Порядок вересковые (Ericales) (Ю. П. Кожевников)
Семейство актинидиевые (Actinidiaceae)
В семействе 3 рода и около 350 видов, распространенных в странах с теплым влажным климатом. Актинидиевые представлены деревьями, кустарниками и лианами с относительно крупными очередными листьями. Цветки у них в пазушных верхоцветных соцветиях, обоеполые или реже однополые, 5-членные, с 10 или многими тычинками. Пыльники вскрываются короткими щелями или верхушечными порами. Гинецей из 3-5 или реже многих плодолистиков; столбики свободные или реже более или менее сросшиеся; завязь верхняя, обычно с многочисленными семязачатками в каждом гнезде. Плод - ягода или реже коробочка. Семена мелкие.
Род заурауйя (Saurauia) насчитывает около 300 видов невысоких деревьев и кустарников, распространенных в горных областях тропической и отчасти субтропической Америки, Гималаев и Юго-Восточной Азии, с одним видом в Северо-Восточном Квинсленде (Австралия). Заурауйя имеет простые кожистые листья длиной до 42 см и шириной 14 см, обычно собранные пучками на концах ветвей. Для многих кустарниковых заурауй характерны соцветия из большого числа (до 150) мелких цветков, находящихся на концах ветвей. У древесных видов цветков меньше, но их диаметр может достигать 2-2,6 см (рис. 41, 1). Нектарники в цветках в большинстве случаев отсутствуют, однако у некоторых видов обнаружена нектароносная ткань, что свидетельствует об опылении их насекомыми. Характерна протандрия. Плоды заурауйи представляют круглые ягоды с большим количеством мелких семян.
Род актинидия (Actinidia) насчитывает около 40 видов, распространенных в Гималаях и в Восточной и Юго-Восточной Азии. Некоторые виды достигают на севере Сахалина, Приморья и южных Курильских островов. Актинидии - это деревянистые лианы с опадающими листьями, часто с оттянутым кончиком (рис. 41, 2). Цветки разной величины (диаметром 1-1,5 см у актинидии коломикта - A. kolomikta и 3- 4 см у актинидии китайской - A. chinensis, табл. 11), собраны по 3 в пазухах листьев или одиночные, большей частью белые, но бывают золотисто-желтые или оранжевые, чашевидной формы. Гинецей обычно из многих плодолистиков и на крупной округлой завязи лучевидно расходится большое количество рылец. Эта особенность отражена в названии растения (греч. aktis, aktinos - луч, eidos - вид). У одних видов цветки не имеют запаха, у других они сильно ароматичны, например актинидия многодомная (A. polygama) и актинидия коломикта. Чашелистиков и лепестков по 5, но иногда их число от 3 до 6.
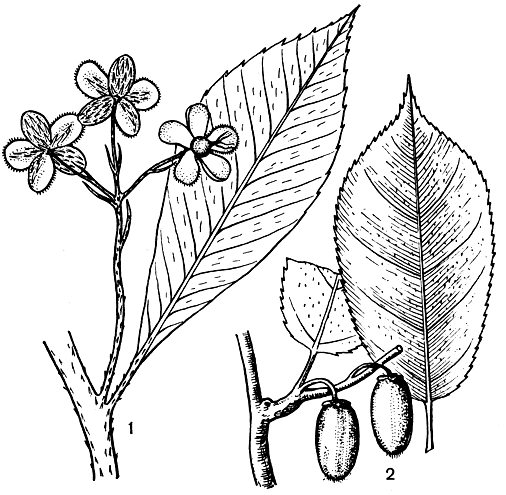
Рис. 41. Актинидиевые. Заурауйя чешуеплодная (Saurauia squamifructa): 1 - часть побега с чашелистиками цветков, на которых заметен различный характер опушения. При сомкнутых чашелистиках опушение бутона равномерное. Актинидия коломиктa (Actinidia kolomikta): 2 - часть побега с плодами
Хотя актинидии обычно строго двудомные растения, в женских цветках имеются и тычинки, которые, однако, не вырабатывают жизнеспособной пыльцы. В мужских же цветках гинецей лишен рылец, а завязь - семязачатков. Однополые цветки актинидии коломикта на ранних стадиях формируются как обоеполые. В опылении актинидий участвуют и ветер, и насекомые.
Плоды актинидий - продолговатые желто-зеленые или светло-оранжевые ягоды длиной до 3-4 см. У некоторых актинидий ягоды имеют кисловато-сладкий ананасный вкус и очень богаты витамином С. Однако не у всех видов они съедобны. У актинидии многодомной ягоды перчат так, что обжигают слизистую оболочку рта, поэтому местные жители так и называют эти ягоды "перцем". Из плодов актинидии китайской выделен протеолитический фермент актинидии, предотвращающий свертывание крови. В некоторых странах, например в Китае, этот вид широко культивируется из-за своеобразного вкуса его довольно крупных плодов. В Новой Зеландии существуют плантации актинидии китайской, плоды которой служат продуктом экспорта. В СССР эта актинидия введена в культуру в Батуми, Ташкенте и других южных городах; культивируется также актинидия острая (A. arguta) и очень морозостойкая актинидия коломикта, которая способна плодоносить в открытом грунте северных ботанических садов, например в Ленинграде. Очень приятны на вкус плоды коломикты, богатые витамином С. В Японии в вареном виде употребляют в пищу листья актинидии многодомной. В Юго-Восточной Азии едят молодые побеги актинидии мозолистой (A. callosa).
Как и большинство лиан, побеги актинидии растут очень быстро, до 5-7 м за лето. Наиболее крупная актинидия острая нередко достигает в длину 25 м. В Уссурийском крае был даже найден экземпляр длиной более 100 м. В дождливые годы актинидия острая иногда выпускает на концах побегов воздушные корни, благодаря которым при соприкосновении с землей или гнилыми пнями она может многократно укореняться. Размножается актинидия в основном корневой порослью. По мере роста лианы поворачиваются вокруг своей центральной оси и одновременно обвивают деревья или другую опору. Три вида актинидии на нашем Дальнем Востоке, произрастая совместно, да еще с амурским виноградом, иногда образуют труднопроходимые заросли.
Третий род семейства актинидиевых - клематоклетра (Clematoclethra) - насчитывает около 10 видов, которые распространены в горных лесах Западного и Центрального Китая. Это небольшие кустарники, побеги которых способны виться. Внешне клематоклетра сходна с актинидией, которая также имеет иногда кустарниковую форму роста. В цветках клематоклетры 10 тычинок и один длинный столбик с 5-раздельным рыльцем. Ее плод - суховатая ягода с 5 гнездами, в каждом из которых находится по одному семени.
Семейство клетровые (Clethraceae)
Семейство включает лишь один род клетра (Clethra), насчитывающий более 60 видов, распространенных в Восточной и Юго-Восточной Азии, в Центральной и тропической Южной Америке, с 1 видом на острове Мадейра (клетра древовидная - С. arborea) и 2 видами в юго-восточных штатах Северной Америки (карта 6). Виды клетры - небольшие кустарники или деревья, листопадные или реже вечнозеленые. Листья очередные. Цветки в верхушечных кистях или метелках (см. табл. 14, рис. 42), обоеполые, 5-членные. Тычинок 10-12, в 2 кругах. Пыльники стреловидные, вскрывающиеся верхушечными порами. Гинецей из 3 плодолистиков, с 3-лопастным на верхушке столбиком; завязь верхняя, 3-гнездная, с многочисленными семязачатками на интрузивных плацентах. Плод - локулицидная коробочка. Семена многочисленные, часто крылатые, распространяются ветром.
