 видов). Кипрей - космополит, но его виды преимущественно обитают в горах Евразии (80 видов), в Северной Америке (45 видов), значительное число видов распространено в Новой Зеландии. Кипрей - единственный представитель семейства (исключая заносных), встречающийся на материке Австралия. Среди кипреев много гибридов. Энотеры также легко образуют гибриды и служат нередко объектами генетического и цитологического изучения. Энотера Ламарка (O. lamarkiana) была привлечена в опытах X. Де Фриза при исследовании мутаций у растений. Многие виды энотеры занесены из Америки в Евразию и на другие континенты. В СССР встречаются 11 видов, в том числе энотера двулетняя, занесенная в Европу в 1614 г.; позднее она распространилась как сорное.
Многие кипрейные встречаются в культуре как декоративные. Это прежде всего кларкии (Clarkia, 36 видов), растущие в западной части Северной Америки и в Чили. Среди кларкий наиболее известны прекрасные обильно цветущие однолетники кларкия ноготковая (C. unguiculata), более известная в садоводстве под названием кларкии изящной (C. elegans), и кларкия прелестная (C. amoena), обычно называемая годецией (Godetia). Оба вида представлены значительным разнообразием форм и сортов. В садоводческой практике также используют цаушнерию калифорнийскую (Zauschneria californica). Это низкий полукустарник, большими красными длиннотрубчатыми цветками схожий с фуксиями, но отличающийся от них плодами коробочками. Из рода гауры (18 видов) в садах часто разводят гауру двулетнюю (G. biennis).
Особое место среди декоративных кипрейных давно уже занимают фуксии (первые фуксии культивировали в Европе с 1806 г.). Название рода дано в честь выдающегося медика-ботаника XVI столетия Леонарда Фукса. Фуксия забытая (F. neglecta), фуксия магелланская (F. magellanica), фуксия щитковидная (F. corymbiflora) и многие другие послужили основой для создания более чем 2 тыс. сортов, среди которых много махровых.
Некоторые виды фуксий пригодны не только для комнатных культур: их выращивают во влажных почти субтропических районах Советского Союза. В этом отношении представляют интерес небольшие или карликовые фуксии главным образом из Мексики, Чили, Фолклендских островов и Новой Зеландии. Ягоды фуксий съедобные.
В жизни человека кипрейные особой роли не играют, хотя некоторые из них и обладают многочисленными полезными качествами. Иван-чай - один из лучших травянистых медоносов. С 1 га зарослей иван-чая можно получить до 600 кг прекрасного прозрачного меда. Его листья используют в лекарственных целях. Они содержат значительное количество дубильных веществ, витамина С. Иногда листья заваривают как чай - "капорский чай". Корни этого вида съедобны, слегка сладковатого вкуса. Иван-чай - ценное кормовое растение, имеющее высокое содержание каротина, протеина и жира. Особенно представляет интерес как пастбищное растение в горах и на севере Якутии - корм для северных оленей. Хорошими медоносами являются и такие виды
кипрея, как кипрей опушенный (E. hirsutum). Молодые листья некоторых кипреев употребляют как салат. Людвигию ползучую (L. repens) в некоторых областях используют как кормовое, лекарственное, пищевое, а людвигию болотную (L. palustris) для окраски ткани в желтый цвет. Энотеру двулетнюю используют не только как декоративное растение (ее разводят в садах из-за ароматичности цветков), но и как пищевое (мясистый корень в первый год жизни можно употреблять в пищу как овощ) и техническое (содержит около 28% масла).
Семейство рогульниковые, или водноореховые (Trapaceae) (В. Н. Васильев, А. П. Белавская)
В семейство рогульниковых входит только один современный род водяной орех (Trapa), распространенный в Евразии и Африке от умеренной зоны до тропиков. Водяной орех насчитывает около 30 видов. Однако некоторые авторы сводят число видов этого рода до одного полиморфного вида, в то время как В. Н. Васильев, напротив, доводит их число до 200. Это семейство стоит ближе всего к семейству кипрейных, особенно к роду людвигия (Ludwigia).
Латинское название trapa произошло от слова calcitrappa - рогатка. "Рога" водяного ореха отдаленно напоминают рогатку, употреблявшуюся древними римлянами против конницы.
Водяной орех имеет очень обширный географический ареал, но распространен отдельными островами. Современная северная граница отдельных изолированных зарослей водяного ореха проходит около 54-57° с. ш.
