Порядок имбирные (Zingiberales) (С. С. Морщихина)
Семейство стрелитциевые (Strelitziaceae)
Стрелитциевые объединяют всего 3 рода и 7 видов и имеют удивительным образом разорванный ареал. Знаменитое "дерево путешественников" - равенала мадагаскарская (Ravenala madagascariensis, рис. 217, 1-4) представляет монотипный род, эндемичный для Мадагаскара. Один вид рода фенакоспермум (Phenakospermum, рис. 217, 5-7) произрастает в тропических дождевых лесах Южной Америки. В восточной части Южной Африки распространено 5 видов стрелитции (Strelitzia, рис.. 218, табл. 46, 1, 2). В семействе преобладают древовидные растения с высоким древеснеющим стволом (но без вторичного роста) и двурядно расположенными листьями. До высоты 15 м поднимаются мощные стволы равеналы, увенчанные гигантским веером из 20-30 листьев. Фенакоспермум, по облику похожий на равеналу, имеет не столь высокий ствол, лишь до 9 м. Стрелитция Николая (S. nicolai, табл. 46, 1) вырастает до 5 м, у других видов стволы намного меньше, а у стрелитции королевской (S. rеginae, рис. 218, табл. 46, 2) стебель совсем укорочен и только листья поднимаются над поверхностью земли.
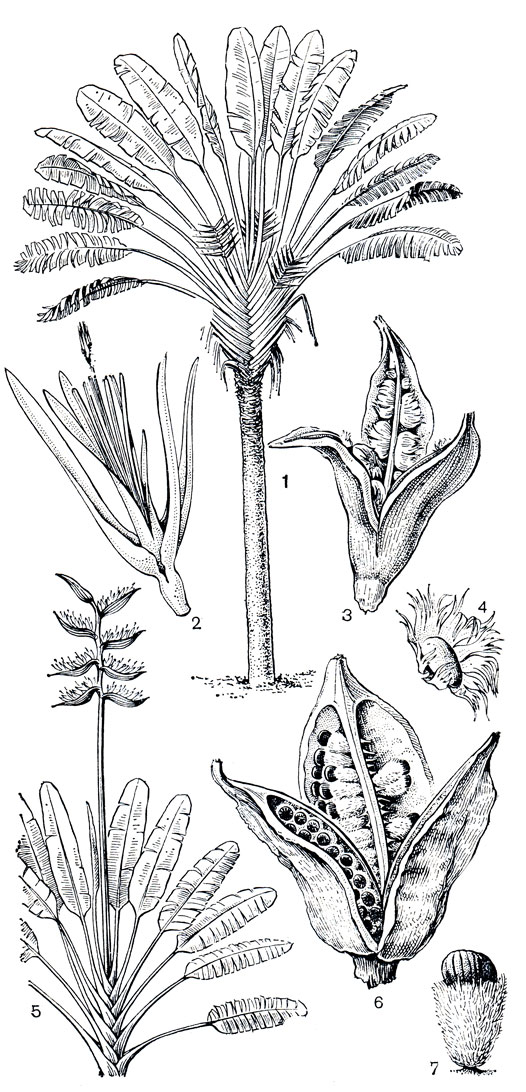
Рис. 217. Стрелитциевые. Равенала мадагаскарская (Ravenala madagascariensis): 1 - цветущее растение; 2 - цветок; 3 - открывшийся плод; 4 - семя с ариллусом. Фенакоспермум гвианский (Phenakospermum guianense): 5 - цветущее растение; 6 - открывшийся плод; 7 - семя с ариллусом
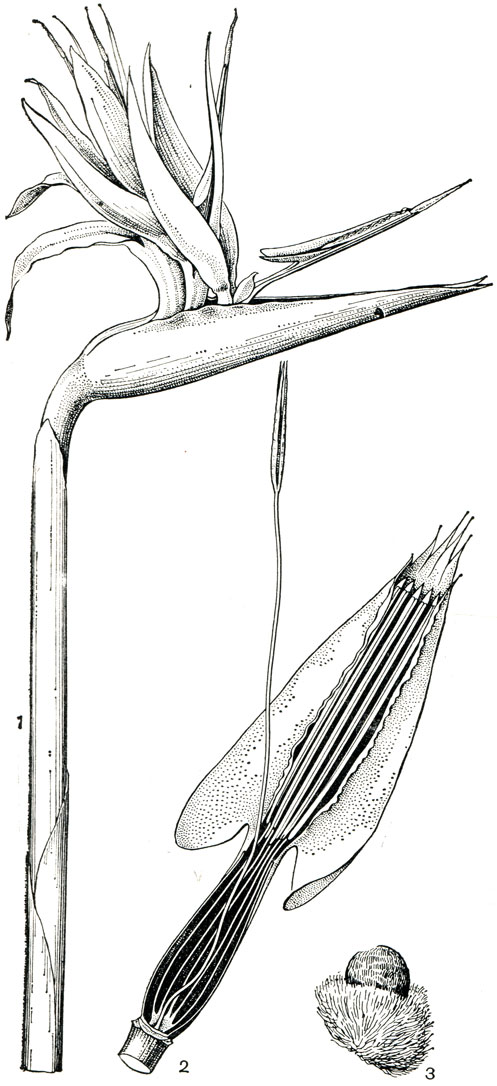
Рис. 218. Стрелитция королевская (Strelitzia reginae): 1 - соцветие; 2 - стреловидный орган из лепестков раскрыт, столбик поднят, видны 5 тычинок; 3 - семя с ариллусом

Таблица 46. Стрелитциевые и банановые: 1 - стрелитция Николая (Strelitzia nicolai), оранжереи Ботанического института АН СССР в Ленинграде; 2 - стрелитция королевская (S. reginae), оранжереи фирмы "Цветы" в Ленинграде; 3 - банан японский (Musa basjoo), Батумский ботанический сад; 4 - банан текстильный (М. textilis), о. Маврикий, ботанический сад
У самого основания стволов стрелитциевые образуют отпрыски, лишь у фенакоспермума горизонтальные симподиальные корневища, покрытые чешуевидными листьями, растут под землей и уже на некотором расстоянии от материнского ствола развивают новые стебли. Листья стрелитциевых с длинными черешками, имеющими влагалища, и овальными или удлиненными пластинками. По сторонам от крупной главной жилки - многочисленные, слегка изогнутые, почти параллельные боковые жилки, по которым крупные листья разрываются от ветра; целыми их можно увидеть лишь у экземпляров, выращенных в оранжереях. У равеналы длинные влагалища с сомкнутыми краями наполнены выделенной тканями жидкостью, значение которой в жизни растения пока неясно. В этой жидкости кишат микроорганизмы, личинки насекомых и даже мелкие земноводные, что делает эту воду абсолютно непригодной в качестве питья для усталых путешественников вопреки распространенной версии, породившей бытующее название растения.
Каждый новый лист у стрелитциевых развивается под защитой влагалища предыдущего листа, он трубчато свернут, и наружная его половина, стесненная влагалищем, остается уже внутренней, так что весь лист несколько асимметричен. В клетках листьев многочисленные включения кремнезема и кристаллы оксалата кальция. Проводящая система стволов равеналы лишена сосудов, они имеются лишь в ее корнях, а у фенакоспермума и стрелитции и в корнях, и в стеблях есть сосуды с лестничной перфорацией.
Соцветия образуются обычно в пазухах листьев, только фенакоспермум имеет верхушечное соцветие. На главной оси соцветия двурядно расположены крупные кожистые кроющие листья. Они продольно сложены, словно ножны, и заключают в себе парциальные соцветия - завитки, в которых цветки обращены лишь к верхней стороне. Цветение начинается с нижних завитков и в каждом из них также от основания парциального соцветия. Соцветия равеналы длиной более 1,5 м несут по 10-12 пар-циальных соцветий; ее огромные, длиной около 50 см, кроющие листья по форме напоминают ладью и содержат 10-15 последовательно открывающихся цветков.
Цветки стрелитциевых снабжены прицветничком, обоеполые, у равеналы и фенакоспермума со слабо выраженной зигоморфией, а у стрелитции - зигоморфные. Сегменты околоцветника располагаются по 3 в 2 круга. У равеналы и фенакоспермума все они лепестковидные, белые, почти одинакового размера; лишь один сегмент внутреннего круга, обращенный к главной оси соцветия, несколько уже и короче остальных. В цветках стрелитции форма, окраска и функции сегментов наружного и внутреннего кругов значительно дифференцированы. Тычинки с 2-гнездными линейными пыльниками имеют упругие и прочные нити. Цветки равеналы содержат 6 тычинок, а у представителей других родов их только 5, не развитой остается тычинка у медианного лепестка, обращенного к главной оси соцветия. Цветки стрелитциевых протандричны. Для них характерен так называемый "механизм взрыва", когда зажатые лепестками тычинки мгновенно освобождаются опылителем и выбрасывают пыльцу. Пыльцевые зерна стрелитциевых крупные, с безапертурной оболочкой, слипаются хлопьями благодаря нитевидным образованиям, окружающим их в пыльнике. Гинецей стрелитциевых синкарпный, из трех плодолистиков, завязь 3-гнездная, нижняя, с многочисленными анатропными семязачатками; столбик обычно с 3-лопастным рыльцем. В цветках равеналы лопасти рыльца короткие, а у стрелитции - длинные, немного неравные, с волнистой воспринимающей поверхностью, обращенной кверху. В тканях верхней части завязи расположены септальные желёзки, открывающиеся у основания столбика. Нектар наполняет всю ладью кроющего листа и вытекает за его края. Цветки стрелитциевых не имеют заметного запаха, но яркой окраской и обилием нектара привлекают птиц.