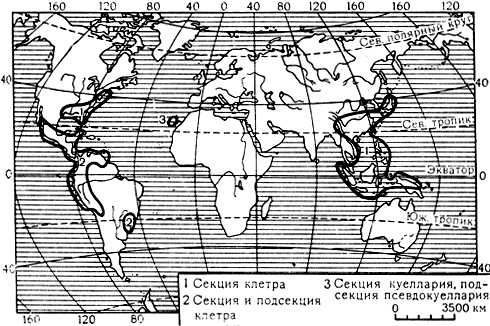
Карта 6. Ареал рода клетра
Цветки клетры опыляются насекомыми. Близ основания завязи у некоторых видов обнаружена нектароносная ткань, хотя нектарный диск у клетры отсутствует. Цветки многих видов сильно пахнут. Характерна протандрия, предотвращающая самоопыление. Так, у клетры олъхолиетной (C. alnifolia, рис. 42) в полностью раскрытом цветке с пылящими тычинками гинецей оказывается еще незрелым. Кроме того, пыльники при созревании разворачиваются на 180° и высыпают пыльцу в сторону, противоположную гинецею. Когда же вся пыльца высыпалась, столбик начинает быстро расти, а рыльца становятся клейкими; у других видов пыльца высыпается из пыльников еще в бутоне.

Рис. 42. Репродуктивные органы клетры. Клетра ольхолистная (Clethra alnifolia): 1 - цветок; 2 - раскрывшаяся коробочка, окруженная одревесневшим и чашелистиками. Клетра бородчато жилковатая (С. barbinervis): 3 - отдельный лепесток с тремя тычинками; 4 - завязь и столбик с тремя рыльцами
Семейство вересковые (Ericaceae)
Вересковые - крупное семейство; в нем насчитывается более 100 родов и свыше 3000 видов. Вересковые широко распространены по земному шару. Они не встречаются только в степях и пустынях, а в тропиках растут преимущественно в высокогорьях.
Большинство вересковых - кустарники или кустарнички, иногда травы, в том числе сапрофитные, но среди них имеются и крупные деревья. Так, оксидендрум (Oxydendrum) в Америке и эрика древовидная (Erica arborea) на Канарских островах достигают в высоту 20 м с диаметром ствола до 0,5 м. Еще одно крупное вересковое - рододендрон древовидный (Rhododendron arboreum) - растет в Гималаях. Один из американских видов рода пиерис (Pieris, табл. 13 - лиана, поднимающаяся вверх по стволам болотного кипариса (Taxodium) до 10 м. Но тот же пиерис растет и в виде куста. Среди кустарников и кустарничков из вересковых имеются стелющиеся формы. Такова всем известная клюква (Oxycoccus), отдельные рододендроны. Некоторые виды маклинии (Mackleania) и агапетеса (Agapetes, табл. 12) представляют собой кустарники с длинными цепляющимися побегами. Среди азиатских видов рододендрона, а также среди вакциниевых, в том числе маклинии, известны эпифиты. Растут они в тропических дождевых лесах, а также высоко в горах Юго-Восточной Азии, в так называемых мшистых лесах. К ним относятся рододендрон камелиецветный (R. camelliiflorum), рододендрон голубичный (R. vaccinioides), рододендрон деревцовый (R. dendricola) и др. Под верхним слоем эпидермы листья эпифитов имеют слой крупных тонкостенных клеток, служащих для запасания воды. Этот признак отсутствует у вересковых, ведущих наземный образ жизни. Семена у некоторых видов снабжены своеобразным придатком, похожим на летучки, что является приспособлением в данном случае не для переноса, а для зацепки за ветви деревьев, на которых они начинают развиваться. Среди эпифитных рододендронов преобладают растения со светлой, иногда чисто белой окраской цветков, но есть и такие, у которых цветки окрашены в грязные тона. Большая часть эпифитных вересковых может селиться также на скалах и гребнях гор, где они значительно меняют свой облик. Кроме того, среди вересковых имеется группа растений-полуэпифитов. Так, черника лавролистная (Vaccinium laurifolium) начинает свое развитие как эпифит, но когда ее свисающие корни достигают земли, она начинает расти как наземное растение и достигает в высоту 5 м, прислоняясь стволом к дереву, на котором начала свое развитие.
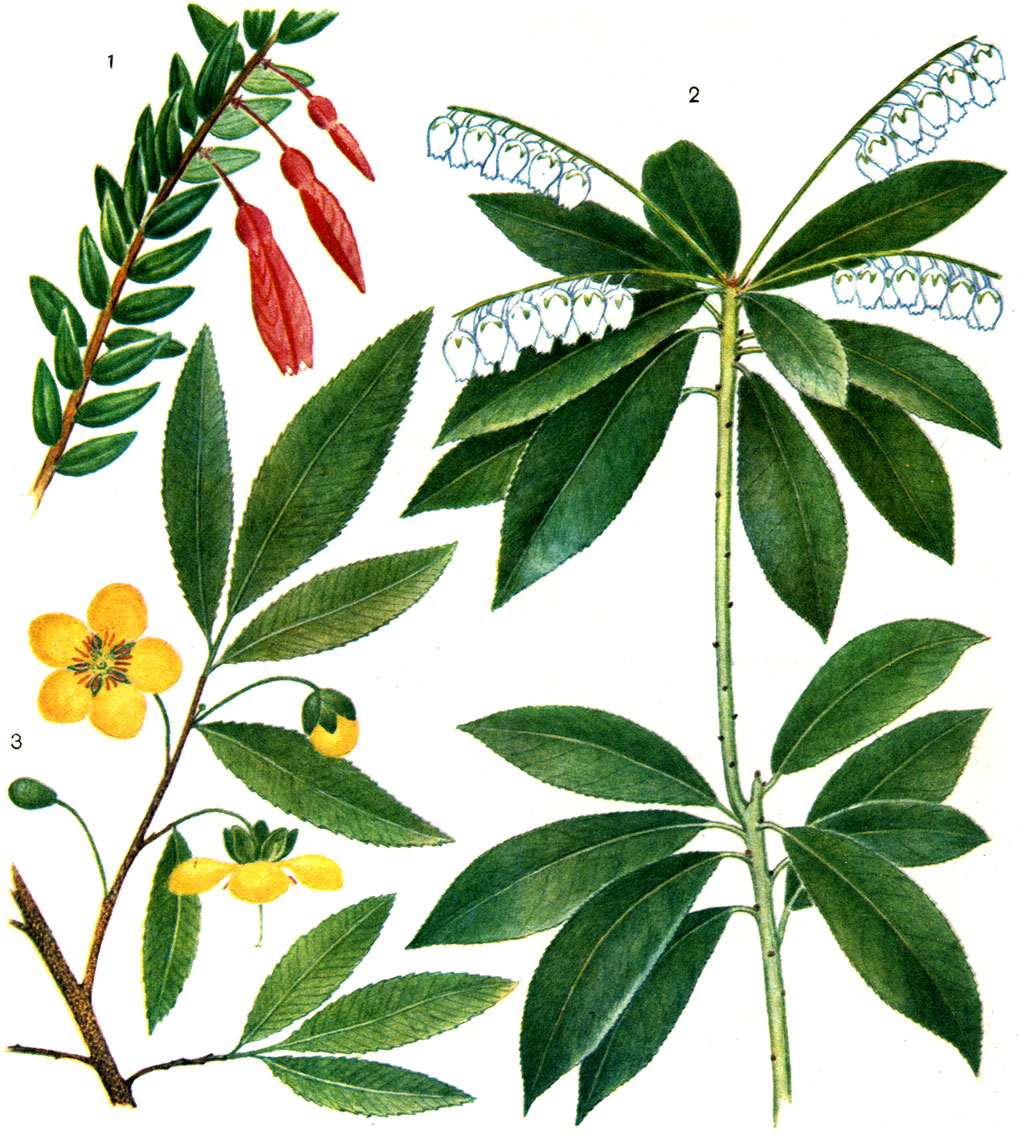
Таблица 12. Вересковые и охновые: 1 - агапетес ползучий (Agapetes serpens); 2 - кальмия многолистная (Kalmia polifolia); 3 - охна темно-пурпурная (Ochna atropurpurea)
Облик вересковых весьма своеобразен, поэтому в ботанике существует понятие "эрикоидный облик", который имеют и некоторые растения, не относящиеся к семейству вересковых. Эрикоидный облик создают деревянистые побеги, покрытые мелкими кожистыми листьями. Во многих случаях листья концентрируются на концах побегов, что хорошо видно у рододендронов. У одних видов листья совершенно голые и имеют глянцевитую поверхность, у других они по-разному опушены. Опушение развивается обычно на нижней поверхности листа, где у большинства вересковых находятся устьица. Потянув за кустик многих кустарничков из семейства вересковых - черники обыкновенной (Vaccinium myrtillus), подбела (Andromeda), брусники (Vaccinium vitisidaea), хамедафне болотной (Chamaedaphne calyculata), мы обычно видим, что от кустика отходит длинное корневище в виде шнура, которое вскоре подводит нас к следующему кустику, а далее - к следующему, и оказывается, что большая группа кустиков имеет общее корневище. У взрослой черники обыкновенной оно достигает 3 м. У других черник корневище тянется на 10 м. У многих вересковых (кустарничков и низких кустарников) в почве скрыт почти полностью стволик кустика, например у багульника (Ledum), толокнянки (Arctostaphylos). У третьих укореняются полегающие ветви, например у Кассиопеи (Cassiope), вереска (Calluna). Древесина у вересковых очень плотная и часто смолистая, напоминающая древесину хвойных. Смолы содержатся и в листьях, иногда выступая на поверхность.