Водяной орех - травянистое однолетнее растение, хотя, по некоторым сведениям, может быть и многолетним (Флеров, 1925). Гибкий, плавающий в воде стебель прикреплен ко дну, как якорем, прошлогодним орехом, а также нитевидными буроватыми корнями (рис. 116). При подъеме уровня воды водяной орех может отрываться от грунта и превращаться в растение свободно плавающее. Плавая по мелководью и достигая подходящей глубины, он снова заякоривается и укореняется. Для водяного ореха, как и для многих других водяных растений, характерна гетерофиллия, или разнолистность. Прежде всего на стебле появляются нитевидные, рано опадающие листья. Позже по обе стороны листовых подушек вырастают длинные, рассеченные на волосовидные доли фотосинтезирующие органы. На водной поверхности водяной орех развивает одну или несколько розеток ромбических, зазубренных листьев, несколько напоминающих по форме листья березы. Листья расположены мозаично благодаря разной длине черешков. В верхней части черешков имеются так называемые "плавательные пузыри" - вздутия, заполненные воздухоносной тканью (аэренхимой). Цветки одиночные, в пазухах листьев, 4-членные, с белыми, изредка розоватыми (у африканских видов) прозрачными лепестками, до 8-10 мм в диаметре. Верхние доли чашечки одревесневают и превращаются при плодах в легко отламывающиеся колючки или шипы. Плод односемянный, костянкообразный, с каменистым эндокарпием, который и является твердой оболочкой ореха с 4 или 2 рогами - выростами эндокарпия. Семена без эндосперма, зародыш с неравными семядолями: одна большая и мясистая, а другая в виде чешуйки. При прорастании семени большая остается в плоде, а маленькая выходит наружу вместе с зародышевым корешком и стеблем. Зародышевый корешок водяного ореха, в отличие от корня всех прочих растений, начинает расти не вниз, а вверх и лишь после появления стебля изгибается дугой и прикрепляется к грунту. Для прорастания семян водяного ореха нужен период покоя 4-6 месяцев, а после этого достаточно высокая температура воды (не ниже +10, + 12 °С).
Рис. 116. Водяной орех плавающий (Trapa natans): 1 - общий вид растения; 2 - цветок; 3 - начало образование плода; 4 - плод
Механизм опыления цветков, которые раскрываются утром и только на несколько часов, не совсем ясен. Согласно большинству наблюдений у водяного ореха преобладает самоопыление, причем иногда цветки его даже не раскрываются. В то же время в литературе встречаются отдельные указания на возможность опыления цветков водяного ореха какими-то насекомыми, в частности перепончатокрылыми (шмели, осы, пчелы), в изобилии летающими над ним рано утром. Зато зрелые плоды его могут сохраняться в иле, не теряя всхожести, до 10 лет, а по Гамсу (1925) даже до 45-50 лет.
Водяной орех был известен человеку с глубокой древности из-за своих съедобных вкусных орехов, ядро которых содержит до 50% крахмала.
Фракийцы умели изготовлять из него хлеб. В средние века его широко употребляли в пищу в Южной Франции, Италии, Хорватии и в других европейских и восточных странах. С давних пор и до наших дней его собирают и даже культивируют в Китае, Японии и особенно в Индии, где он не раз спасал население от голода. В большом количестве собирают чилим и в Центральной Африке, где есть даже озеро Ньяса, что означает "жилище водяного ореха". Собирают водяной орех с лодки, получая до 600 кг чистого ядра с 1 га. Его едят в печеном и вареном виде, причем по вкусу он напоминает вареные каштаны, откуда и произошло одно из его названий - водяной каштан. Просушенное и измельченное ядро может идти на крупу (типа манной) и на муку, из которой с добавлением пшеничной получают неплохой хлеб.
Водяные орехи - не только пищевой продукт. Это также ценное лекарственное и красильное сырье. Древние народы употребляли их орехи от различных заболеваний, для украшения одежды и как талисманы.
В нашей стране во многих местах (Белоруссия, центр Русской равнины, Башкирия) проводят опыты по разведению водяного ореха.
Семейство сланоягодниковые (Haloragaceae) (А. П. Белавская)
Семейство сланоягодниковых включает 6 родов и около 130 видов, распространенных по всему земному шару, но в основном в южном полушарии. В СССР представлен единственный род семейства уруть (Myriophyllum).
Сланоягодниковые - водные, болотные, иногда наземные травы, отдельные представители которых приближаются к полукустарничкам. Листья очередные, супротивные или мутовчатые, разной формы и размеров. Цветки мелкие, анемофильные, собранные в верхушечные соцветия или сидячие в пазухах листьев, однополые или реже обоеполые, как правило, 4-членные, иногда 2- или 3-членные. Лепестки часто значительно крупнее чашелистиков, обычно опадающие, иногда отсутствуют. Тычинок обычно 8 в 2 кругах, реже тычинок 4 или 3, обычно с короткими нитями и относительно крупными пыльниками. Гинецей из 4, 3 или 2 плодолистиков, со свободными столбиками; завязь нижняя, 2-4-гнездная, с 1 висячим семязачатком в каждом гнезде. Плоды мелкие, ореховидные или костянковидные.
Наиболее крупный род семейства - сланоягодник (Haloragis) - включает около 75 видов, произрастающих в Южной, Юго-Восточной и Восточной Азии (на севере до острова Хоккайдо), в умеренных областях Северной и Южной Америки, в Австралии и Тасмании, Новой Зеландии, на островах Тихого океана, а 1 вид встречается на Мадагаскаре. Большинство видов произрастают в Австралии, Тасмании и Новой Зеландии. У некоторых видов сланоягодник древеснеют основания побегов, что сближает их с полукустарничками. Такая же особенность отмечена и у видов лауренбергии (Laurenbergia) - тропических трав южного полушария. Переходной ступенью к водным представителям семейства можно считать мезиеллу (Meziella) - мелкое травянистое растение Австралии. К настоящим
водным растениям относятся уруть - род, включающий около 40 видов, и два мелких рода (1-4 вида) - прозерпинака (Proserpinaca) из Северной Америки и лаудония (Loudonia) из Австралии.