Английский ботаник Дж. Ф. С. Эллиот (1890) наблюдал в Форт-Дофине на цветках равеналы мадагаскарской нектарниц (Nectarinia sonimanga). Особенно специализированы к птицеопылению цветки стрелитции. У открывающегося цветка два ланцетных чашелистика, обращенных к главной оси соцветия, стоят почти вертикально над горизонтально расположенным кроющим листом. Под чашелистиком тоже почти вертикально поднят небольшой лепесток. Два других, значительно более крупных, остаются вместе с третьим чашелистиком зажатыми кроющим листом. Эти парные лепестки срастаются в единый стреловидный заостренный орган с продольным килем. Сложенные края этого органа держат "в плену" упругий столбик и 5 длинных тычинок. У стрелитции королевской ярко-синий или фиолетовый цвет лепестков контрастирует с оранжевым цветом чашелистиков, что, несомненно, играет роль в привлечении птиц. Птица, садясь на края стреловидного органа, отгибает их и тем самым освобождает упругие тычинки, которые, выгибаясь, растрескиваются и выбрасывают пыльцу, обдавая ею грудку птицы. Благодаря такому триггериому механизму стреловидный орган словно катапультирует пыльники, но продолжает еще удерживать столбик и длинные лопасти рыльца, которые освобождаются лишь при повторных посещениях цветка нектарницами и опыляются принесенной ими пыльцой других цветков.
Плоды стрелитциевых - локулицидные коробочки с жесткими деревянистыми стенками, семена с ярким ариллусом. У равеналы коробочки длиной 7-10 см содержат уплощенные, овальные, почти черные семена с пленчатым ариллусом чистого голубого цвета. Черные, яйцевидные, немного уплощенные семена фенакоспермума несут крупный красновато-оранжевый пушистый ариллус из многочисленных гигроскопичных волосков. Похожий ворсистый ариллус и у округлых семян стрелитции. Ариллус, по-видимому, играет роль при раскрывании плодов и, несомненно, привлекает внимание птиц, однако о распространении семян стрелитциевых известно, к сожалению, очень мало, для этого нужны тщательные наблюдения в природе.
Зародыш стрелитциевых большей частью прямой (у равеналы - изогнутый), лежит в перисперме и окружен остатками эндосперма. Семена равеналы, высыхая, быстро теряют жизнеспособность, но во влажной почве могут долго сохраняться и прорастать при осветлении леса после вырубки. Поэтому равенала быстро заселяет просеки, заброшенные плантации, обочины дорог. Это светолюбивое растение распространено преимущественно на востоке Мадагаскара на опушках и полянах влажных тропических лесов, на болотистых прибрежных равнинах, но может удерживаться и в саванне, теряя часть листьев в засушливый сезон. Фенакоспермум - обитатель влажных тропических лесов по низким болотистым берегам Амазонки и ее притоков. Стрелитции, в отличие от первых двух родов,- растения субтропических лесов Южной Африки, но тоже предпочитают влажные и открытые местообитания, поляны, берега рек.
Стрелитции с их необычными соцветиями, напоминающими голову фантастической птицы, издавна заняли место среди растений оранжерейных коллекций. Стрелитция Николая была описана Э. А. Регелем по экземпляру, зацветшему впервые в 1853 г. в Санкт-Петербургском ботаническом саду. Стрелитцию королевскую выращивают в промышленных хозяйствах Европы для букетов. Равенала часто украшает сады во всех тропических странах. На Мадагаскаре ее стволы используют для построек и на топливо. Семена равеналы съедобны.
Семейство банановые (Musaceae)
Это небольшое палеотропическое семейство состоит из 2 родов и примерно 50 видов.
Род банан (Musa, рис. 219, табл. 46, 2, 2) объединяет свыше 40 видов, распространенных в тропической Южной Азии, на островах Малайского архипелага, Новой Гвинее, в Северо-Восточной Австралии и на островах Тихого океана. Далее всего к западу распространен банан Маклая (М. maclayi, рис. 219, 7), растущий на Новой Гвинее, Таити, Новой Каледонии и Фиджи, откуда он, видимо, занесен на Гавайские острова. Южная граница банана проходит в Квинсленде у 16° ю. ш. Центр сосредоточения наибольшего числа видов банана и происхождения его культурных форм находится в Индии, на полуострове Индокитай, где произрастает около 20 видов банана, и на островах Малайского архипелага, несколько уступающих по числу его видов. Некоторые виды банана выходят за пределы тропиков в теплые субтропические районы. В Индии, в Ассаме и в Юго-Западном Китае бананы встречаются до 27° с. ш. Такой же широты достигает банан японский (М. basjoo, табл. 46, 3) на островах Рюкю.
Род энсета (Enseta) состоит из 7 видов, произрастающих в тропической Африке и Азии. От Камеруна до Эфиопии и к югу до Трансвааля распространена энсета вздутая, или "абиссинский банан" (Е. ventricosum, рис. 219, 8-10). На Мадагаскаре встречается только 1 вид - энсета Перръе (Е. perrieri). В Азии ареал энсеты простирается от Северо-Восточной Индии, Бирмы и Таиланда до Южного Китая, Филиппин, Новой Гвинеи и Явы. Здесь наиболее распространенным видом является энсета сизая (Е. glaucum).
Банановые - гигантские многолетние травы с мощными подземными корневищами и укороченными клубневидными стеблями, которые почти не выступают над землей и несут спирально расположенные огромные листья с необычайно длинными влагалищами. Влагалища охватывают друг друга и образуют плотную многослойную трубку ложного стебля. Ложные стебли нередко достигают в высоту 5-6 м. Растения энсеты вздутой возвышаются до 13 м, а банан огромный (М. ingens), растущий на Новой Гвинее, бывает высотой до 15 м и имеет листья длиной 5-6 м, шириной 0,6-1 м. Наряду с такими гигантами существует банан шершавоплодный (Musa lasiocarpa), растущий в горах провинции Юньнань в Китае, который представляет собой растение не выше 60 см с листьями длиной до 30 см.
Так же как у стрелитциевых и других представителей порядка имбирных, листья банановых развиваются внутри влагалища предыдущего листа, трубчато свернуты и потому несимметричны. У банана правая, наружная половинка листа, стесненная трубкой ложного стебля, всегда уже внутренней, левой. С увеличением числа листьев диаметр полости ложного стебля сокращается и асимметрия развивающихся листьев увеличивается. От мощной главной жилки листа почти под прямым углом с равными интервалами отходят боковые жилки, по которым лист легко разрывается от ветра и дождей. Подобно равенале, бананы в природе почти всегда имеют разорванные листья.
Листья банановых нередко покрыты восковым налетом. Устьица окружены несколькими побочными клетками, мало отличающимися от остальных эпидермальных клеток. Сосуды проводящей системы имеют поперечные стенки с простой перфорацией или заканчиваются косой стенкой с лестничной перфорацией. Вдоль проводящих пучков располагаются ряды клеток с утолщенными стенками и включениями кремнезема. Кристаллы кремнезема и оксалата кальция очень часто встречаются и в обычных паренхимных тонкостенных клетках вегетативных органов. Клеточный сок, вытекающий при порезах ложного стебля, окисляясь на воздухе, становится буровато-оранжевым, а у некоторых видов, например у банана Маклая, он постоянно окрашен в розоватый или фиолетовый цвет благодаря содержанию антоциана.
Растут бананы поразительно быстро. Огромные, 7-8-метровые ложные стебли вырастают всего за 8-10 месяцев, и в этом возрасте растения обычно переходят в репродуктивную фазу. Листья перестают образовываться. Точка роста, заключенная в трубке листовых влагалищ, развивает цветоносный стебель, который быстро растет внутри ложного стебля, и через несколько недель наверху между листьями появляется крупное верхушечное соцветие. После цветения и плодоношения вся надземная часть отмирает. У основания ложного стебля бананы образуют боковые подземные побеги. Они растут некоторое время горизонтально, затем поворачивают к поверхности почвы и дают начало новым ложным стеблям с листьями. Виды энсеты являются монокарпическими растениями. Они обычно отмирают, не образуя отпрысков. Известны лишь отдельные случаи образования вегетативного потомства у старых, отмирающих клубневидных стеблей энсеты.
Соцветие банановых несет на мощной оси крупные плотные кроющие листья, которые в отличие от стрелитциевых, расположены не двурядно, а по спирали, против часовой стрелки. В пазухах кроющих листьев боковые ветви соцветия имеют такую укороченную и приросшую ось, что выглядят просто поперечными выступами главной оси, несущими два ряда цветков. Развитие цветков происходит закономерно, начиная справа, поочередно во внутреннем и наружном ряду. Очевидно, как и у стрелитциевых, это тоже завиток, но претерпевший сильный метаморфоз. У некоторых видов развивается лишь один ряд цветков. Молодое соцветие банановых выглядит как огромная почка, где кроющие листья сближены, сложены черепитчато., как у видов энсеты или у банана текстильного (М. textilis, табл. 46, 4). У культурных бананов они полностью окутывают почку. Эти листья могут быть зелеными, но чаще розовые или ярко-красные, пурпурные либо с фиолетовым оттенком. Раскрываются они поочередно, обнажая цветки, число которых в двурядных парциальных соцветиях может достигать 40. В солнечную жаркую погоду могут раскрываться по 2-3 боковых соцветия, в дождливую они раскрываются вяло, по одному, с большими интервалами. У бананов кроющие листья опадают на вторые сутки, а у энсеты сохраняются и в соплодии. Ось соцветия непрерывно растет, междоузлия его вытягиваются, и на конце все время остается почка, размеры которой по мере цветения уменьшаются.
Цветки банановых зигоморфные, обычно однополые,. В первых, нижних парциальных соцветиях развиваются женские цветки, дающие плоды; в последующих иногда обоеполые, но не завязывающие плодов; затем, до самого верха, образуются мужские цветки, которые после цветения опадают. Сегменты околоцветника у банановых лепестковидные, беловатые или желтые, пять из них срастаются вместе, одевая цветок снаружи (рис. 219, 4, 5). Свободным остается один сегмент внутреннего круга, обращенный к оси соцветия. В цветках банановых обычно 5 тычинок, шестая (у основания свободного лепестка) превращена в стаминодий. В цветках энсеты вздутой развиты все 6 тычинок. Тычинки имеют 2 линейных пыльника, растрескивающихся продольно. Пыльцевые зерна крупные, тяжелые, с безапертурной оболочкой. Гинецей синкарпный, из трех плодолистиков; завязь нижняя, 3-гнездная, с многочисленными анатропными семязачатками, расположенными двумя рядами в центральном углу гнезда. Столбик несет 3- или 6-лопастнре головчатое рыльце. В верхней части завязи в ткани погружены септальные нектарные железки, открывающиеся у основания столбика. Особенно сильно они развиты в мужских цветках, где выделение нектара - единственная функция редуцированной завязи. У культурных бананов один женский цветок дает в сутки 0,10-0,27 г нектара, а мужской - 0,42-0,59 г.