Имеются вечнозеленые и листопадные виды вересковых. Среди разнообразия листьев вересковых выделяют три типа (рис. 43): 1) тип рододендрона (лист плоский или со слегка завернутыми вниз краями), такой лист имеют клюква, голубика и др.; 2) игловидный лист, напоминающий хвоинку, как у некоторых видов эрики (Erica), хариманеллы (Harrimanella); 3) эрикоидный лист с сильно завернутыми краями (у вереска). В некоторых родах, например у Кассиопеи, разные виды имеют различный тип листа. У некоторых вересковых листья вообще уникальны. Так, Кассиопея Редовского (C. redowskii) имеет лист, напоминающий палец перчатки, т. е. с почти замкнутой полостью. У многих вересковых лист может свертываться при наступлении засушливого периода; это отмечено, в частности, для брусники. Таким образом, у ряда вересковых существуют динамические реакции на изменения погодных условий. Чрезмерное испарение воды в сухую погоду ослабляет напряжение в слоях мезофила на нижней стороне листа, поэтому лист выгибается, скручивается; контакт листовой полости с окружающей средой уменьшается. Имеются, однако, сведения, что у очень многих вересковых, в частности у самого вереска, устьица есть и на верхней стороне листа. Они отсутствуют лишь на боковых его сторонах, покрытых мощной кутикулой. Таким образом, испарение происходит даже при полностью закрытой листовой полости. Значение этой полости становится понятным, если лист опустить в воду. При этом верхняя поверхность его смочится, тогда как в желобке на нижней поверхности останется пузырек воздуха, который не удалить и встряхиванием листа. Таким образом, устьица в листовой полости всегда остаются сухими и могут испарять влагу (транспирировать) даже под водой.
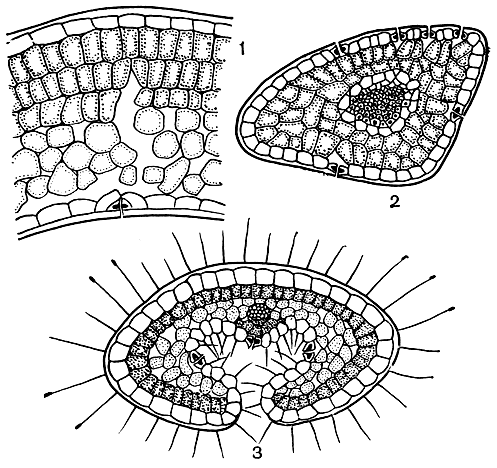
Рис. 43. Поперечные разрезы листьев вересковых: 1 - клюква (Oxycoccus); 2 - хариманелла (Harrimanella); 3 - эрика тетраликс (Erica tetralix)
Цветочные почки (бутоны) у некоторых видов очень крупные, напоминающие шишки хвойных. Большие почечные чешуи иногда ярко окрашены, например у тибодии (Thibaudia) из Южной Америки. Цветки самой разнообразной величины (рис. 44) собраны обычно в кистевидное соцветие на концах ветвей, реже соцветие бывает щитковидное (у багульника или рододендрона). Большинство вересковых имеет 5 чашелистиков, 5 лепестков, 10 тычинок, в завязи 5 гнезд, на рыльце можно рассмотреть 5 лучей. Однако четырехчленный план цветка считается вполне типичным для семейства (у вереска, брусники, клюквы, дабеции - Dabecia). Встречаются также двучленные (у симпиезы - Sympieza) и трехчленные цветки (у кладотамнуса - Cladothamnus), а у бежарии (Bejaria) они, как правило, семичленные. Характерной чертой цветка японских эллиотий (Elliotia) является то, что в нем проявляется смешанный план строения. Чашечка состоит из пяти чашелистиков, а все остальные органы трехчленны. Хотя почти у всех вересковых цветки актиноморфные, в отдельных случаях наблюдаются уклонения от лучевой симметрии. У некоторых видов рододендрона, например, при 5 чашелистиках число лепестков 3 и они образуют зев. Отмечено, что среди огромного числа видов эрики почти нет таких, у которых венчики были бы одинаковы (табл. 14). У некоторых вересковых (например, у багульника) лепестки свободные. Однако у большинства видов лепестки срастаются при основании или до половины своей длины так, что, потянув за один лепесток, мы отделим весь венчик. Наличие нектарного диска является характерным признаком семейства, хотя и в данном случае не обходится без исключений. У некоторых видов эрики диска нет. Он отсутствует и у бежарии, у которой обильный нектар вырабатывается внутренней стенкой чашечки. Причем нектар весьма клейкий, и поэтому цветки этого вечнозеленого кустарника называют смоляными или мухоловами, так как мелкие мушки, желая полакомиться нектаром, приклеиваются к нему. Бежария рассматривается как один из наиболее примитивных родов среди вересковых. Род бежария с 30 видами целиком американский (преимущественно южноамериканский). Внешне бежария напоминает рододендрон, и если последний часто называют "альпийской розой", то бежарию величают "розой Анд".

Таблица 14. Клетровые и вересковые: 1 - клетра бородчато-жилковая (Clethra barbinervis); 2 - эрика церинтовидная (Erica cerinthoides); 3 - эрика банксия, разновидность пурпурная (E. banksia var. purpurea)
О тычинках вересковых нужно рассказать особо. В пределах этого семейства разнообразие тычинок по их форме и взаимодействию между собой и с другими частями цветка очень велико и порой просто изумительно (рис. 44, 3-11). Обычно тычинки расположены двумя кругами: один - против лепестков, другой - против чашелистиков (как бы между лепестками). Общее для всех тычинок только то, что они двугнездны и вскрываются чаще всего порами. Кроме того, они часто снабжены придатками, по наличию которых вересковые назывались ранее Bicornes, т. е. двурогие, хотя тычинки обширного рода рододендрон, а также грушанковых и других лишены придатков. Эти придатки достигают венчика и служат при посещении насекомыми плечами рычага, способствующего высыпанию на них пыльцы. Пыльцевые зерна у большинства вересковых соединены по 4 в тетрады. В виде тетрад пыльца и попадает на рыльце, где все пыльцевые зерна сразу прорастают. Тетрады образуются у насекомоопыляемых растений, тогда как ветроопыляемые вересковые имеют пыльцу из одиночных зерен.