Название Myriophyllum происходит от двух слов: miros - бесчисленный и phyllum - листок. И действительно, длинные (до 1,5 м ) стебли урути с глубоко рассеченными на нитевидные доли листьями (диаметр 220-380 мкм), собранные в мутовки, образуют в воде густое кружево подводных зарослей. С первого взгляда уруть похожа на роголистник (Ceratophyllum), но при более внимательном рассмотрении видно, что перистые листья урути хорошо отличаются от вильчато разветвленных и более ломких листьев роголистника. Мелкие розоватые или беловатые цветки урути собраны в редкий колос, возвышающийся над водой. Интересно, что в одном колосе могут быть цветки трех типов: обоеполые, мужские и женские. Опыляются цветки урути ветром, хотя отмечены и случаи энтомофилии. Ветроопылению способствуют тонкие нити пыльников, приходящие в движение от малейшего ветерка. Самоопыление встречается как исключение. Иногда бывают заросли урути и с полностью стерильными цветками. Плоды созревают в середине сентября, а еще через месяц начинают опадать, что растягивается на несколько месяцев. Вегетативное размножение осуществляется турионами (зимующими почками), а также любым отрезком побега. Между половым и вегетативным размножением существует явная корреляция - чем слабее развито плодоношение, тем интенсивнее образуются турионы.
Виды урути живут в стоячих или медленно текучих пресных водах и, как исключение, на морских побережьях. Из пяти видов урути, встречающихся в СССР, наиболее широко распространена и наиболее обильна уруть колосистая (M. spicatum, см. рис. 236, с. 448). Заросли ее приурочены обычно к глубинам от 0,3 до 2 м, к илистым грунтам и к водам, богатым кальцием. При высоком содержании в воде кальция листья урути часто покрываются известковой корочкой. Уруть колосистая очень чувствительна к температуре воды и менее - к освещенности.
Подводные луга из урути играют очень важную роль в жизни водоема. В ее зарослях отмечаются большие скопления мелких беспозвоночных животных, которые являются пищей для многих обитателей водоема. Сама же уруть служит кормом для растительноядных рыб и птиц (семена), а также субстратом для метания икры рыбами и убежищем для всего животного населения водоема, особенно для мальков рыб.
Семейство ризофоровые (Rhizophoraceae) (С. С. Морщихина)
Слово "ризофоровые" невольно напоминает о мангрове. Действительно, из всего семейства, охватывающего около 170 видов, относящихся к 17 родам, наиболее известны и изучены представители 4 родов: ризофора (Rhizophora), бругиера (Bruguiera), цериопс (Ceriops) и канделия (Kandelia), образующие основную массу мангровых зарослей. Мангровы окаймляют побережья тропических морей в зоне приливов и отливов, в бухтах, в эстуариях рек, где происходит отложение ила и песка. Неудивительно, что необычные деревья-амфибии, регулярно затопляемые соленой морской водой, всегда привлекали внимание исследователей.
Ризофоровые, как правило, небольшие деревья или кустарники. Некоторые из них достигают в высоту 30-40 м, например бругиера голокорневая (B. gymnorhiza, рис. 117, табл. 33), пеллакаликс Лобба (Pellacalyx lobbii). Многие виды имеют тенденцию к образованию придаточных корней в нижней части ствола, иногда образуются досковидные корни, придающие устойчивость высоким деревьям. Кора стволов, ветвей и придаточных корней снабжена чечевичками, межклеточные пространства которых связаны с межклетниками внутренних тканей, и через них, как и через устьица, осуществляется газообмен.

Таблица 33. Комбретовые, кипрейные и ризофоровые: 4 - бругиера голокорневая (Bruguiera gymnorhiza), оранжереи Ботанического института АН СССР в Ленинграде
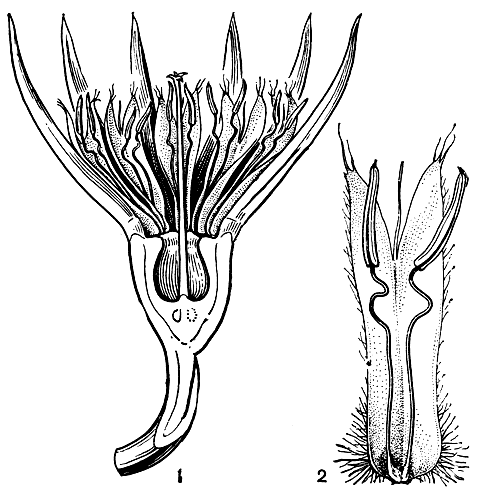
Рис. 117. Бругиера голокорневая (Bruguiera gymnorhiza): 1 - цветок в разрезе; 2 - развернувшийся лепесток, освободивший тычинки
Ветви в узлах у ризофоровых обычно вздуты, у кроссостилиса (Crossostylis), гинотрохеса (Gynotroches) и пеллакаликса - внутри полые. Листья простые, цельные, нередко кожистые, большей частью супротивные, с перистым жилкованием. У видов анизофиллеи (Anysophyllea) и других близких к ней родов листья очередные, нередко с дугонервным жилкованием, без прилистников, иногда чередующиеся с более мелкими чешуевидными листьями.