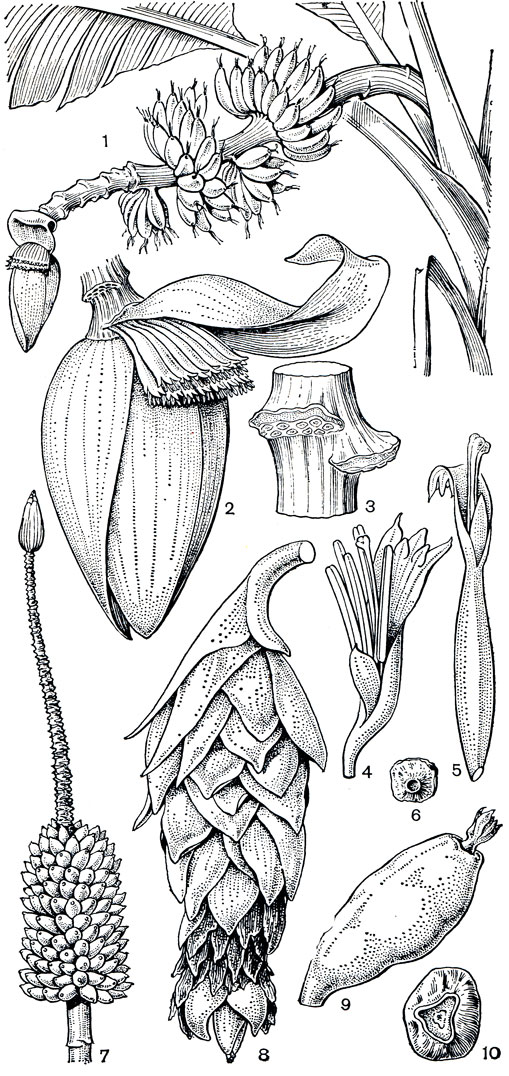
Рис. 219. Банановые. Банан заостренный (Musa acuminata): 1 - соцветие, у основания завязавшиеся плоды; 2 - верхушка соцветия с мужскими цветками; 3 - часть оси соцветия со следами от опавших цветков; 4 - цветок с недоразвитым гинецеем, функционально мужской; 5 - женский цветок; 6 - семя. Банан Мак лая (М. maclayi): 7 - соплодие. Энсета вздутая (Ensete ventricosum): 8 -соплодие; 9 -плод; 10 - семя
Бананы с повисающими соцветиями расцветают вечером, а энсета - в полночь. Цветки издают специфический запах, привлекающий летучих мышей. Ван дер Пэйл (1936) наблюдал на бананах летучих мышей из подсемейства длинноязычковых (Macroglossinae). Подробные наблюдения за опылением бананов провел на Яве и Суматре индонезийский ботаник Назар Hyp (1976). Ночью бананы посещают летучие мыши. Желудки их полны нектара, головы покрыты пыльцой, а на соцветиях остаются следы их когтей. Наутро цветки посещают птицы и многочисленные насекомые. Бананы с пря-мостоячими соцветиями зацветают утром и опыляются главным образом яркими птицами-нектарницами (Nectarinia calcostetha) и мелкими млекопитающими - тупайями. Тупайи, подобно белкам, живут на деревьях и питаются в основном плодами, часто лакомятся нектаром и могут служить переносчиками пыльцы. Интересно, что бананы с вертикально стоящими соцветиями, цветущие днем, имеют цветки, лишенные запаха и выделяющие более жидкий нектар. Н. Hyp наблюдал среди опылителей банана также бабочек, пчел, ос и муравьев.
Многие виды банана, например банан бархатистый (М. velutina) и банан кроваво-красный (М. sanguinea) из Ассама, обладают способностью завязывать плоды и семена и при самоопылении. Это свойство присуще главным образом видам, обитающим на границе ареала, и помогает им выжить в крайних условиях. В большинстве случаев при отсутствии естественных опылителей бананы не плодоносят или иногда образуют бессемянные партенокарпические плоды.
Если цветущие бананы активно посещают животные, то после созревания плодов их буквально атакуют летучие мыши, многочисленные птицы, обезьяны и тупайи. В лесах Явы трудно найти неповрежденные животными зрелые плоды.
Плод банановых - ягода с кожистой оболочкой и сочной мякотью, в которую погружены многочисленные семена. У энсеты плоды довольно сухие, но не раскрывающиеся. Растрескиваются только зрелые плоды банана схизокарпного (М. schizocarpa), растущего на северовосточном побережье Новой Гвинеи. Это редкое для банановых свойство отражено в его видовом эпитете, который буквально означает - "с растрескивающимся плодом". Иногда, правда, раскрываются также и плоды банана бархатистого.
Плоды бананов - удлиненные, цилиндрические, несколько граненые и серповидно изогнутые - знакомы всем. Кроме этой характерной формы, у некоторых видов встречаются короткие овальные, почти круглые или, наоборот, тонкие длинные, роговидно заостренные плоды. При созревании плоды желтеют или краснеют. Соплодия банановых могут быть очень крупными. Так, например, мадагаскарская энсета Перрье дает соплодия массой 25-30 кг, содержащие до 200 плодов, а соплодие культурных сортов банана может состоять из 300 плодов общей массой 50-60 кг.
В плодах банановых 50-100, а иногда даже до 200 семян. Семена уплощенные, округлые или неправильной формы, с твердой темно-коричневой или черной оболочкой. В отличие от стрелитциевых, банановые не имеют ариллуса. На ранних стадиях развития на семяножках имеются нити, которые можно считать таким образованием, но в дальнейшем они дегенерируют. Наличие сочных душистых плодов обеспечивает зоохорное (с участием животных) распространение семян. У банана семя диаметром 3-11 мм, у энсеты семена крупнее, диаметром до 17 мм. Зародыш у банана прямой, а у энсеты - изогнутой Т-образной формы. Питательные вещества запасаются в мучнистом перисперме, эндосперм мало развит. Семена могут долго сохранять всхожесть в почве, покрытой растительным опадом, и прорастают при осветлении участка после вырубок или ветровала. Прорастание подземное, главный корень очень рано отмирает, уступая место многочисленным придаточным корням. Первый лист проростка - влагалищный, не имеет развитой пластинки.
Банановые - обитатели солнечных, открытых полян, лесных опушек, берегов рек. Они образуют заросли во вторичных формациях, на вырубках, заброшенных плантациях, по обочинам дорог. В глубине тенистых лесов они перестают плодоносить и постепенно гибнут. Исключение составляет банан огромный (М. ingens), растущий в густых нотофагусовых лесах в горах Новой Гвинеи. Его сеянцы хорошо развиваются и в густом подлеске. Иногда причиной гибели бананов является не наступление лесной растительности, а конкуренция со злаками, которой они не выдерживают. В сообществах со злаками лучше других уживается банан Балъбиса (М. balbisiana) и энсета Омбле (Е. homblei), встречающиеся в светлых лесах и саваннах. Запас воды и питательных веществ в клубневидном стебле помогает видам энсеты переживать засушливый период, растения теряют листья и иногда выдерживают пожары, сохраняя точку роста, спрятанную в чехле ложного стебля. Довольно выносливы к засухе и виды банана, распространенные в муссонном климате Юго-Восточной Азии. Большинство бананов является обитателями влажного тропического климата и приурочено к небольшим высотам над уровнем моря. В то же время есть горные виды, которые плохо переносят постоянно влажный и жаркий климат. Банан Маклая, бессемянный на небольших высотах, образует семена в горах, на высоте 900-1100 м над уровнем моря. Банан огромный, поднимающийся в горах Новой Гвинеи до 2100 м, гибнет от грибковых заболеваний в посадках близ моря.
Банан - важнейшая культура тропического земледелия. Во многих развивающихся странах экспорт бананов составляет основу экономики. Мировое производство плодов составляет около 24 млн. т и главным образом сосредоточено в странах Латинской Америки. Почти четверть урожая приходится на Индию, Малайзию, Индонезию. Более миллиона тонн бананов выращивают в странах Африки. Создание выносливых сортов позволило продвинуть культуру бананов до 30° с. ш. и 31° ю. ш. в теплые субтропические районы, в Ливан, Испанию, Флориду. На Канарские острова бананы были завезены португальскими мореплавателями еще в 1482 г. Неудивительно, что род банан стал известен ботанике по культурному экземпляру, описанному Карлом Линнеем в 1753 г. в первом издании его знаменитого труда "Species Plantarum" и названному им бананом райским (Musa paradisiaca). Во втором издании своей работы (1763) Линней добавил банан браминов, или банан мудрецов (М. sapientum), тоже относившийся к культурным сортам. Под обоими названиями долгое время в научной литературе фигурировали культурные сорта различного происхождения. По современным представлениям, большинство культурных сортов является результатом длительного отбора мутационных форм банана заостренного (М. acuminata, рис. 219, 1-6) и его скрещивания с бананом Балъбиса (М. balbisiana).
Банан заостренный - широко варьирующий вид, внутри которого выделяют 5 подвидов, легко скрещивающихся между собой. Вид распространен в Южной Индии, на полуострове Индокитай, полуострове Малакка, островах Малайского архипелага, на Новой Гвинее и в Северо-Восточной Австралии. В этом же географическом районе произрастает и банан Бальбиса, который продвигается несколько севернее в Индии до Ассама и в Южный Китай, но не растет в Австралии. Этот вид не обладает такой широкой изменчивостью. В тропической Азии известны естественные межвидовые гибриды этих видов. Оба они имеют гаплоидный набор из 11 хромосом. Культурные сорта большей частью являются триплоидами и вследствие этого не способны к половому размножению. Селекционерами принято условное обозначение набора хромосом банана заостренного латинской буквой А, а банана Бальбиса - латинской буквой В. Диплоидные сорта негибридного происхождения, созданные отбором форм банана заостренного, обозначаются шифром А А. Эти малоурожайные, неустойчивые к заболеваниям сорта имеют ограниченное распространение. Их оттеснили триплоидные сорта с генотипом AAA. К таким сортам относится известный высокорослый сорт "Гро Мишель" ("Cros Michel"). Одно его соплодие может содержать 250 плодов, масса каждого из которых до 200 г. Другую триплоидную мутацию банана заостренного представляет собой популярный карликовый сорт "Дворф Кавендиш" ("Dwarf Cavendish"). Он был распространен в культуре в Южном Китае и поэтому был известен как "китайский банан" (M. chinensis) или "карликовый банан" (M. папа), либо, наконец, как "банан Кавендиша" (M. cavendishii). Высота растений этого сорта около 1 м. Он быстро растет и плодоносит при выращивании в оранжереях ботанических садов. Описанный Линнеем банан райский представляет собой триплоидный гибридный сорт. Среди таких триплоидных гибридов с генотипом ААВ известна большая группа сортов, так называемых плантенов, выращиваемых главным образом в Центральной Африке. Плантены - овощные сорта, плоды которых не используют сырыми. Их пекут в банановых листьях, варят, перерабатывают на муку.