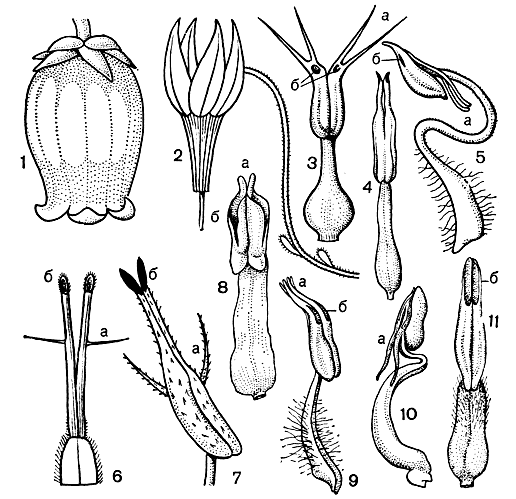
Рис. 44. Цветки и тычинки вересковых. Цветки: 1 - хамедафне болотная (Chamaedaphne calyculata); 2 - клюква (Oxycoccus). Тычинки: 3 - зеновия припудренная (Zenobia pulverulenta); 4 - хамедафне болотная (С. calyculata); 5 - лиония кустарниковая (Lyoniafruticosa); 6 - черника тычиночная (Vaccinium stamineum); 7 - черника обыкновенная (V.myrtillus); 8 - эллиотия кистевидная (Elliotia racemosa); 9 - гаультерия лежачая (Gaultheria procumbens); 10 - пиерис пышноцветный (Pieris floribunda); 11 - оксидендрум древовидный (Oxydendrum arboreum); a - тычиночные придатки; б - поры
У многих видов цветки яркие и приспособлены для опыления насекомыми. В тропиках и субтропиках есть вересковые, которые опыляются птицами. Приспособления для опыления весьма многообразны. У растений с повисающими колокольчатыми или кувшинчатыми цветками тычинки обычно не выступают из венчика и пыльники снабжены придатками, которые при касании их насекомыми способствуют высыпанию пыльцы. В цветках с широко раскрытым венчиком тычинки выставлены наружу и поэтому пыльники не имеют никаких придатков. Пыльца этих растений распространяется насекомыми в виде довольно крупных комочков, соединенных паутинообразными волокнами.
С одной из форм опыления у вересковых можно познакомиться на примере вереска обыкновенного (Calluna vulgaris), детально описанного в "Принципах экологии опыления" К. Фегри и Л. ван дер Пейла (1979). Чашевидный околоцветник четырехчленного цветка этого растения состоит из небольших розовых чашелистиков и еще более мелких и менее заметных розовых же лепестков. Нижние части лепестков мясистые, причем увеличение их толщины способствует раскрыванию цветка. Нижний лепесток отгибается несколько больше, чем остальные, что делает цветок слегка зигоморфным. Мясисты также нижние части тычиночных нитей, но их основания очень тонкие. Под завязью находится хорошо развитый нектарный диск. Нектар доступен даже самым короткохоботным насекомым. Просовывая свой хоботок между тычиночными нитями, насекомое раздвигает пыльники. Гнезда пыльников раскрываются латерально, и поэтому при раздвигании пыльников пыльца освобождается. Нектар производится в большом количестве, и вереск обыкновенный посещают многие мухи, перепончатокрылые и чешуекрылые. В почти горизонтально ориентированных цветках тычинки и столбик загибаются кверху, так что насекомому удобнее всего добраться до нектара снизу. Ко времени раскрывания цветка тычинки раскрываются и столь сильно отгибают кнаружи жесткие, усаженные волосками придатки пыльников, что их неизбежно должно коснуться всякое насекомое, проникающее хоботком к нектару. Действуя как плечо рычага, придаток способствует высыпанию пыльцы на насекомое. Благодаря тому что столбик с рыльцем, созревающим лишь после опорожнения пыльников, значительно высовываются над тычинками, уменьшаются шансы самоопыления. В дальнейшем продукция нектара уменьшается, тычиночные нити выпрямляются и пыльца рассеивается ветром. Это делает ветроопыление вполне возможным, тем более что продукция пыльцы чрезвычайно высокая. Но самой интересной особенностью в биологии опыления вереска обыкновенного является участие тениотрипса (Taeniothrips ericae) из отряда трипсов, или пузыреногих, открытое О. Хагерупом (1950) на Фарерских островах. В цветках вереска всегда можно обнаружить этих мелких насекомых, находящих здесь приют. Они находятся постоянно в движении и достигают частей, которые менее доступны крупным опылителям. Супруги Хагеруп (1953) показали, что аналогичное опыление с помощью трипсов наблюдается и у эрики тетраликс (Erica tetralix).
Весьма оригинальное приспособление для перекрестного опыления насекомыми наблюдается у американского рода калъмия (Kalmia, табл. 12 и 13). Тычинки, вытягиваясь в еще не раскрывающемся цветке, упираются в карманообразные полости на лепестках. Когда цветок раскрывается, то лепестки увлекают за собой тычинки, в нитях которых создается напряжение. Стоит тяжелому насекомому сесть на лепесток, как тычинки выскакивают из отгиба лепестка под действием своей пружинящей нити и хлопают пыльником по насекомому, осыпая его пыльцой.
Среди вересковых вполне обычно явление функциональной однополовости цветков. У рододендронов, багульника и луазелерии (Loiselenria), например, наблюдается протогиния, у "альпийской розы" (Rhododendron ferrugineum) - протандрия, а у черники обыкновенной (Vaccinium myrtillus) - слабая протандрия.
Весьма характерным для вересковых типом цветка является бокальчатый цветок (рис. 44, 1,2). Он присущ как ягодным вересковым (бруснике, чернике), так и неягодным, которых большинство. У клюквы, однако, полностью развившийся цветок напоминает цикламен, так как лепестки у него заворачиваются назад, а вперед выставлен длинный гинецей, к столбику которого тесно прилегают длинные пыльники; цветки при этом поникающие.
О. Хагеруп изолировал соцветия многих вересковых с помощью мешочков и убедился, что, несмотря на невозможность перекрестного опыления, плоды у них образовывались, но при условии, что цветки встряхивались. В природе встряхивание производит ветер, и поэтому ягод черники или голубики меньше в тех местах, где деревья сближены и гасят ветер. Для многих вересковых с поникшими цветками характерна гравитационная автогамия.
Плоды вересковых представляют собой септицидную или локулицидную коробочку, ягоду или же костянку. У некоторых видов рода гаультерия (Gaultheria) плод представляет собой коробочку, которую облекает сильно разросшаяся мясистая чашечка (рис. 45). Сходные плоды имеют дипликосия (Diplycosia) и пернеттия (Pernettya), причем у последней плод - настоящая ягода, сидящая на мясистой чашечке, и ягода выглядит двухэтажной (рис. 45).
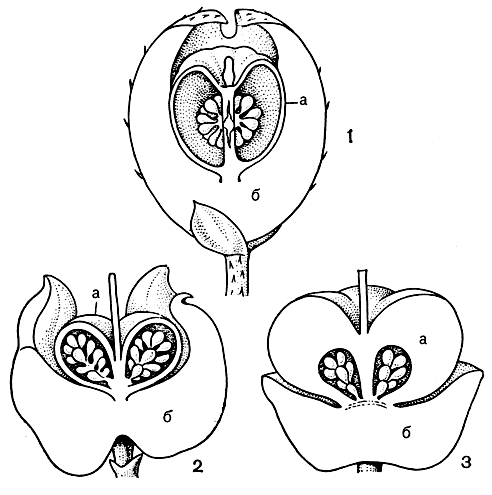
Рис. 45. Плоды вересковых: 1 - гаультерия щетинистоволосая (Gaultheria hispidula); 2 - гибрид гаультерии прижатой (G. depressa) и пернеттии крупно-рыльцевой (Pernettya macrostigma); 3 - пернеттия крупнорыльцевая; а - ягода; б - чашечка
Семена некоторых вересковых, снабженные мясистым придатком, растаскиваются муравьями. Ягодные вересковые распространяются птицами и млекопитающими. У сухоплодных растений после раскрытия коробочки семена распространяются ветром.