Для ризофоровых характерен моноподиальный тип ветвления. Межчерешковые крупные прилистники защищают верхушечную точку роста и опадают при разворачивании новой пары листьев, которые нередко появляются вместе с заложенными в их пазухах бутонами и боковыми побегами. Многие ризофоровые имеют тенденцию к непрерывному росту, и годичных колец в их древесине нет. Анатомическое строение древесины у видов, обитающих на суше, значительно отличается от древесины мангровых.
Цветки ризофоровых преимущественно в небольших верхоцветных соцветиях (табл. 33), иногда одиночные в пазухах листьев, актиноморфные, 4-5-членные, обычно обоеполые. Иногда встречаются виды с однополыми цветками или даже двудомные растения (анизофиллея). Цветоложе часто бокаловидное или блюдцевидное, вокруг гинецея с кольцевидным диском, который снабжен желёзками, выделяющими нектар. Чашечка неопадающая, из 3- 16 мясистых или кожистых створчатых долей, сросшихся у основания. Лепестки свободные, в числе, равном чашелистикам, края их нередко изрезанные, бахромчатые или реснитчатые; лепестки неяркие, чаще белые, желтоватые или коричневатые. Тычинок обычно в 2 (иногда в 3-4) раза больше, чем лепестков, расположены они по краю околопестичного диска, обычно в один круг. Завязь, как правило, нижняя, реже полунижняя, у видов гинотрохеса и кассипуреи (Cassipourea) верхняя, образована 2 или 4, редко 6 плодолистиками, с гнездами соответственно их числу; иногда завязь одногнездная в результате деградации перегородок (у видов канделии). Каждое гнездо содержит 2-4 семязачатка, прикрепленных в центре завязи. Столбик обычно простой, рыльце разделено на 2 или 4 лопасти в зависимости от числа плодолистиков. У видов анизофиллеи и близких к ней родов плодолистики срастаются только в нижней части, образуя 3-4 столбика с простыми рыльцами.
Цветение и опыление лесных видов ризофоровых мало изучено, известны лишь наблюдения над мангровыми видами. Исследования П. Б. Томлинсона, Р. Б. Примака и Дж. С. Банта (1979) показали, что цветки мангровых ризофоровых протандричны и опыляются преимущественно ветром, хотя их посещают и насекомые. Пыльники открываются еще в бутоне, но самоопыления не происходит, так как рыльце созревает лишь на второй день. У видов бругиеры и цериопса существует особый механизм выстреливания пыльников, который подробно наблюдал на Яве немецкий ботаник Карл Германн в 1911 г. Он нашел, что обильно выделяющие нектар неяркие цветки бругиеры активно посещают птицы-нектарницы и многочисленные насекомые, в том числе бабочки из семейства бражников (Sphingidae), имеющие длинный хоботок. В только что раскрытом цветке сложенные продольно лепестки держат в плену тычинки, нити которых пружинообразно извиты. Края лепестков покрыты щетинистыми жесткими волосками, образующими особенно густую зону в самом основании лепестка. Прикосновение к этой зоне вызывает мгновенное разворачивание лепестков, тычиночные нити распрямляются, пыльники вскрываются обычно продольной щелью, выстреливая облачко пыльцы. Опустошенные пыльники вскоре опадают вместе с лепестками, и тогда начинает функционировать рыльце, выделяющее каплю сахаристого нектара.
Плоды ризофоровых всегда несут чашелистики. Преобладает плод-ягода, обычно не очень сочная, или коробочка, раскрывающаяся простой трещиной. У многих представителей плоды односемянные. Анизофиллея имеет костянковидные плоды, у мангровых ризофоровых - жесткие деревянистые односемянные плоды.
Семена у большинства ризофоровых с эндоспермом. Зародыш, содержащий хлорофилл, прямой, обычно с 2 семядолями, у некоторых видов бругиеры 3 или 4 семядоли. Семена видов кроссостилис и кассипурея имеют ариллус. Семена макаризии (Macarisia) снабжены крыловидным выростом.
Наземные ризофоровые в своем большинстве являются элементом подлеска и нижних ярусов влажного тропического леса. Лишь немногие достигают его полога: пеллакаликс Лобба, гинотрохес пазушный (Gynotroches axillaris) и караллия раскидистая (Carallia brachiata). Более светолюбивые виды являются пионерами при зарастании вырубок, встречаясь массово во вторичных формациях. Сплошные заросли образует на низинных торфяных болотах островов Сулавеси и Калимантан комбретокарпус округлый (Combretocarpus rotundatus).