В Индии и странах Юго-Восточной Азии распространены сорта с геномом ABB. Современная селекция направлена на создание устойчивых к грибковым заболеваниям урожайных сортов, получены тетраплоидные гибриды.
Плоды столовых сортов содержат около 75% воды, 22% Сахаров, 1,3% белков и около 10 мг/% витаминов. Они представляют собой ценный диетический фрукт. Особая группа мучнистых сортов происходит от банана Маклая, или океанийского (M. maclayi, или M. fehi, рис. 219, 7), распространенного на островах Океании и в Австралии. Это овощные сорта с оранжевыми плодами, мякоть которых желтого цвета, семена их тоже используются в пищу.
Важной технической культурой тропиков является банан текстильный (М. textilis, табл. 46, 4), происходящий с Филиппинских островов, где его называют "абака". Прочное волокно из его расщепленных влагалищ известно как "манильская пенька". Оно устойчиво к гниению и является ценным сырьем для производства канатов и технических тканей. Подобное же применение находит банан японский (М. basjoo, табл. 46, 3), из которого в Японии изготовляют различные плетеные изделия.
Этот вид выдерживает кратковременные понижения температуры до -8° С, теряя листья и отрастая вновь от корневища, его успешно выращивают на Черноморском побережье Кавказа в качестве декоративного растения.
Энсета вздутая, или "абиссинский банан" (Е. ventricosum, рис. 219, 8-10), растет в тропической Африке во влажных речных долинах, на болотистых местах и лесных полянах, используется в пищу в качестве овощного растения. Молодые стебли и соцветия пекут, молодые плоды маринуют или едят свежими.
Семейство геликониевые (Heliconiaceae)
Это тропическое семейство включает всего лишь один род геликония (Heliconia, рис. 220, табл. 47 и 48) со 150 видами, распространенными главным образом во влажных тропиках Центральной и Южной Америки. Лишь один вариабельный вид - геликония индийская (Н. indica) произрастает в восточном полушарии на островах Океании.

Таблица 47. Геликониевые, имбирные и костусовые: 1 - геликония попугайная (Heliconia psittacorum), Сингапурский ботанический сад; 2 - геликония банановая (Н. bananae), Шри-Ланка, сад в Канди; 3 - гедихиум красивый (Hedychium speciosum), оранжереи Ленинградского государственного университета; 4 - альпиния зерумбет (Alpinia zerumbet), о. Маврикий, ботанический сад; 5 - костус мексиканский (Costus mexicanus), оранжереи Ботанического института АН СССР в Ленинграде

Таблица 48. Геликониевые: 1 - геликония металлическая (Heliconia metaliica); 2 - геликония Вагнера (Н. wagneriana), растение с плодами; 3 - геликония Коллинса (Н. collinsiana), соцветие, опыляемое колибри; видны завязавшиеся плоды
Геликонии - крупные многолетние травянистые растения, по облику напоминающие бананы. Они имеют симподиально нарастающие корневища, укороченные надземные стебли с крупными листьями, влагалища которых образуют ложные стебли. Растения некоторых видов невысоки. Так, у геликонии густоцветковой (Н. densiflora) высота ложного стебля всего 60 см. В то же время у многих видов они вырастают до 2-5 м, а у геликонии Марии (Н. mariae) достигают 8-10 м. Листья геликоний внешне похожи на листья бананов. Они могут достигать у высокорослых видов в длину 3 м и в ширину 1 м. Так же как у бананов, они несколько асимметричны из-за развития в тесной полости ложного стебля и имеют такое же жилкование. Отличает геликонии от банановых двурядное расположение листьев. У некоторых видов, например у геликонии жестковолосистой (Н. hirsuta), двурядность ложная, возникающая вторично при разрастании листовых влагалищ, которые в почке стебля закладываются по спирали. Переход от спирального расположения чешуевидных листьев на корневище и листовых зачатков в конусе нарастания надземного стебля к двурядному размещению листьев на взрослом растении можно проследить также у геликонии попугайной (Н. psittacorum, рис. 220, 8, табл. 47, 1).
Пазушные почки у основания надземного стебля дают начало новым столонам корневища. У геликонии повислой (Н. pendula), геликонии низкой (Н. humilis) и других видов растущее корневище почти сразу загибается к поверхности почвы (рис. 220) и развивает надземный стебель в тесном соседстве с материнским, что приводит к образованию очень плотных куртин. У геликонии ростральной (Н. rostrata, рис. 220, 6) нарастание новых стеблей происходит лишь в одну сторону, линейно. У геликонии Вагнера (Н. wagneriana, рис. 220, 7, табл. 48, 2) нет подземного корневища, она образует новые стебли из почек в нижней части стебля над поверхностью почвы. В то же время многие виды, например геликония попугайная, геликония прямая (Н. stricta), имеют довольно длинные горизонтальные корневища, образующие новые стебли на некотором расстоянии друг от друга. Такие виды с более рыхлыми куртинами быстрее заселяют пространство. У геликонии широкопокровной (Н. latispatha, рис. 220, 9), растущей часто на горных склонах, корневище стелется по поверхности земли.
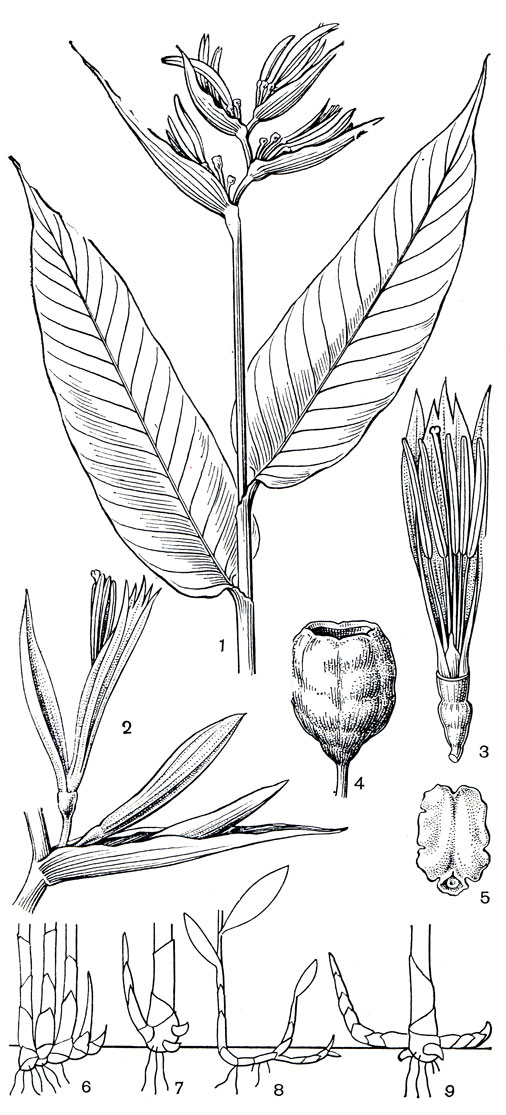
Рис. 220. Геликониевые. Геликония канновидная (Heliconia cannoidea): 1 - цветущее растение; 2 - парциальное соцветие; 3 - цветок, свободный чашелистик удален, видны столбик, 5 тычинок, стаминодий; 4 - плод; 5 - семя. Схематический рисунок строения корневищ: 6 - геликония ростральная (Н. rostrata); 7 - геликония Вагнера (Н. wagneriana); 8 - геликония попугайная (Н. psittacorum); 9 - геликония широкопокровная (Н. lаtispatha)
Геликонии растут быстро и зацветают обычно на второй год. Стебель, остававшийся укороченным в вегетативной фазе, при переходе к цветению быстро растет внутри ложного стебля, вынося наверх соцветие, которое у одних видов стоит вертикально, а у других принимает горизонтальное положение или повисает вниз. Ось соцветия несет двурядно расположенные боковые парциальные соцветия типа завитка, заключенные, как у стрелитциевых, в крупные ладьевидные заостренные на вершине кроющие листья. Они чаще всего необычайно яркой окраски - желтые, оранжевые, розовые, красные. Иногда у края их цвет переходит в другой, контрастный. Например, красные кроющие листья могут иметь зеленые или желтые края.
У низкорослых геликоний соцветие бывает обычно длиной около 10-30 см и несет всего 4-5 кроющих листьев, как у геликонии попугайной. Крупные геликонии нередко имеют соцветия длиной 1 -1,5 м. Соцветие геликонии короткопокровной (Н. curtispatha) содержит 25-40 кроющих листьев и достигает в длину 2,5 м, а у геликонии Марии в соцветии 40- 70 ярко-розовых кроющих листьев. Парциальные соцветия геликоний могут состоять всего из двух цветков и иметь небольшие кроющие листья длиной 6-8 см. У геликонии Вагнера, геликонии широкопокровной и других видов в одном кроющем листе последовательно развивается до 20 цветков. Кроющие листья, содержащие такие многоцветковые завитки, имеют длину 20-25 см. Соцветие этих видов содержит в целом до 250 цветков, а у геликонии Марии число их достигает 900. Цветение такого соцветия длится более полугода. У большинства видов соцветие функционирует 3-4 месяца, цветение начинается с нижних парциальных соцветий, от их основания, продвигаясь к вершине кроющих листьев.