Весьма характерным свойством вересковых является их способность произрастать в весьма неблагоприятных условиях, на кислых почвах. Щелочных почв они, как правило, не выносят. Вереск, например, растет на сухих песчаных почвах, нередко среди лишайникового ковра, где способны выжить весьма немногие виды. Однако мы встречаем вереск и на болотах, где он растет на моховых буграх и выглядит ничуть не хуже, чем на песках. Долгое время считалось, что эрикоидные кустарнички болот, в том числе и вереск, не способны поглощать воду из-за ее низкой температуры. Таким образом, находясь в воде, растения как бы страдают от жажды ("физиологическая жажда"). Поэтому у них якобы и выработался эрикоидный облик. Однако в последние десятилетия было установлено, что эрикоидные растения на болоте испаряют огромное количество воды, следовательно, они и поглощают ее такое же количество, а их облик обусловлен иными причинами, среди которых на первом месте - азотная недостаточность болотных почв.
Жизнь на бедных почвах выработала у вересковых ряд приспособлений, важнейшее из них - симбиоз с грибами в форме микоризы. Корни почти всех вересковых тесно оплетают грибные нити, поставляющие им питательные вещества из перегноя. Грибы как бы в обмен получают вещества, вырабатываемые эрикоидным кустарничком. Существует экто- и эндомикориза. В последнем случае некоторые простейшие грибы (тело которых состоит всего из нескольких клеток) живут целиком в клетках корня вересковых и постепенно перевариваются ими. Микориза имеет огромное положительное значение в жизни вересковых. В некоторых случаях (например, у земляничного дерева - Arbutus, табл. 13) зараженные корешки превращаются в грушевидные клубеньки (микодоматии), эпидермальные клетки которых преобразуются в корневые волоски. Установлено, что семена вереска, например, прорастают только с помощью микоризы. Некоторые исследователи считают, что вересковые потому и живут на кислых почвах, что грибы, сожительствующие с ними, не выносят щелочных почв.
Семейство вересковые очень неоднородно и делится на ряд подсемейств. В 1971 г. английский ботаник П. Ф. Стивене предложил новую систему этого семейства, которая имеет ряд преимуществ по сравнению со старыми системами. В его системе вересковые делятся на 6 подсемейств.

Таблица 13. Вересковые и диапенсиевые: 1 - филлодоце голубая (Phyllobjce caerulea), Мурманская область; 2 - земляничное дерево красное (Arbutus andrachne), Крым; 3 - пиерис красивый (Pieris Formosa), оранжереи Ботанического института АН СССР в Ленинграде; 4 - кальмия широколистная (Kalmia latifolia),Батумский ботанический сад; 5 - рододендрон Делавэ (Rhododendron delavayi), оранжереи Ботанического института АН СССР в Ленинграде; 6 - луазелерия лежачая (Loiseleuria procumbens),Мурманская область; 7 - диапенсия лапландская (Diapensia lapponica), там же
Наиболее примитивно подсемейство рододендровых (Rhododendroideae), куда входят исключительно деревья и кустарники. Венчик у них свободнолепестный или сростнолепестный, обычно с широким зевом. Тычинки без придатков; пыльники вскрываются верхушечными порами или короткими или длинными щелями. В пыльниках обычно имеются шелковистые нити, связывающие пыльцу в комочки. Завязь 2-14-гнездная, часто покрытая железистыми волосками. Плоды - коробочки, обычно септицидные. Семена обычно многочисленные. В подсемейство входят американский род бежария (около 30 видов), рододендрон (вероятно, до 800 видов), мензисия (Menziesia, 7 видов в умеренной Азии и Америке), багульник (около 10 видов в северной умеренной зоне и в Арктике), эпигея (Epigaea, 3 вида в Аджарии и Лазистане, в Японии и в Северной Америке), кальмия (Kalmia, 8 видов в Северной Америке и на Кубе), филлодоце (Phyllodoce, 7 видов в северной умеренной зоне и в Арктике), родотамнус (Rhodothamnus, 2 вида в Восточных Альпах и на северо-востоке Малой Азии), монотипный род бриантус (Bryanthus, Камчатка, Командоры, Курилы и Япония), луазелерия (Loiseleuria, 1 циркумполярный вид, табл. 13) и некоторые другие.
Наиболее известен самый крупный в подсемействе род - рододендрон (табл. 13), приобретший исключительное значение в декоративном садоводстве. Большое разнообразие рододендронов, их высокие декоративные качества уже с давних пор привлекли к ним внимание. Рододендроны распространены в умеренной зоне северного полушария, но большая часть видов обитает в Восточных Гималаях и в Восточной Азии от Западного Китая до Японии, много видов рододендрона также в Северной Америке. Встречаются рододендроны также в горах Южной и Юго-Восточной Азии и Новой Гвинеи, а 1 вид обитает в Северной Австралии. В СССР 20 видов, произрастающих на Кавказе (главным образом в Западном Закавказье), на западе европейской части СССР, в Восточной Сибири и на Дальнем Востоке. Наиболее декоративны и в научном отношении наиболее интересны рододендрон Смирнова (R. smirnowii), произрастающий в нижнем и среднем поясе гор Аджарии, а за пределами СССР - в Артвинском округе Турции, и рододендрон Унгерна (R. ungernii) - узкий эндемик Аджарии и Лазистана (Турция). Эти два вечнозеленых древовидных рододендрона представляют собой третичные реликты, связанные родственными узами с рододендронами Гималаев и Восточной Азии. На Кавказе произрастают также два других вечнозеленых вида - рододендрон понтийский (R. ponticum) и рододендрон кавказский (R. caucasicum). В горах Кавказа произрастает также листопадный рододендрон желтый (R. luteum), известный под названием азалии понтийской (Azalea pontica). Отдельными островками он встречается также на западе европейской части СССР. Из дальневосточных рододендронов особый интерес представляют рододендрон Фори (R. fauriei) и рододендрон Шлиппенбаха (R. schlippenbachii). Из числа гималайских видов исключительно декоративен рододендрон Нутталла (R. nuttallii) - кустарник (иногда эпифитный) или небольшое дерево, называемый "царем рододендронов" из-за огромных (диаметром до 15 см) цветков. Очень декоративен также гималайский рододендрон древесный (R. arboreum) - крупный кустарник или дерево с темно-розовыми цветами.
Одним из интересных представителей подсемейства рододендроновых является также небольшой род эпигея. Виды эпигеи - низкие лежачие вечнозеленые кустарники, опушенные длинными рыжеватыми железистыми волосками. Цветки обоеполые или двудомные. Венчик у них опадающий, бокальчатый или воронковидный. Географическое распространение рода представляет собой яркий пример разорванного (дизъюнктивного) ареала: один вид эпигеи распространен в приатлантических областях США, другой - в Японии, а третий - среди вечнозеленого подлеска колхидского леса, среди реликтовых видов рододендрона в южной Аджарии и в Лазистане. Колхидский вид эпигея гаулътериевидная (E. gaultherioides) был некогда описан в качестве самостоятельного рода орфанидезия (Orphanidesia), но в 1941 г. А. Л. Тахтаджяном был переведен в род эпигея. Это одно из самых красивых растений Колхиды, которое в то же время является одним из наиболее интересных третичных реликтов. Как и некоторые рододендроны, он занесен в "Красную книгу".
В подсемейство эриковые (Ericoideae) входят вечнозеленые кустарники с мелкими листьями с сильно завернутыми краями. Венчик всегда сростнолепестный, остающийся. Пыльники вскрываются очень крупными порами или щелями; придатки пыльников, когда они имеются, представляют собой шпоры; шелковистые нити в пыльниках отсутствуют. Плод обычно локулицидная коробочка. Семян в каждом гнезде от 1 до многих. Сюда относятся прежде всего большой род эрика (более 500 видов, распространенных от Макаронезии до Западного Закавказья и в тропической и особенно в Южной Африке), монотипный род вереск и другие.