Таблица 33. Комбретовые, кипрейные и ризофоровые: 5 - караллия ланцетолистная (Carallia lancifolia), оранжереи Ботанического института АН СССР в Ленинграде
Большинство ризофоровых приурочено к постоянно влажному климату, лишь немногие входят в состав муссонных лесов и формаций типа саванн. На склонах гор ризофоровые сравнительно редко переходят границу тропической зоны, т. е. 1000 м над уровнем моря. Рекорд высотного распространения принадлежит роду гинотрохес, который встречается в Юго-Восточной Азии, в том числе на островах Малайского архипелага, от низинных болотистых лесов до 2250 м на склонах гор.
Представители ризофоровых, обитающие в мангровах, являются обычно главным ее компонентом, определяющим облик этих зарослей. Мангровы обычно состоят из вечнозеленых деревьев высотой 10-15 м. Наиболее высокоствольные виды (до 27-30 м) встречаются в мангровах восточного полушария: бругиера голокорневая, б. шестиугольная (B. sexangula) и ризофора остроконечная (R. mucronata). Виды бругиеры образуют на более сухих, внутренних участках мангров исключительно густые прямоствольные стояния, под пологом которых постоянно царит полумрак. Виды ризофоры, как правило, пионерные, растущие с мористой сторон мангровы и принимающие на себя шквалы ветра и удары волн. Во время прилива морская вода затопляет их нередко до самых крон, а отлив обнажает стволы и густое сплетение корней. Ризофоровые, обитающие в мангровах, обладают способностью образовывать придаточные опорные, так называемые ходульные корни на нижней части ствола. У ризофоры такие корни появляются и на нижних ветвях, достигая почвы; они обычно ветвятся и придают деревьям устойчивость против штормов. Корневая система мангровых ризофоровых неглубокая, распростертая в верхних слоях грунта. У бругиеры радиально отходящие от основания ствола горизонтальные корни образуют высокие вертикальные узловатые и корявые коленчатые выросты, торчащие из грунта. Нижняя их часть, погруженная в почву, несет многочисленные питающие корни, верхняя с годами нарастает в высоту и покрыта пробкообразной коркой, через поры которой происходит снабжение кислородом тканей корневой системы.
Во время отлива грунт значительно подсыхает и содержание соли в нем возрастает в несколько раз. Деревья мангров обладают удивительной способностью выносить большие колебания концентрации солей (главным образом поваренной соли) в почве. Корни их обеспечивают всасывание опресненной воды путем ультрафильтрации, как показали исследования П. Ф. Сколандера, проведенные в 1968 г. Жидкость, поступающая в сосуды древесины мангровых, содержит всего около 0,03% соли. И все же соль накапливается в тканях, особенно сильно в старых листьях за счет длительной транспирации.
В необычных экологических условиях обитания сложилось особое свойство мангровых ризофоровых - живорождение, вивипария, т. е. прорастание семени в плоде, находящемся на материнском растении (рис. 118). Крупный вытянутый зародыш развивается непрерывно, без периода покоя и через 11-13 недель после оплодотворения пробивает стенку плода и продолжает энергично расти. Зеленые веретеновидные проростки в изобилии висят на ветвях деревьев, у ризофоры остроконечной они достигают в длину 1 м.
Интересное исследование строения и условий питания зародыша ризофоровых опубликовала в 1940 г. А. Кипп-Голлер. Семядоли, сросшиеся в единое тело, окружающее точку роста, тесно срастаются с внутренним интегументом, образуя плацентарный орган, через который осуществляется питание растущего зародыша. Здесь на пути через ткани интегумента и семядолей происходит очень важный процесс опреснения воды и снижения осмотического давления растворов с перепадом на 1300-1700 кПа. Благодаря этому процессу дерево, растущее на засоленном грунте и накапливающее в своих тканях поваренную соль, обеспечивает развивающиеся на нем проростки почти пресной водой. Проростки остаются на дереве 30-39 недель, иногда целый год, затем падают под влиянием силы тяжести, втыкаясь почти вертикально в мягкий ил. На более плотном грунте опавшие проростки остаются лежать на поверхности и укореняются лежа, постепенно поднимаясь. Нередко при этом часть их подвергается высушиванию на солнце. В эксперименте, проведенном в 1954 г. К. Д. Ла Рю и Т. Дж. Мьюзиком, проростки ризофоры мангле (R. mangle, рис. 118), лежавшие 68 дней на лабораторном столе и потерявшие одну треть массы, в течение двух дней восстановили потерю воды при посадке во влажный песок и через две недели укоренились. В такой же срок произошло укоренение проростков ризофоры мангле в ботаническом саду Ботанического института АН СССР в Ленинграде, когда были посажены в ил проростки, находившиеся 52 дня в гербарии.