Цветки геликоний обоеполые, зигоморфные, у основания снабжены прицветничками. Сегменты околоцветника, расположенные по 3 в 2 круга, обычно лепестковидно окрашены, белые, желтые, оранжевые, розовые или красные. У геликонии попугайной оранжевые сегменты околоцветника имеют черное пятно в верхней части. Три сегмента внутреннего круга, т. е. лепестки, примерно на две трети своей длины срастаются в трехзубчатый орган, свернутый в трубку (рис. 220, 3). Смыкающиеся его края находят друг на друга, но не срастаются. Снаружи к трубке прирастают два узких линейных чашелистика. Третий, довольно широкий чашелистик остается свободным и плотно примыкает к трубке, замыкая снаружи ее свернутые края. Внутри трубки 5 фертильных тычинок с линейными пыльниками, открывающимися продольной щелью. Шестая тычинка, расположенная у свободного чашелистика, стерильна и превращена в стаминодий, который, словно дополнительный клапан, создает герметичность трубки цветка.
Гинецей синкарпный, из трех плодолистиков, столбик нитевидный, с головчатым рыльцем, завязь нижняя, 3-гнездная. В каждом гнезде завязи, в основании, по 1 анатропному семязачатку. В ткань верхней части завязи погружены септальные нектарники, выделяющие нектар из 3 пор у основания столбика.
Цветки геликоний раскрываются рано утром и функционируют в течение одного дня. Они протандричны; пыльники вскрываются сразу, и пыльца, высыпаясь, попадает на рыльце, которое еще не начало функционировать. Пыльцевые зерна крупные. Они объединяются в хлопья благодаря игловидным кристаллам, образующимся в гнездах пыльников. После высыпания пыльцы столбик продолжает удлиняться. Бразильский систематик Умберто де Соуза Баррейрос (1974), изучающий геликонии, отмечает наличие у некоторых видов гетеростилии, которая помогает избежать само-опыления. Однако многие геликонии и при самоопылении способны образовывать полноценные семена.
Геликонии образуют обширные заросли в дождевых тропических лесах на опушках, открытых полянах, вырубках, по берегам рек и ручьев, а также по низким морским побережьям, у лагун. Лишь немногие виды могут выживать в глубине тенистых лесов.
Яркие, богатые нектаром соцветия геликоний не испытывают недостатка в опылителях. Их посещают черные пчелы из рода тригона (Trigona) и многочисленные птицы. Биологи, наблюдавшие в природе цветение геликоний, с восторгом описывают незабываемое красочное зрелище - яркие соцветия и порхающие около них пестрые тропические птицы. Возможно, птиц привлекает не только нектар, но и живущие в соцветиях насекомые и их личинки. Особенно богатая фауна в прямостоячих соцветиях геликоний, в кроющих листьях которых накапливается дождевая вода. В ней разлагаются опадающие околоцветники, поселяются простейшие, кишат личинки москитов. Р. П. Зайферт (1975) насчитал в соцветии геликонии Вагнера в Коста-Рике 25 видов насекомых, среди которых было особенно много личинок мух и жуков, питающихся растительными остатками.
Американский ботаник А. Ф. Скатч (1933) проследил развитие цветков геликонии Марии и геликонии Бихаи (Н. bihai), протекающее под водой, постоянно стоящей в кроющих листьях. Он установил, что погруженные вводу бутоны так герметично закрыты, что вода не проникает в них. Ночью, накануне раскрытия цветка, цветоножка усиленно растет и поднимает трубку цветка на 2 см над водой. Свободный чашелистик отгибается, открывая до-ступ в трубку, полную нектара. Тот же чашелистик служит и посадочной площадкой для пчел, а колибри (Phoethornis longirostris), постоянно порхающие у геликоний, пьют нектар, вибрируя крыльями и повисая при этом в воздухе над цветками.
Подробно наблюдавший цветение геликоний в Коста-Рике Ф. Г. Стайлс (1975) установил специализированные связи видов геликонии с опылителями. Геликонии с короткой, длиной не более 33 мм, трубкой цветков (геликония Марии, геликония широкопокровная и геликония черепитчатая) опыляют преимущественно колибри с коротким прямым клювом. Цветки геликонии Вагнера с трубкой околоцветника длиной около 48 мм посещают колибри с длинным изогнутым клювом. Крупные колибри (Eutoxeres aquila) опыляют цветки только одного вида - геликонии бородоцветковой (Н. pogonantha). Часто встречаются на соцветиях геликоний и яркие птички танагры из отряда воробьиных. Они пьют воду из соцветий, лакомятся насекомыми и питаются плодами геликонии, причем нередко уносят их, способствуя распространению семян.
Плод геликонии - коробочка с тремя округлыми гранями. Сначала он довольно сочный, но, созревая, делается кожистым, сухим и, наконец, вскрывается по створкам, септицидно, или распадается на три дробных односемянных части. Семена без ариллуса, уплощенные, овально-удлиненные, с волнисто выемчатыми краями, покрыты твердой оболочкой. Прямой зародыш, окруженный хорошо развитым эндоспермом, ориентирован продольно. Микропиле, хорошо заметное на семени, прикрыто круглой крышечкой, которую корень зародыша выталкивает при прорастании.
У видов с прямостоячими соцветиями плоды созревают при полном погружении и, только созрев, поднимаются на растущей плодоножке над водой. У многих видов плоды становятся ярко-синими и особенно заметными в красных или желтых кроющих листьях соцветия.
Геликонии с их орнаментальными листьями и необычайно яркими соцветиями стали украшением садов в тропиках и оранжерейных коллекций Европы. Особенно часто в ботанических садах выращивают геликонию металлическую (Н. metallica, табл. 48, 1), геликонию Бихаи и геликонию индийскую (Н. indica). К последнему виду преимущественно относятся пестролистные культурные садовые формы с белыми, желтоватыми или розовыми жилками, появившиеся в культуре главным образом в Малайзии и на островах Малайского архипелага, откуда их завозили в конце прошлого века в оранжереи Европы.
Семейство ловиевые (Lowiaceae)
Семейство ловиевые включает всего один род орхиданта (Orchidantha, рис. 221) с 7 или 8 видами, распространенными в Южном Китае, на полуостровах Индокитай и Малакка, а также на острове Калимантан.
Виды орхиданты - многолетние травы высотой 0,3-1,5 м, с симподиально ветвящимся корневищем и довольно толстыми корнями. Стебли сильно укорочены, иногда ветвятся у основания и несут двурядно расположенные листья на длинных черешках с влагалищами, охватывающими друг друга. Пластинка удлиненно-эллиптическая, ланцетовидная или почти линейная, с развитой средней жилкой и почти параллельными боковыми жилками. У орхиданты бахромчатой (О. fimbriata) длина пластинки листа достигает 1 м, у других видов обычно не превышает 30-50 см.
Цветки орхиданты обоеполые, зигоморфные, по облику напоминают цветки орхидных. Развиваются они в верхушечных соцветиях на коротких облиственных стеблях. Иногда листья под соцветием отмирают и стебель обнажается. Соцветие состоит из нескольких серий симподиально нарастающих элементов, по строению близких к завитку. Каждый элемент соцветия несет в основании чешуевидный лист с двумя килевидными выступами, затем два кроющих листа и на конце - цветок, завязь которого одета прицветничком. Кроющие листья соцветия зеленые или пурпурные, в основании трубчатые, несут в пазухе почки, из них последовательно развиваются такие же ветви соцветия, каждая из которых заканчивается цвет
ком. Первой всегда пробуждается почка верхнего кроющего листа, находящегося непосредственно под цветком, и дает начало целой системе монохазиальных элементов. Почка нижнего кроющего листа развивается значительно позже и образует новую серию монохазиев. Соцветие функционирует долго. По следам от опавших цветков и кроющих листьев можно проследить его историю. Появляются цветки с интервалом в 10-14 дней, причем каждый раскрывается на 1-2 дня.
Цветки орхиданты 3-членные, сегменты околоцветника дифференцированы на чашелистики и лепестки. В верхней части они свободные, а внизу образуют массивную трубку, сросшуюся с очень длинной (8-10 и даже до 20 см) нижней завязью (рис. 221, 1). Чашелистики линейные или ланцетные, у основания иногда темно-пурпурные или коричневатые. Лепестки очень неравные, 2 маленьких ланцетовидных располагаются по сторонам от большого, с овальной отогнутой пластинкой. Этот лепесток - самая заметная часть цветка, губа, на которую садятся насекомые. Лепестки чаще кремовые или белые, нередко с пурпурными пятнами, иногда коричневые или фиолетовые. Губа обычно отличается по окраске от остальных элементов цветка, иногда она испещрена мелкими пятнами или имеет темное пятно у зева. Пластинка ее может быть тупой или заостренной, обычно она несколько гофрированная, иногда разделена на три лопасти. Основание губы трубчато свернуто вокруг столбика и 5 тычинок. Тычинки с крупными пыльниками, открывающимися продольной щелью, тесно сгруппированы у зева трубки напротив губы. Шестая тычинка, место которой у основания губы, полностью редуцирована или присутствует в виде стаминодия, как в цветках орхиданты сиамской (О. siamensis).
Гинецей ловиевых синкарпный, из 3 плодолистиков, завязь 3-гнездная, с многочисленными анатропными семязачатками. Столбик несет 3-лопастное рыльце с изрезанными бахромчатыми краями. Лопасти развернуты в одной плоскости узким веером, воспринимающей поверхностью рыльце обращено к губе. Оно нависает над входом в цветочную трубку, готовое встретить опылителя.