Обширное подсемейство вакциниевые (Vaccinioideae) включает древесные растения очень разного облика - от небольших деревьев до маленьких кустарничков. Венчик обычно сростнолепестный, редко свободнолепестный, обычно кувшинчатый или трубчатый. Пыльники очень разных типов, лишены шелковистых нитей. Завязь верхняя или нижняя, 3-10-гнездная. Плод - ягода, костянка или локулицидная коробочка, обычно с несколькими семенами в в гнезде, редко с одним. Сюда относятся арбутус (Arbutus, 20 видов в Западной Европе, Средиземноморье, Западной Азии и Северной и Центральной Америке; один вид - земляничное дерево красное - A. andrachne, табл. 13, растет у нас на Южном берегу Крыма и изредка встречается в Абхазии и Аджарии), Кассиопея (12 видов, распространенных циркумполярно и в Гималаях), энкиантус (Enkianthus, 10 видов, от Гималаев до Японии), подбел (1-2 вида в умеренных и холодных областях северного полушария), арктерика (Arcterica, 1 вид в Северо-Восточной Сибири, на Сахалине, Камчатке, Курилах и в Японии), гаультерия (Gaultheria, около 200 видов, распространенных преимущественно вокруг Тихого океана), вакциниум (Vaccinium, около 350 видов, распространенных в умеренных и холодных областях северного полушария, в горах тропиков, за исключением Африки, в Андах, в Южной Африке, на Мадагаскаре; клюква включается многими авторами в вакциниум) и др.
Среди представителей большого рода вакциниум целый ряд интересных и полезных для человека растений. Из видов, произрастающих на территории СССР, следует прежде всего отметить распространенный на Кавказе и в Северной Малой Азии (а также в Юго-Восточной Болгарии и в Северном Иране) третичный реликт - черника кавказская (Vaccinium arctostaphylos), представляющий собой крупный листопадный кустарник или небольшое деревце, высотой до 3 м, со съедобными плодами, произрастающий в колхидских лесах и на полянах. Ближайший вид - черника черемухолистная (V. padifolium) - обитает на острове Мадейра. Черника обыкновенная (V. myrtillus) - листопадный кустарник высотой 15-40 см, с угловато-ребристыми ветвями. Плод - более или менее шаровидная черная ягода с сизым налетом. Растет в умеренных и холодных областях Евразии и Северной Америки в хвойных и смешанных влажных или заболоченных лесах, а также в тундре и в высокогорьях. Ягоды съедобны, их употребляют в пищу в сыром и сушеном виде, а также для приготовления настоев, отваров и киселей. Голубика (V. uliginosum) - листопадный кустарник высотой от 30 см до 1 м с сизоватыми листьями и синими ягодами с сизоватым оттенком. Широко распространена в умеренных и холодных областях северного полушария. Ягоды съедобны, содержат сахар, кислоты и витамины А и С; их используют на варенье, сушат впрок. В отличие от упомянутых выше видов брусника (V. vitisidaea) - растение вечнозеленое. Это кустарничек высотой до 25 см с кожистыми листьями и многочисленными шаровидными ярко-красными ягодами. Растет в хвойных и смешанных лесах, кустарниках, тундрах, на высохших торфяниках в умеренных и холодных областях северного полушария. Плоды содержат в большом количестве сахара и органические кислоты, а также гликозид вакцинин и другие вещества. Их используют для приготовления варенья, повидла, маринадов, а также в кондитерской промышленности.
Далеко не всеми признаваемый и часто объединяемый с вакциниумом род клюква (Oxycoccus) содержит не более 4 видов, распространенных в умеренных и холодных областях северного полушария. Вечнозеленые, большей частью стелющиеся мелкие полукустарники с тонкими стеблями, мелкими кожистыми листьями и темно-красными ягодами. Клюква болотная (O. palustris) широко распространена по сфагновым и торфяным болотам, местами образует обширные заросли. Плоды клюквы - ценный пищевой продукт. Они содержат органические кислоты, пектиновые вещества, гликозид вакцинин, витамины С и Р. Широко используются в пищевой и ликеро-водочной промышленности.
Подсемейство грушанковые (Pyroloideae) небольшое; в него входят 4 рода и около 30 видов, представленные в умеренных и холодных областях северного полушария. Можно даже сказать, что это в основном таежное подсемейство, так как именно в таежной зоне сосредоточена большая часть его представителей. В отличие от всех ранее рассмотренных деревянистых вересковых грушанковые - травянистые вечнозеленые растения. У большинства видов имеется прикорневая розетка глянцевитых листьев, из которых выходит цветочная стрелка с кистевидным соцветием. Цветки 5-4-членные, открытые или бокальчатые. Последние имеют прямой столбик; у открытых он длинный, изогнутый, противопоставленный 10 (реже 8) тычинкам. У зимолюбки зонтичной (Chimaphila umbellata) цветки собраны в щиток, столбик короткий и толстый. Изящная одноцветна (Moneses uniflora) имеет один поникающий цветок (рис. 46), хорошо приспособленный для гравитационной автогамии.
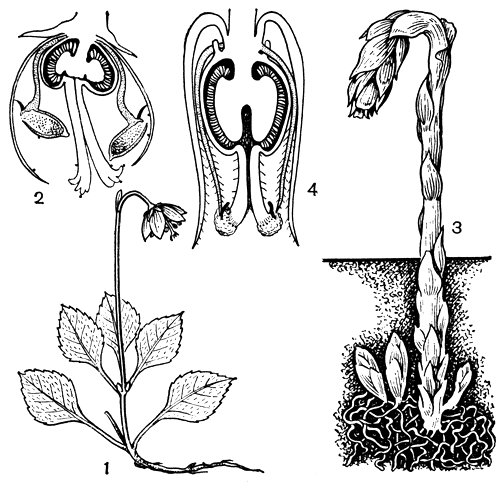
Рис. 46. Вересковые. Одноцветна (Moneses uniflora): 1 - общий вид растения. Грушанка средняя (Pyrola media): 2 - цветок в разрезе. Подъельник (Monotropa hypopithys): 3 - общий вид растения; 4 - цветок в разрезе
Нектарный диск у одних видов вовсе отсутствует, как у грушанки (Pyrola, рис. 46), у других фрагментирован, и только у зимолюбки он, как и у большинства вересковых, целиком окружен завязью. Рядом исследователей показано, что цветки грушанковых могут проходить две фазы развития. В первой фазе возможно только перекрестное опыление. При этом у одних представителей (грушанка, одноцветка, зимолюбка) его производят насекомые, у других, например у ортилии (Orthilia),- воздушные течения (в связи с чем у ортилии одиночные пыльцевые зерна, а у других грушанковых они объединены в тетрады). Вторая фаза происходит лишь в случае, если перекрестное опыление не произошло. При этом расположение тычинок и рыльца изменяется таким образом, чтобы обеспечить самоопыление цветка.
Плод у грушанковых - сухая локулицидная коробочка с большим количеством очень мелких семян, лишенных семядолей, и с зародышем, который состоит из нескольких клеток. Семена могут распространяться токами воздуха, которые возникают под пологом тайги.
Микориза для существования грушанковых еще более необходима, чем для других вересковых. Существует предположение, что продукты фотосинтеза необходимы грушанковым только для образования семян, а другие нужды растения вполне удовлетворяются питанием с помощью микоризы. В ряде случаев, впрочем, микоризное питание грушанок имеет для них трагические последствия. В благоприятных условиях микоризный гриб разрастается столь пышно, что вызывает отмирание корней у некоторых грушанок, а иногда и гибель растения. Симбиоз переходит в данном случае в односторонний паразитизм гриба.
Все представители грушанковых имеют длинное корневище и только у одноцветки оно заменено многолетними корнями. Корневище грушанки круглолистной (P. rotundifolia) может достигать в длину 6 м. Наземные побеги образуют целый клан сестринских особей.
После прорастания семени у некоторых грушанковых образуется так называемый предстебель. В стадии предстебля растение питается и развивается исключительно за счет микоризы, т. е. собственных органических веществ из неорганических в это время не вырабатывает. Из предстебля образуется корневище, а у одноцветки из него можно вырастить и наземные побеги. Так, в искусственных условиях был выращен предстебель грушанки круглолистной длиной 30 см. Установлено, что чем более редуцирован зародыш у разных видов грушанковых, тем ранее предстебель вступает в контакт с грибом. В пределах подсемейства можно проследить различные степени связи с грибами. Например, калифорнийская грушанка безлистная (P. aphylla) полностью перешла на довольствие за счет микоризы, и растение утратило зеленую окраску.