Таблица 33. Комбретовые, кипрейные и ризофоровые: 6 - участок мангровых зарослей на острове Куба из ризофоры мангле (Rhizophora mahgle)
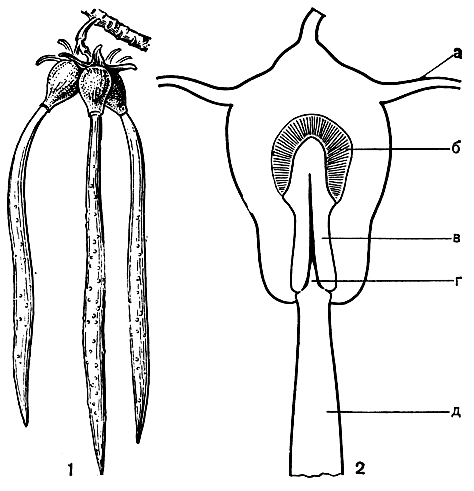
Рис. 118. Ризофора мангле (Rhizophora mangle): 1 - плоды с проростками на ветке; 2 - схема продольного разреза проросшего плода (а - чашелистик, б - ткань интегумента, в - семядольное тело, г - плюмула проростка, д - гипокотиль проростка)
Унесенные морской волной, проростки ризофоровых могут совершать длительные (до года) морские путешествия, сохраняя при этом жизнеспособность. Это является основным фактором, обеспечивающим широкое распространение ризофоровых на тропических морских побережьях. Местами они переходят линию тропиков, где теплые течения и климат благоприятствуют развитию мангровы. Наиболее северные точки распространения мангров - около 32° с. ш. во Флориде и на Бермудских островах, в Красном море (залив Акаба) и на юге Японии. В южном полушарии мангровы отходят еще дальше от экватора. На восточном побережье Африки они достигают окрестностей Дурбана (33° ю. ш.), а в Восточной Австралии -38° ю. ш. Наименьшую протяженность ареал мангровы имеет на Тихоокеанском побережье Америки, от Нижней Калифорнии до 3°48' ю. ш. в Южной Америке, где ее распространение ограничивает холодное течение Гумбольдта. В Атлантическом океане мангровы достигают 28°2(У на юге Бразилии, на побережье Африки они приурочены главным образом к эстуариям крупных рек, и распространение их к югу останавливает около 9° ю. ш. сухой, почти пустынный климат юга континента.
Экономическое значение ризофоровых невелико. Древесина некоторых видов, главным образом мангровых, служит топливом, используется в строительстве, в основном для свай в подводных и подземных сооружениях. Кора мангровых ризофоровых, содержащая до 40% дубильных веществ, находит применение в кожевенном производстве и для получения красителей. Пога маслянистая (Poga oleosa) из тропических лесов Западной Африки содержит в семенах около 60% пищевого масла, используемого местным населением.
Семейство лецитисовые (Lecythidaceae) (С. С. Морщихина)
Семейство лецитисовых охватывает 24 рода и около 450 видов, распространенных в тропиках обоих полушарий.
Среди лецитисовых нередки мощные, 30-40-метровые деревья, входящие в верхний ярус влажных тропических лесов. Иногда их колонновидные стволы диаметром около метра снабжены у земли опорными досковидными корнями и несут ветви лишь на высоте 15-18 м. Таких гигантов особенно много среди видов рода лецитис (Lecythis) и эшвейлера (Eschweilera) из тропических лесов Южной Америки. Немало в семействе и низкорослых деревьев из нижних ярусов леса высотой всего 2-5 м. Лишь совсем немногие виды представлены невысокими кустарниками. В Бразилии, в штате Мату-Гросу встречается кустарник эшвейлера карликовая (E. nana) высотой всего 30-40 см.
Для лецитисовых характерны очередные цельные листья, сконцентрированные на концах ветвей. У некоторых видов густавии (Gustavia) длина листьев больше 80 см. Как у многих тропических деревьев, молодые листья лецитисовых имеют розоватую окраску и нежную, повисающую пластинку.
Цветки лецитисовых обычно крупные, диаметром до 5-20 см, яркие и душистые, расположены одиночно в пазухах листьев или в бокоцветных соцветиях. У некоторых видов баррингтонии (Barringtonia), курупиты (Couroupita) и других представителей семейства наблюдается явление каулифлории.
Околоцветник в нижней части образует трубку, полностью приросшую к завязи. Чашечка обычно 4-6-лопастная, редко из 2-3 долей, иногда доли чашечки полностью срастаются и рвутся при раскрывании цветка на 2-5 неправильных частей, такая чашечка у некоторых видов баррингтонии. Лепестки обычно крупные, мясистые, белые, кремовые, розовые или красные, тоже, как правило, в числе 4-6; у некоторых лецитисовых цветки лишены лепестков. Число тычинок колеблется: у куратари продолговатолистной (Couratari oblongifolia) их всего 10, а у лецитиса Пуато (Lecythis poiteau) и некоторых видов густавии (Gustavia) больше тысячи. Тычинки располагаются в нескольких кругах, число которых варьирует от 3 до 8 и является систематическим признаком, как и все строение андроцея. Тычиночные нити у лецитисовых срастаются в основании в трубчатый или чашевидный андрофор. Наружный или внутренний круг может быть образован стерильными тычинками-стаминодиями. У видов африканского рода наполеона (Napoleona) актиноморфные цветки лишены лепестков, а три наружных круга тычинок полностью стерильны и образуют ярко окрашенную венчиковидную структуру (рис. 119). В наружном круге сросшиеся стаминодии напоминают лучевидно плиссированный воротник, второй круг - из более или менее свободных линейных стерильных тычинок и третий состоит из 20-60 закрученных внутрь стаминодиев, имеющих у основания шпорцевидные выросты. Только четвертый, внутренний круг содержит фертильные тычинки, чередующиеся со стаминодиями.