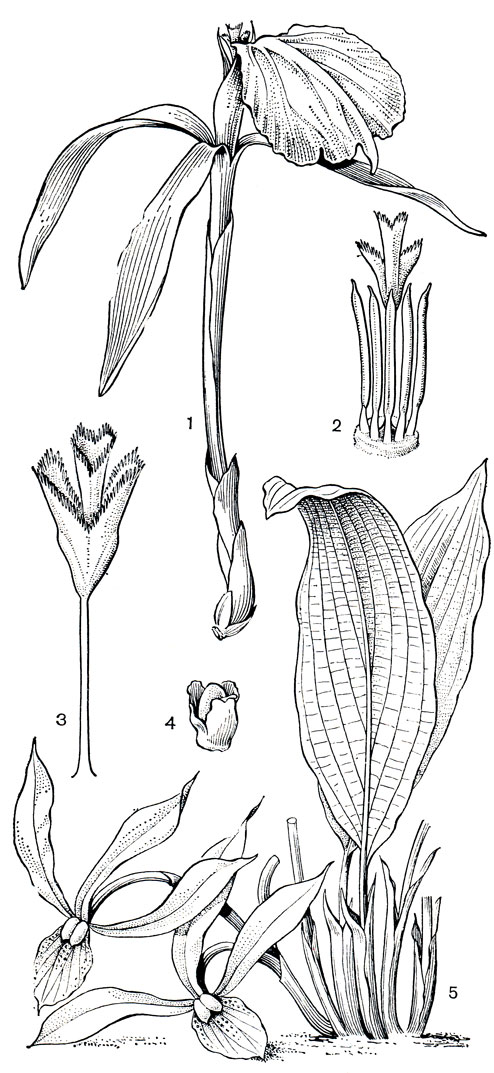
Рис. 221. Ловиевые. Орхиданта длинноцветковая (Orchidantha longiflora): 1 - цветок; 2 - тычинки и рыльце со спинной стороны; 3 - столбик и рыльце с воспринимающей поверхностью. Орхиданта максиллар невидная (О. maxillarioides): 4 - семя; 5 - цветущее растение
По сообщению известного английского ботаника Р. Э. Холттума (1970), цветки орхиданты бахромчатой издают неприятный запах, напоминающий запах клопов. В Сингапурском ботаническом саду он наблюдал на них многочисленных мух, которые, однако, не опыляли цветки, так как после посещения цветков мухами плоды не образовывались. Губа цветка, блестящая от слизистых выделений, через 6-8 ч вянет и быстро отмирает. Самостерильность цветков орхиданты свидетельствует о перекрестном опылении их в природе, однако этот процесс еще недостаточно изучен.
Плоды орхиданты - локулицидные коробочки цилиндрической формы - содержат многочисленные эллиптические семена с 3-лопастным ариллусом. Способ распространения семян в природе не изучен, но можно предположить, что здесь определенную роль играет мирмекохория. Орхиданта - лесное растение, ее виды отличаются теневыносливостью и приурочены к сырым тенистым местообитаниям во влажных тропических лесах. Крупные, высотой до 1,5 м, растения орхиданты длинноцветковой (О. longiflora) образуют местами заросли в лесах Малайзии, другие виды встречаются рассеянно, отдельными экземплярами, чаще всего их находят в вегетативном состоянии.
Виды орхиданты пока еще редки в коллекциях ботанических садов и являются интересным объектом исследования.
Семейство имбирные (Zingiberaceae)
Имбирные, насчитывающие около 47 родов и более 1000 видов, произрастают главным образом в лесах Южной и Юго-Восточной Азии, на островах Малайского архипелага и на Новой Гвинее. Лишь немногие виды алъпинии (Alpinia, табл. 47, 4), амомума (Amomum), имбиря (Zingiber, табл. 49, 3-4) и куркумы (Curcuma) встречаются в Северо-Восточной Австралии. В тропической Америке имбирные представлены 50 видами единственного рода ренеалъмия (Renealmia). В тропической Африке распространены виды эндемичных родов афрамомум (Aframomum) и ценковскиелла (Cienkowskiella). В лесах на западе континента произрастают около 25 видов ренеальмии и 1 вид аулотандры (Aulotandra), остальные 5 видов которой - обитатели Мадагаскара. Вне тропиков встречаются лишь отдельные виды имбирных. Самого северного положения около 30° с. ш. достигают они в Японии на острове Кюсю, где в лесах растут алъпиния японская (Alpinia japonica) и имбирь японский (Zingiber mioga). Примерно до такой же широты распространены имбирные в Китае. В южном полушарии имбирные также достигают 30°. В Южной Африке встречается несколько видов ценковскиеллы, а на юге Бразилии - виды ренеальмии.
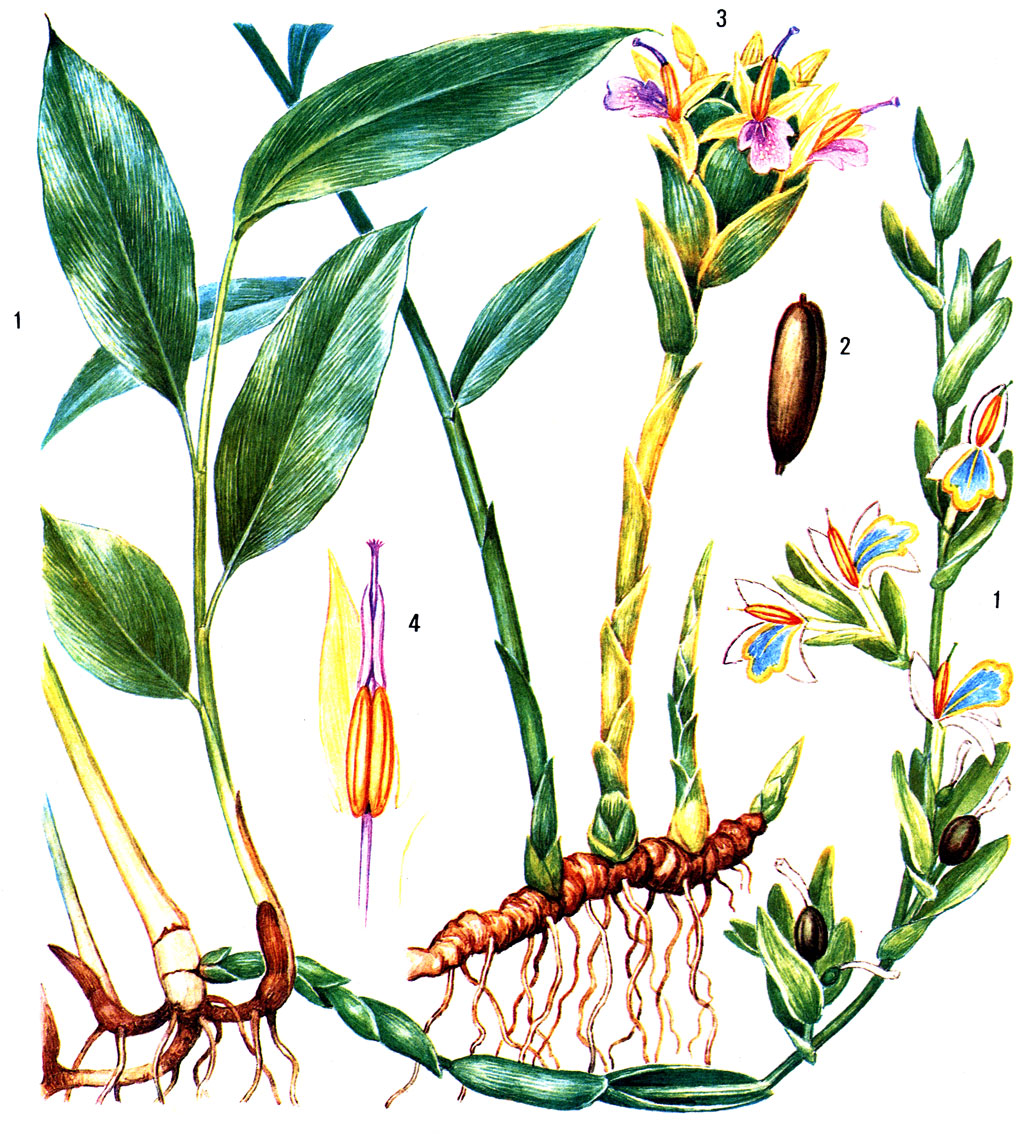
Таблица 49. Имбирные. Кардамон настоящий (Elettaria cardamomum): 1 - цветущее растение; 2 - плод. Имбирь аптечный (Zingiber officinale): 3 - цветущее растение; 4 - лепесток, тычинка с пыльниками и столбик, обернутый надсвязником
Имбирные - характерный элемент тропических дождевых лесов, где они образуют нередко сплошные заросли в густой тени, на полянах, по берегам рек и ручьев, на болотистых и периодически затопляемых низинах. В листопадных муссонных лесах Юго-Восточной Азии они образуют также пышный травяной покров. Некоторые виды ренеальмии, гедихиума (Hedychium, табл. 47, 3) и других родов являются эпифитами. Мощные двухметровые растения бэрбиджеи узкоцветковой (Bnrbidgea stenantha) растут в диптерокарповых лесах Калимантана на стволах деревьев в нескольких метрах от земли. Хотя среди имбирных преобладают лесные растения, некоторые из них встречаются и в саваннах. К таким растениям относится афрамомум бело-фиолетовый (Aframomum albo-violaceum), широко распространенный в африканских саваннах по обе стороны от экватора, где он переносит ежегодные пожары. Нередки имбирные во вторичных формациях на местах вырубок, на заброшенных плантациях. Как правило, они обитают на небольших высотах над уровнем моря, но некоторые виды произрастают высоко в горах. Стадиохилус бирманский (Stadiochilus birmanicus) массово растет на деревьях и скалах в горных лесах Бирмы на высоте 1700 м, а местами даже до 2600-3000 м над уровнем моря. Этой же высоты достигают в Гималаях виды родов роскоя (Roscoea), котлия (Cautleya) и каулокемпферия (Caulokaempferia), а в горах Колумбии ренеалъмия высотная (Renealmia alticola).