К подсемейству вертляницевых (Monotropoideae) относятся 10 родов и 12 видов. Большинство родов этого подсемейства монотипны. Вертляницевые полностью перешли на питание с помощью микоризы, в связи с чем утратили зеленую окраску. Многие из них имеют тусклый землистый цвет, но некоторые окрашены ярко. Так, монотипный род саркодес (Sarcodes), произрастающий на юго-западе США, где его называют снежным цветком, имеет малиновую окраску всего растения (т. 5, ч. 1, табл. 15). Распространены вертляницевые в умеренных областях северного полушария, а также в горах тропиков. Больше всего их в Северной Америке. Основным условием существования этих своеобразных растений является наличие на почве мощной подстилки из растительного опада, богатой органическими веществами, которые потребляются с помощью грибов. Недавно было установлено, что гифы одних и тех же грибов проникают в корни подъельника (Monotropa, рис. 46, 3-4) и ближайших деревьев. С помощью радиоактивных (меченых) элементов питания выяснили, что подъельник получает некоторые питательные вещества из корней дерева, используя для этого гифы грибов как бы в качестве шланга. Вещества, которые подъельник получает таким способом, необходимы ему для формирования семян, и благодаря этому приспособлению у подъельника нет зеленых частей, как у грушанок.
Вертляницевые имеют толстый, мясистый стебель с листьями в форме чешуи. Цветки у вертляницевых собраны в конечное поникающее соцветие, за исключением подъельника одноцветкового (М. uniflora). Цветки 5-членные или реже 4-8-членные. Цветок имеет колокольчатую форму, но у одних представителей лепестки срастаются у основания, а у других остаются свободными. Нектарный диск имеется у большинства представителей этого подсемейства (его нет у саркодеса и подъельника, у последнего, однако, еще сохраняются редуцированные сосочки в основании завязи). В большинстве случаев диск не сплошной, а разделен на лопасти. У подъельника одноцветного, как и у некоторых других видов, нектар выделяется утолщенными основаниями лепестков. Плод- локулицидная коробочка, часто перепончатая.
Монотропсис пахучий (Monotropsis odorata), растущий в Северной Америке, имеет любопытный цикл развития. Это растение живет под листовой подстилкой и на поверхности не появляется. Тем не менее считается, что оно опыляется насекомыми, так как имеет сильно пахнущие и окрашенные цветки. Пыльца у него высыпается на лепестки еще в бутоне. Цветочный побег монотропсиса пахучего появляется под листовой подстилкой осенью и перезимовывает. Весной рост его продолжается и цветки распускаются. Осенняя и весенняя формы принимаются иногда за разные растения и по ним были даже описаны два вида.
Плоды монотропсиса растаскивают муравьи, тогда как у подъельника семена очень легкие (0,000003 г) и снабжены "хвостиком", разносятся токами воздуха. Мелкие семена птероспоры (Pterospora) снабжены крылышком, значительно превосходящим по размеру само семя.
Подсемейство виттштейниевых (Wittsteinioideae) с монотипным австралийским родом виттштейния (Wittsteinia) связывает вересковые с семейством эпакрисовые. Виттштейния - это мелкий вечнозеленый кустарник со стелющимися побегами и мясистым плодом.
Семейство водяниковые (Empetraceae)
В семействе три рода: корежа (Corema, 2 вида), распространенная на северо-востоке США и в Канаде, на Азорских и Канарских островах, в Южной Португалии и Юго-Западной Испании; цератиола (Ceratiola, монотипный род), обитающая на юго-востоке США, и водяника, или вороника, или шикша (Empetrum), которая растет в таежном и тундровом поясах северного полушария, а также на юге Южной Америки, на островах Хуан-Фернандес, на Фолклендских островах и на островах Тристан-да-Кунья. По Р. Гуду, водяника насчитывает только 2 вида, в то время как В. Н. Васильев доводит число видов до 15-16.
Таким образом, в распространении водяниковых есть две особенности. Первая - это трансокеаническое распространение коремы (уже знакомый нам по другим вересковым тип распространения). Вторая особенность географии водяниковых - биполярное распространение водяники. В ледниковую эпоху, когда на всем земном шаре температуры понизились, ряд северных растений проник по горным цепям в Южную Америку, среди них была и водяника. По окончании холодного периода водяника почти исчезла из приэкваториальной области американских материков, но изолированно встречается еще и теперь на юге США и в Чили. Южноамериканская водяника оказалась географически обособленной, что привело к образованию особого вида - водяники красноплодной (E. erythrocarpum). Однако среди красноплодных экземпляров водяники в Южной Америке очень редко встречаются и черноплодные: возврат к исходному предку - водянике черноплодной (E. nigrum). Все водяниковые имеют типичный эрикоидный облик. Единственный вид цератиолы так и называется - цератиола эрикоидная (C. ericoides). Там, где совместно произрастают водяника и филлодоце из вересковых (табл. 13), их легко спутать, если на растениях нет цветков или плодов. Корема и цератиола - невысокие кустарники, а водяника всегда кустарничек со стелющимися по земле побегами. Она растет пятнами - куртинами, представляющими собой одну особь, которая, разрастаясь, постепенно занимает все большее и большее пространство, так как ветви образуют придаточные корни. В середине старой куртины зеленые побеги отмирают. Как и вересковые, водяника вступает в симбиоз с грибами.
Цветки водяниковых одиночные, в пазухах листьев (водяника) или же в малоцветковых верхушечных головках, мелкие, актиноморфные, обычно двудомные или иногда однодомные, редко обоеполые. Околоцветник довольно редуцирован, состоит из маленьких, едва заметных свободных долей. До сих пор еще не вполне ясно, является ли он простым или двойным. Обычно считается, что околоцветник состоит из 1-3 чашелистиков и 1-3 лепестков, но некоторые ботаники считают, что цветки водяниковых безлепестные и околоцветник состоит из одних только чашелистиков. Тычинок 2 (цератиола) или 3 (редко 4), чередующихся с внутренними долями околоцветника. Пыльцевые зерна соединены в тетрады такого же типа, как у вересковых. Нектарный диск отсутствует. Гинецей из 2-9 плодолистиков (водяника) или из 2 плодолистиков (остальные роды), с лопастным или более или менее раздельным коротким столбиком; завязь верхняя, 2-9-гнездная, с 1 семязачатком в каждом гнезде. Плод - сочная или сухая костянка с 2 или более косточками. Семена с прямым длинным зародышем, окруженным обильным мясистым эндоспермом. Водяника - растение преимущественно насекомоопыляемое. Она протогинична и имеет очень долговечное рыльце. Маленькие цветки водяники посещают бабочки, мухи и пчелы. Остальные два рода являются, возможно, ветроопыляемыми растениями.
Семейство эпакрисовые (Epacridaceae)
К вересковым, особенно к подсемейству рододендроновых, довольно близко стоит семейство эпакрисовых, распространенных главным образом в южном полушарии. В семействе около 30 родов и примерно 400 видов. Они обитают главным образом во внетропических областях Австралии, в Тасмании, Новой Зеландии и Новой Каледонии, а также в Юго-Восточной Азии (где некоторые виды достигают северных частей Индокитая и Филиппин), на Гавайских островах и в Патагонии (включая остров Огненная Земля). Обычно кустарники или небольшие деревья с большей частью тесно расположенными очередными или редко супротивными узкими цельными плотными листьями. У рода змеелистник, или дракофиллум (Dracophyllum, около 50 видов в Новой Зеландии и окружающих островах, в Новой Каледонии, Австралии, Тасмании и на острове Лорд-Хау), и близких родов листья с влагалищным основанием и параллельным жилкованием, чем очень напоминают листья однодольных. Поэтому некоторые виды внешне похожи на ирисы, а другие - на юкки. Для эпакрисовых характерен симбиоз с грибами.
Многие виды эпакрисовых образуют кустарниковые заросли. Некоторые обитают на болотах, в прибрежных дюнах, на скалистых склонах, в грасслендах, лесах, другие - в субальпийской зоне.