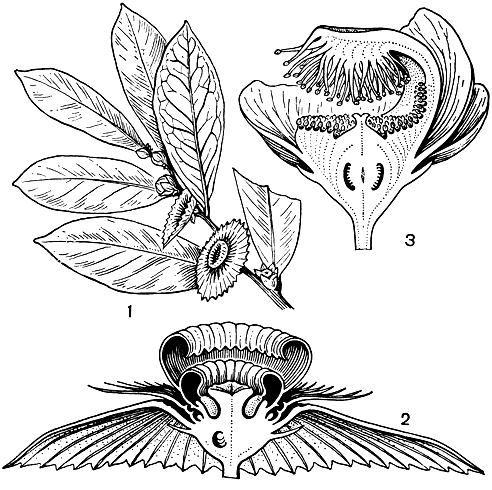
Рис. 119. Лецитисовые. Наполеона остроконечная (Napoleona cuspidata): 1 - ветвь с цветками; 2 - продольный разрез цветка. Курупита гвианская (Couroupita guineensis): 3 - продольный разрез цветка
У южноамериканских лецитисовых прослеживается переход от актиноморфных цветков густавии и гриаса (Grias) к зигоморфному строению андрофора в цветках лецитиса, курупиты и других родов. Наружное кольцо их сросшихся тычинок имеет с одной стороны вытянутый лентовидный вырост, придаток, нередко сложно спирально закрученный или в виде капюшона нависающий над центром цветка (см. рис. 119). Придаток может нести стаминодии, выделяющие нектар, или фертильные пыльники. Он обычно ярко окрашен; строение его чрезвычайно разнообразно.
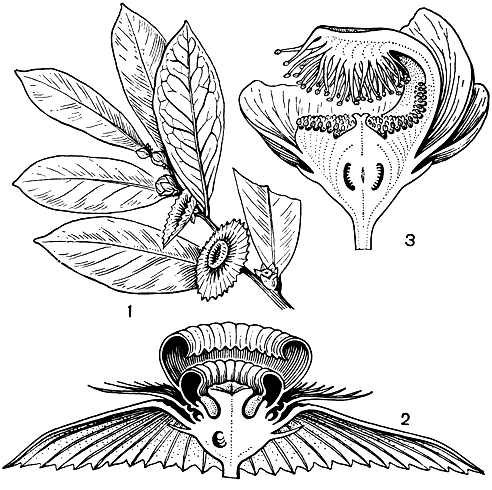
Рис. 119. Лецитисовые. Наполеона остроконечная (Napoleona cuspidata): 1 - ветвь с цветками; 2 - продольный разрез цветка. Курупита гвианская (Couroupita guineensis): 3 - продольный разрез цветка
Для лецитисовых характерно наличие в цветке нектарного диска, выпуклым валиком окаймляющего столбик завязи. Гинецей из 2-6 плодолистиков, завязь преимущественно нижняя, соответственно числу плодолистиков 2-6-гнездная. Семязачатки анатропные, обычно многочисленные, редко по 4-2 или по одному в гнезде, расположены вертикально, в центральном углу гнезда. Столбик простой, с головчатым или лопастным рыльцем.
По наблюдениям американских биологов С. А. Мори, Г. Т. Пранса и А. Б. Болтена (1978), проведенным в лесах Южной Америки, для лецитисовых I и II яруса дождевого леса характерно массовое кратковременное цветение, а для деревьев подлеска - растянутое до 5-6 месяцев. Обычно раскрывается одновременно по одному цветку в соцветии, каждый цветок функционирует чаще всего в течение одного светового дня, вечером лепестки и андроцей уже опадают.
Для лецитисовых характерно перекрестное опыление насекомыми, преимущественно пчелами На цветках густавии великолепной (Custavia superba) было найдено 11 видов из 5 различных родов насекомых. Лецитисовые, имеющие цветок с асимметричным андрофором, более специализированы в отношении опылителей. Их опыляют крупные пчелы из рода ксилокопа (Xylocopa), африканская раса медоносной пчелы (Apis mellifera) и шмели (Bombidae), посещающие цветки с рассвета до 11 ч утра. Днем выделение нектара прекращается, пыльники вянут Пчелы садятся на придаток андрофора и залезают под него, поворачиваясь вниз спинкой, которая при этом трется о фертильные тычинки и уносит с них пыльцу. В окрестностях Манауса в Бразилии Прайсом были найдены на цветках лецитиса, эшвейлеры и голопиксидиума (Holopyxidium) жуки Cyclocephala verticalis из семейства пластинчатоусых (Scarabaeidae), которые опыляют также цветки виктории амазонской и других нимфейных.