Имбирные - многолетние корневищные растения, у которых все вегетативные органы и семена содержат эфирные масла со специфическим ароматом. Корневища имбирных обычно довольно толстые или клубневидные, покрыты двурядно расположенными чешуевидными влагалищными листьями и несут на каждом междоузлии придаточные корни. У некоторых видов роскои, глоббы (Globba), куркумы, ганъепении (Gagnepainia) корни клубневидно утолщены. Корневище нарастает симподиально. Каждая растущая его ветвь, сформировав несколько междоузлий, начинает расти к поверхности почвы и образует надземный облиственный стебель (рис. 222). Нередко еще до окончания роста стебля у его основания на корневище в пазухе чешуевидного листа пробуждается почка, дающая начало новой части корневища. Число междоузлий корневища, вырастающих до образования стебля, постоянно для каждого вида и обычно невелико. Благодаря тому что междоузлия короткие, имбирные образуют довольно густые куртины. Необычное корневище, приподнятое на придаточных корнях над поверхностью почвы, имеет ренеалъмия Елены (Renealmia helenae), растущая в болотистых лесах Панамы. У хорнштедтии большой (Hornstedtia grandis) из лесов Малайзии мощные корневища поднимаются на полметра над землей на толстых красноватых ходульных корнях. Наличие мясистых корневищ, в которых запасаются вода и питательные вещества, позволяет многим имбирным пере-живать засушливый период и обеспечивает процветание этого семейства в области муссонного климата. Виды, растущие в постоянно влажном климате, обнаруживают тенденцию к непрерывному росту, но у обитателей муссонных лесов и саванн существует период покоя, когда надземные части отмирают. Контрактильные корни, которые имеются у некоторых видов, втягивают корневище поглубже в почву на время засухи. Кемпферия округлая (Kaempferia rotunda), культивируемая в Юго-Восточной Азии, сохраняет ежегодный период покоя даже при выращивании в условиях постоянной влажности. Такой же генетически закрепленный период покоя имеют виды афрамомума из саванн Африки, глобба марантовая (Globba marantina), обитающая в северной части полуострова Малакка, и многие другие.
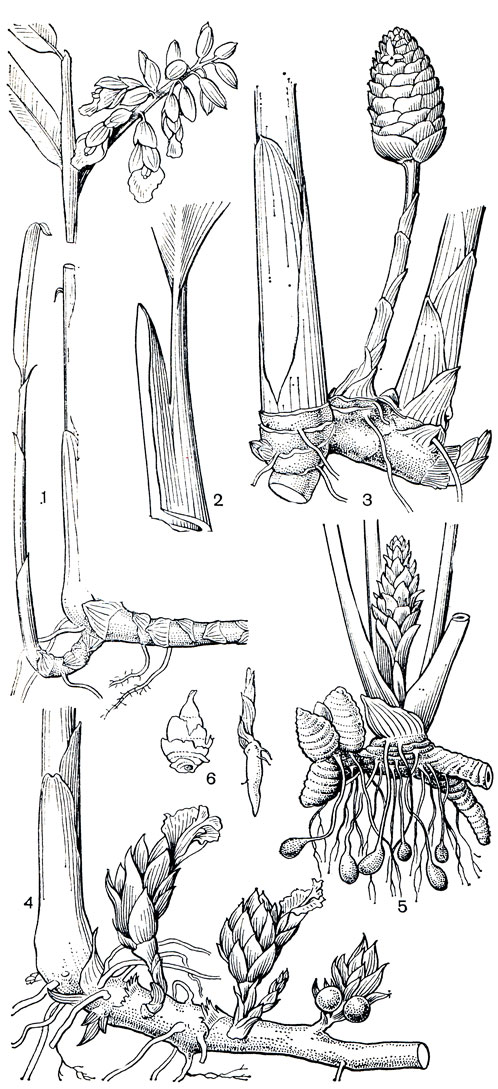
Рис. 222. Имбирные. Альпиния тупоконечная (Alpinia mutica): 1 - цветущее растение; 2 - черешок с влагалищем. Имбирь зерумбет (Zingiber zerumbet): 3 - корневище с цветоносным стеблем. Амомум розовый (Amomum roseum): 4 - корневище с соцветиями и плодами на коротких генеративных стеблях. Куркума домашняя (Curcuma domestica): 5 - корневище с клубнями и клубневидными корнями, несущее стебель с соцветием; 6 - бульбиллы, образующиеся в соцветиях: слева - глоббы поникшей (Globba cernua); справа - глоббы ломкой (G. fragilis)
Стебель надземного побега в вегетативной фазе остается коротким и утолщенным, с сильно сближенными междоузлиями. На нем тесно двумя рядами располагаются очередные листья с длинными, обычно незамкнутыми влагалищами. Лишь у некоторых видов роскои и у котлии изящной (Cautleya gracilis) листовые влагалища трубчатые. Влагалище охватывает стебель, края его находят друг на друга, образуя трубку, внутри которой развивается следующий лист. Так же как у бананов, у имбирных развиваются из вложенных друг в друга влагалищ полые ложные стебли, которые могут достигать большой высоты. На Соломоновых островах известно несколько гигантских альпиний, высотой до 8 м, а алъпиния односторонняя (Alpinia unilateralis) достигает в высоту даже 10 м. В то же время у аулотандры мадагаскарской (Aulotandra madagascariensis) очень короткие стебли несут всего по одному листу, а у цифостигмы двулистной (Cyphostigma diphyllum) - по два. На первый взгляд кажется, что листья растут прямо на ползучем корневище, настолько незаметны их стебли.
Число листьев на стебле довольно постоянно у каждого вида имбирных и может служить систематическим признаком. У глоббы, гедихиума и родственных им родов ряды листьев на надземных стеблях точно продолжают расположение рядов чешуевидных листьев на корневищах. У альпинии и близких к ней родов расположение листьев на надземных стеблях поперечное, повернутое на 180° по отношению к рядам чешуй на корневище. Первые листья надземного побега не имеют развитой пластинки, так что основание ложного стебля обернуто снаружи несколькими влагалищными листьями - катафиллами. Листья имбирных часто сидячие или имеют небольшой черешок, верхний край влагалища всегда с хорошо развитым язычком - лигулой. Американский ботаник П. Б. Томлинсон (1956) обнаружил у имбиря, в месте прикрепления пластинки, утолщение на черешке, подобное "подушечке" марантовых. Пластинки листьев у имбирных чаще ланцетовидно удлиненные, овальные, иногда широкоовальные или почти округлые, у некоторых видов линейные, всегда с хорошо развитой главной жилкой и почти параллельными боковыми жилками. Поскольку лист развивается в трубке ложного стебля, он несколько асимметричный. Листья некоторых имбирных имеют опушение, особенно часто на лигуле, по краям и вдоль главной жилки, а иногда и по всей поверхности. Волоски опушения обычно одноклеточные, с основанием, погруженным в эпидермальный слой, в отличие от поверхностных волосков костусовых. Проводящая система имбирных состоит почти исключительно из трахеид, сосуды найдены лишь в стеблях ренеальмии, а у остальных предста-вителей семейства - в корнях.
Цветки имбирных в верхушечных соцветиях. При переходе к цветению побег быстро растет внутри ложного стебля и выносит соцветие над листьями. Только у плагиостахиса (Plagiostachys) из лесов Малайзии соцветие остается на укороченном стебле, не поднимается кверху во влагалищной трубке, а прорывает ее сбоку и появляется наружу у основания ложного стебля. У николайи (Nicolaia), афрамомума, кардамона (Elettaria, табл. 49, 1-2) и многих других имбирных имеются стебли, несущие соцветия, лишенные развитых листьев и покрытые лишь чешуевидными катафиллами. Они образуются из почек на корневище у основания вегетативных стеблей и обычно значительно короче их. У некоторых видов афрамомума и ахасмы (Achasma) цветоносные стебли достигают всего нескольких сантиметров в длину и полностью погружены в землю вместе с нижней частью соцветий; на поверхности почвы видны только яркие цветки, а плоды развиваются под землей. Похожую картину можно наблюдать у видов кардамона и элеттариопсиса (Elettariopsis), обитающих в лесах Малайзии. Соцветия их стелются по земле или развиваются под слоем опавших листьев, над которыми поднимаются цветки.
Для имбирных характерно соцветие типа тирса и его различные модификации. Главная ось соцветия обычно несет спирально расположенные кроющие листья. Они могут быть зелеными или ярко окрашенными - желтыми, оранжевыми, красными, фиолетовыми - и тогда, несомненно, играют роль в привлечении опылителей. Изредка встречаются соцветия с белыми кроющими листьями, как у ренеальмии пирамидальной (Renealmia pyramidalis). У николайи высокой (Nicolaia elatior) с островов Малайского архипелага соцветие окружено снизу крупными ярко-красными кроющими листьями, которые образуют обертку и придают соцветию облик огромного цветка. Некоторые альпинии имеют очень крупные кроющие листья, у других они мелкие или вообще не развиты, а у глоббы они быстро опадают. В то же время у растений с погруженными в землю соцветиями крупные и кожистые кроющие листья играют защитную роль. В пазухах кроющих листьев расположены парциальные соцветия - завитки. Завиток состоит из серии последовательно развивающихся осей, каждая из которых заканчивается цветком и несет под ним прицветник. У видов риделии (Riedelia), ганьепении и гемиорхиса (Hemiorchis) прицветники не развиты. Прицветники могут быть ланцетными или иметь трубчатое строение; обычно они тоже ярко окрашены и несут в пазухе почку, дающую начало новой веточке завитка. Так образуются тирсы с многоцветковыми завитками, как у многих видов ренеальмии, альпинии, глоббы и др.
Укорочение осей завитка приводит к образованию соцветий колосовидного облика, как, например, у куркумы (рис. 222). Плотные кожистые кроющие листья куркумы спиральными рядами густо располагаются на довольно мясистой оси; верхние из них сближены, не несут парциальных соцветий и иногда ярко окрашены в розовый или красный цвет. Края кроющих листьев куркумы несколько свернуты и почти до половины срастаются с соседними. В их пазухах, как и в соцветиях геликоний, накапливается дождевая вода, и развитие цветков происходит в воде, где разлагаются части отцветших цветков. В воде же созревают и затем размокают плоды, а семена выскальзы-вают из слизистой массы.
В процессе еще большего укорочения главной оси соцветия образовались плотные головчатые соцветия, как у афрамомума, хорнштедтии и других родов. Иногда в парциальном соцветии имбирных происходит редукция числа цветков до одного, и соцветие приобретает облик кистевидного, колосовидного или головчатого. Одноцветковые парциальные соцветия имеются почти у всех видов имбиря, у многих видов ренеальмии, амомума, гедихиума, кемпферии. У некоторых видов кемпферии, афрамомума и других родов все соцветие редуцировано до единственного цветка. Примерами могут служить кемпферия сиккимская (Kaempferia sikkimensis) или роскоя альпийская (Roscoea alpina), которую нередко выращивают в ботанических садах.