Цветки эпакрисовых мелкие, одиночные, в кистях или колосьях, иногда в метелках, актиноморфные, большей частью обоеполые. Чашелистиков обычно 5, свободных, остающихся; лепестков обычно 5, прикрепленных к нектарному диску; венчик сростнолепестный, трубчатый (рис. 47). Тычинок 5, приросших к венчику или реже свободных; иногда имеются чередующиеся с тычинками стаминодии; у большинства родов пыльники во время цветения 1-гнездные и вскрываются одной продольной щелью. Пыльцевые зерна одиночные или чаще соединены в тетрады. Гинецей обычно из 5 плодолистиков, с простым столбиком, заканчивающимся головчатым рыльцем; завязь верхняя, часто окруженная у основания нектарным диском, большей частью 5-гнездная, с несколькими или многими семязачатками в каждом гнезде. Плод - локулицидная коробочка или костянка. Семена с прямым цилиндрическим зародышем, окруженным обильным мясистым эндоспермом.
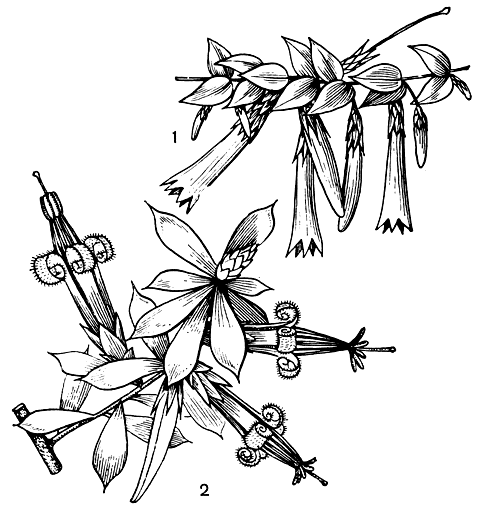
Рис. 47. Эпакрнсовые: 1 - эпакрис длинноцветковый (Epacris longiflora); 2 - стифелия зеленая (Styphelia viridis)
У эпакрисовых опыление производят преимущественно насекомые. Одним из приспособлений к перекрестному опылению является у них дихогамия.
Практическое значение эпакрисовых ничтожно. Некоторые виды дракофилума имеют декоративное значение, а сочные плоды ряда видов местное население употребляет в пищу.
У представителей родственного американского семейства цириллиевых (Cyrillaceae, рис. 48) имеет место партенокарпия.
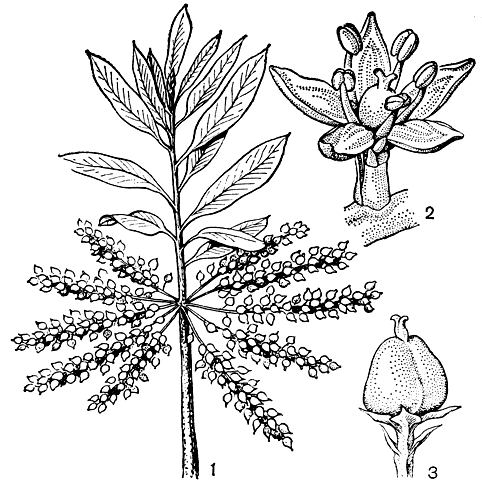
Рис. 48. Цирилла кистецветковая (Gyrilia racemi flora): 1 - ветвь с плодами; 2 - цветок; 3 - отдельный плод
Семейство диапенсиевые (Diapensiaceae)
Состоит из 6 родов и около 20 видов, распространенных в холодных и умеренных областях северного полушария, главным образом в горах и в Арктике. Род бернексия (Berneuxia, 2 вида) обитает в Восточном Тибете, Юго-Западном Китае и в Верхней Бирме. Три вида диапенсии (Diapensia) встречаются в Гималаях и в Западном Китае, а четвертый вид обитает в Арктике и Субарктике, а также на северных горах. Оба вида схизокодона (Schizocodon) встречаются только в Японии. Род шортия (Shortia, рис. 49, 2), насчитывающий 8-9 видов, распространен главным образом в Юго-Западном Китае, на острове Тайвань и в Японии, но 1 вид обитает в юго-восточных штатах США. Монотипные же роды галакс (Galax) и пиксидантера (Pyxidanthera) встречаются только на юго-востоке США. Среди диапенсиевых имеются вечнозеленые травы (шортия), полукустарнички (пиксидантера), подушковидные кустарнички (диапенсия, рис. 49, 3) и низкие кустарники (галакс). Диапенсия и пиксидантера имеют эрикоидный облик. Представители других родов более похожи на грушанок. Листья очередные, цельные, в отличие от вересковых, лишены железистых волосков.
Цветки у некоторых диапенсиевых одиночные, у других собраны в укороченные кисти. Они обоеполые, актиноморфные, чашечка остающаяся, чашелистиков 5, свободных или сросшихся. Венчик сростнолепестный, 5-лопастный, но у галакса лепестки, сросшиеся только у основания. Тычинок 5, приросших к венчику и чередующихся с его долями (рис. 49, 2). У большинства родов (за исключением пиксидантеры и большинства видов диапенсии) имеется второй круг тычинок, расположенных против лепестков, но они превращены в стаминодии. У галакса стаминодии и тычинки срослись вместе, образуя трубочку. Пыльники 2-гнездные или редко 1-гнездные (галакс), вскрывающиеся продольной щелью или редко поперечно (пиксидантера). Пыльцевые зерна всегда одиночные. Нектарный диск отсутствует, но нектар притом иногда выделяется основанием завязи довольно обильно. Гинецей из 3 плодолистиков, со столбиком, заканчивающимся 3-лопастным рыльцем; завязь верхняя, 3-гнездная, с несколькими или многочисленными семязачатками. Плод - локулицидная коробочка. Семена мелкие, с обильным мясистым эндоспермом вокруг прямого или слегка согнутого зародыша.
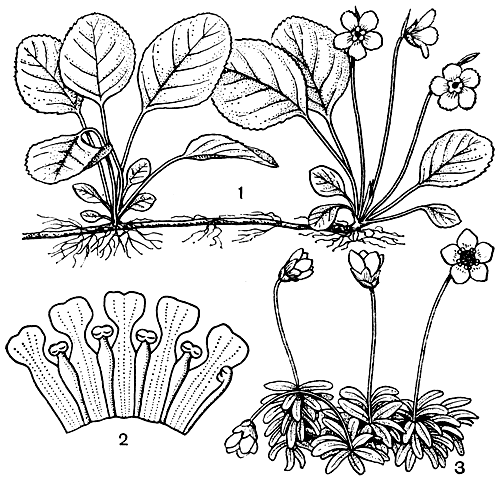
Рис. 49. Диапенсиевые. Шортия галаколистная (Shortia galaxifolia): 1 - ветвь с цветками. Пиксидантера (Pyxidanthera): 2 - препарированный венчик цветка, показывающий положение тычинок, сросшихся с лепестками. Диапенсия лапландская (Diapensia lapponica), 3 - ветвь с цветками
У диапенсиевых цветки белые или слегка розоватые, но у гималайских диапенсий они пурпурные или желтые, иногда с приятным запахом. Процесс опыления изучен только у арктической диапенсий лапландской (D. lapponica, табл. 13), но и то недостаточно основательно. Это растение опыляется насекомыми, причем самыми разными. В некоторых цветках диапенсий раньше созревают тычинки, в других - завязи. Пыльники вскрываются иногда еще в бутоне, и вполне возможно самоопыление цветков. В цветках шортии гинецей созревает раньше тычинок. О насекомоопыляемости диапенсий косвенно свидетельствует тот факт, что пыльца у нее покрыта бородавочками (для лучшего сцепления с телом насекомого).
В плодах диапенсиевых обычно большое количество очень мелких семян, распространяющихся ветром. Семена шортии часто прорастают прямо в коробочках, не успев попасть на землю.
Диапенсиевые весьма неприхотливы в отношении почв, чем напоминают вересковые и водянику. Это сходство усиливается наличием микоризы и анатомическим строением стеблей и листьев.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'