Некоторые лецитисовые, например баррингтония, хиденантус (Chydenanthus), цветут ночью и опыляются птицами-нектарницами и летучими мышами. Лецитис Пуато, растущий в лесах долины Амазонки, раскрывает свои цветки в сумерки. Всю ночь дерево гудит и сотрясается от сотен летучих мышей, привлекаемых обильным нектаром. Пик посещения цветков - с 7 до 9 вечера, в 3 ч ночи начинается опадание лепестков и андрофоров. За ночь с дерева опадает около пятисот цветков. Лакомятся нектаром лецитисовых обезьяны и опоссумы, очевидно, не участвующие в опылении, так же как и попугаи, поедающие в огромном количестве сочные андрофоры цветков.
Плоды лецитисовых созревают долго: так, у бертоллетии высокой (Bertholletia excelsa) в течение 13-15 месяцев, у лецитиса обыкновенного (Lecythis usittata) - 18-22 месяца. Строение и форма плодов у лецитисовых разнообразны: встречаются костянковидные, ягодообразные плоды или коробочки. Для южноамериканских представителей семейства характерен плод-крыночка, т. е. деревянистая коробочка, открывающаяся крышечкой по внутреннему краю околопестичного диска, который хорошо заметен на плоде в виде выпуклого валика (рис. 120). Крыночка имеет обычно округлую или овальную, а иногда более или менее вытянутую форму. Круглые плоды лецитиса называют в Бразилии и Гвиане "обезьяньим горшком", так как ими пользовались для ловли обезьян, закладывая в пустую оболочку плода приманку.

Рис. 120. Плод лецитиса эллиптического (Lecythis elliptica)
По мере созревания плодов перегородки завязи, как правило, разрушаются и остается лишь центральная колонка, несущая семена. У видов аллантомы (Allantoma) и каринианы (Cariniana) колонка прирастает к крышечке и отделяется вместе с ней, вынося семена из плода. У видов лецитиса крышечка отделяется тоже вместе с колонкой, а семена остаются в плоде и выпадают на землю позже, с разлагающимся слизистым, почти жидким мезокарпом.
Наиболее крупные плоды, диаметром до 20 см, имеют некоторые виды курупиты. Курупита гвианская (Couroupita guineensis), или "пушечное дерево", несет на стволе большое количество тяжелых плодов, постукивающих на ветру.
Крупные односемянные плоды характерны для видов баррингтонии, обитающих главным образом по берегам рек и морей и связанных с водой в распространении плодов. Мезокарпий у них представляет легкую губчатую ткань, пронизанную волокнами, благодаря чему плоды обладают хорошей плавучестью. Четырех-или пятигранные плоды баррингтонии азиатской (B. asiatica) часто находят на пляжах в морских выбросах, где они и прорастают.
Семена лецитисовых крупные, лишены эндосперма и полностью заняты зародышем. У баррингтонии зародыш не дифференцирован, как обычно, на гипокотиль и семядоли. Он состоит из крупного осевого органа, несущего в верхушечной части спирально расположенные чешуи. При прорастании сеянец развивает ряд чешуевидных мелких листьев, катафиллов, затем появляется несколько листьев с развитой пластинкой, и снова следует серия катафиллов (рис. 121). Подобное строение зародыша имеют семена карейи (Careya). У бертоллетии зародыш имеет две рудиментарные семядоли.

Рис. 121. Сеянец баррингтонии азиатской (Barringtonia asiatica)
Как у многих тропических растений, семена лецитисовых быстро теряют всхожесть, однако в ботаническом саду АН СССР в Ленинграде удалось получить хорошие всходы густавии великолепной на 21-й день после посева, произведенного 25 дней спустя после сбора плодов. Семена бертоллетии, имеющие твердую оболочку, всходят обычно через 2-3 месяца после посева.
Современное распространение лецитисовых ограничено 30° ю. ш. и 35° с. ш. Лецитисовые произрастают главным образом на небольших высотах над уровнем моря, лишь немногие виды встречаются в горах. Так, баррингтония остроконечная (B. apiculata) поднимается до 1000-1500 м на Новой Гвинее в светлых горных лесах. Баррингтония азиатская и баррингтония кистевидная (B. racemosa) образуют леса на побережьях Индийского и западной части Тихого океана и имеют широкое распространение благодаря плавающим плодам.
В южноамериканских дождевых лесах лецитисовые занимают второе место после бобовых по частоте встречаемости и нередко образуют чистые насаждения. Естественные насаждения бертоллетии высокой, распространенной в лесах Гвианы и Бразилии от устья Амазонки до среднего течения Риу-Негру, эксплуатируют с целью сбора плодов. Плоды бертоллетии заключают 15-25 трехгранных крупных семян, содержащих до 75% пищевого масла. Семена известны под названием "бразильского ореха", высоко ценятся и используются в кондитерской промышленности. Плантации бертоллетии в Бразилии, Индии, Индонезии и на Филиппинских островах сравнительно невелики и не имеют большого экономического значения. Пищевое и техническое масло, используемое в мыловарении, получают также из семян некоторых видов лецитиса. Древесина многих, особенно высокоствольных видов ценится как строительный материал.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'