Цветки имбирных почти всегда обоеполые, зигоморфные, обычно душистые и яркие. Чашечка зеленая, трубчатая. Лепестки более или менее сросшиеся, неравные. Обращенный к оси соцветия (адаксиальный) лепесток, как правило, крупнее остальных и иногда капюшонообразно вогнут. У его основания расположена единственная фертильная тычинка, принадлежащая к внутреннему кругу андроцея, остальные 2 члена которого стерильны и срослись в лепестковидный стаминодий, называемый губой и имеющий у разных родов очень различную форму. Передняя (абаксиальная) тычинка внешнего круга всегда отсутствует, а остальные 2 или тоже отсутствуют, или превращены в маленькие или крупные лепестковидные стаминодии, располо-женные по обе стороны фертильной тычинки, а у имбиря - приросшие к губе. Они хорошо развиты у гедихиума, глоббы и родственных им родов и обычно отсутствуют у альпинии и других близких к ней имбирных. Внешний облик цветка определяет главным образом противостоящая фертильной тычинке лепестковидная губа. Основание губы трубчато свернуто и прикреплено к трубке венчика, а отогнутая пластинка цельная или 2-, 3-лопастная, яркая, иногда с желтым или темным пятном у зева. У многих видов ренеальмии, ринхантуса (Rhynchanthus) и других родов губа небольшая, вертикально стоящая и цветок имеет трубчатую форму.
Строение фертильной тычинки очень варьирует и определяет механизм опыления цветка. Тычиночная нить имбирных широкая и несет 2 одногнездных пыльника. Связник нередко продолжен над ними в крупный надсвязник самой различной формы (рис. 223, 7-9), иногда зубчатый или с выступающими лопастями. У имбиря аптечного (Zingiber officinale, табл. 49, 3, 4) надсвязник обернут вокруг длинного и нежного столбика и защищает его, оставляя свободным рыльце. Необычное строение имеет тычинка мантисии (Mantisia), у которой надсвязник расширен и имеет выступающие в стороны лопасти. Пыльники вскрываются продольно; пыльцевые зерна однобороздные или безапертурные (у альпинии, куркумы и других родов); оболочка их клейкая и поэтому они часто слипаются. Гинецей имбирных из 3 плодолистиков, обычно синкарпный, 3-гнездный, но некоторые виды кемпферии обнаруживают тенденцию к неполному развитию перегородок завязи; глобба, мантисия, ганьепения и гемиорхис имеют паракарпный гинецей и 1-гнездную завязь с постенной плацентацией. Завязь всегда нижняя, семязачатки анатропные или несколько изогнутые. Столбик тонкий, у некоторых видов погружен в желобок на задней стенке цветочной трубки. Вертикального положения столбик достигает лишь при поддержке тычинки, как и у костусовых; наверху он закреплен между пыльниками и выносит рыльце над ними. Рыльце обычно воронковидное, с железистым опушением по краям.
У основания столбика на вершине завязи расположены нектарные желёзки разнообразного строения (рис. 223, 10-14), которые, по мнению А. Кронквиста (1981), вероятно, представляют собой видоизмененные верхушки септальных нектарников. Иногда они неправильной формы, как у некоторых альпиний, или имеют вид двух цилиндрических образований. Чаще всего у имбирных 2 тонких шиловидных нектарника такого типа, как у роскои пурпурной (Roscoea purpurea, рис. 223,14). Выделяемый желёзками обильный нектар привлекает опылителей.
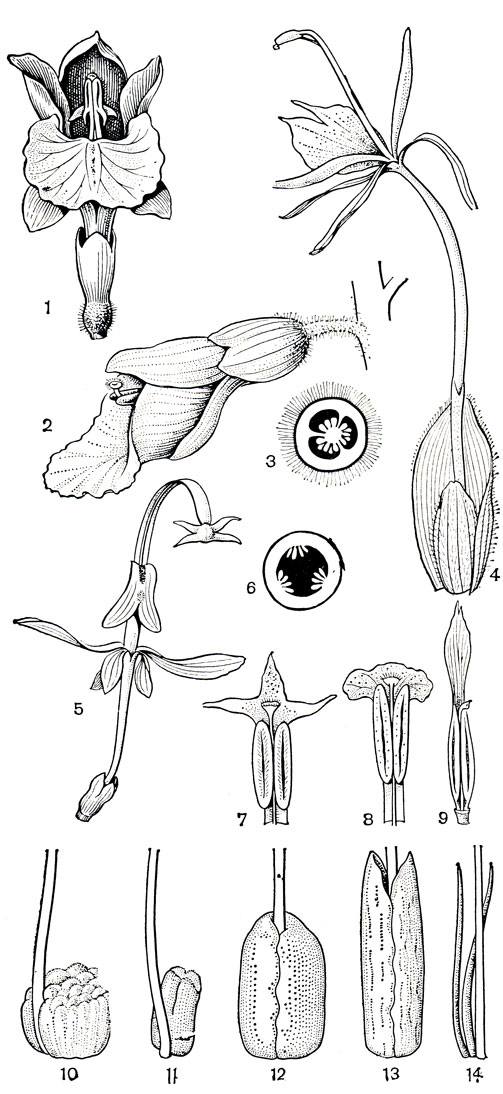
Рис. 223. Имбирные. Куркума австралазийская (Curcuma australasica): 1 - цветок. Альпиния зерумбет (Alpinia zerumbet): 2 - цветок; 3 - поперечный разрез завязи. Гедихиум цилиндрический (Hedychium cylindricum): 4 - цветок. Глобба поникшая (Globba cernua): 5 - цветок; 6 - поперечный разрез завязи. Тычинки с надсвязниками различного строения: 7 - вида амомума (Amomum sp.), 8 - амомума Кёнига (A. koenigii), 9 - бэрбиджеи блестящей (Burbidgea nitida). Нектарные железки, расположенные на завязи у основания столбика: 10 - вида альпинии (Alpinia sp.), 11 -гедихиума цилиндрического, 12 -ценолофона серебристого (Cenolophon argenteum), 13 - горнштедтии войлочной (Hornstedtia tomentosa), 14 - роскои пурпурной (Roscoea purpurea)
Цветки имбирных очень недолговечны. Они раскрываются утром всего на несколько часов и к вечеру уже увядают. В соцветии они зацветают последовательно снизу вверх. У большинства имбирных одновременно в соцветии открыто по одному-два цветка, редко больше. Для имбирных характерна протандрия.
Наблюдений за опылением цветков имбирных, к сожалению, немного. Р. Э. Холттум (1950) для имбирных Малайзии указывает в качестве опылителей дневных бабочек с длинными хоботками. На белых душистых цветках гедихиума венценосного (Hedychium coronarium) и других видов с длинными тычинками замечены бабочки-бражники (Sphingidae), которые высасывают нектар, паря перед цветком. Мало изучено опыление имбирных с цветками, находящимися у земли. Г. Н. Ридли (1899) наблюдал на цветках хорнштедтии мух и пчел, но все ли они на самом деле являются опылителями, неясно.
Голландский ботаник П. Мас (1977), подробно изучавший ренеальмии в тропической Америке, выделяет у них два типа опыления в зависимости от строения цветка. Виды ренеальмии с трубчатым строением цветка опыляют колибри; у других видов крупная отогнутая губа служит посадочной площадкой для пчел. Несомненно, что закономерность, отмеченная П. Масом для ренеальмии, распространяется и на имбирные восточного полушария. Известный исследователь биологии опыления П. Кнут указывает (1898) на птицеопыление соцветий николайи высокой. Однако, как показали наблюдения, большинство имбирных с крупной губой опыляют пчелы. Пробираясь за нектаром в трубку цветка, опылитель прежде всего касается торчащего над пыльниками рыльца и оставляет на нем принесенную пыльцу, а затем проходит под пыльниками и обсыпается пыльцой этого цветка. Особый механизм опыления имеется у цветков роскои, котлии, куркумы и ряда других родов, у которых нижняя часть пыльников стерильна и превращена в шпорцы. Особенно специализирован аппарат опыления у роскои (рис. 224), у которой пыльники подвижно прикреплены к нити в середине спинной стороны. При раскрытии цветка поднятые вверх боковые стаминодии загнутыми краями оттягивают назад рыльце и связник, а шпорцы выдвигаются вперед, как взведенный курок. Насекомое, пытающееся проникнуть за нектаром, невольно нажимает на шпорцы, и сложный механизм приходит в действие. Пыльники, как рычаг, наклоняются и вытряхивают пыльцу на спинку насекомого. Столбик, свободно двигающийся в желобке тычинки и между пыльниками, выдвигается, словно струна, вынося рыльце довольно далеко, и касается спинки насекомого за пределами зоны попадания своей пыльцы. К сожалению, неизвестно, какие насекомые опыляют цветки роскои в Гималаях. В ботаническом саду Бергена их опыляли крупные шмели. Директору сада Р. Нордхагену (1932) удалось заметить, что, когда шмель вылезает из цветка, волоски рыльца защищают его от повторного попадания пыльцы, обеспечивая таким образом перекрестное опыление
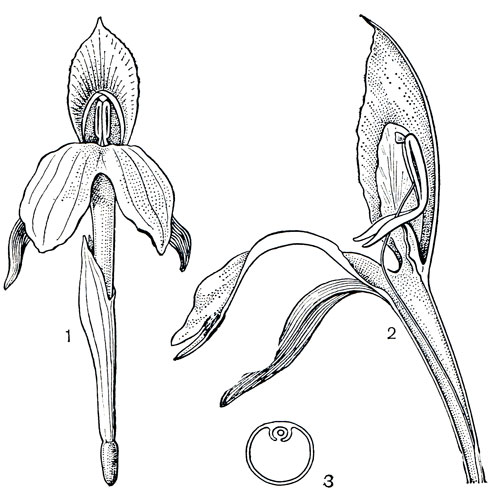
Рис. 224. Роскоя пурпурная (Roscoea purpurea): 1 - цветок; 2 - продольный разрез цветка, видны торчащие вперед шпорцы пыльников и выгнутый столбик; 3 - поперечный разрез цветочной трубки, видны выросты, образующие желобок, в котором фиксирован столбик
Для гедихиума и некоторых других родов отмечена самостерильность, но многие имбирные завязывают плоды и при самоопылении.
Плоды у имбирных двух типов: преобладают локулицидные коробочки, но встречаются и сочные нераскрывающиеся ягодообразные плоды, как, например, у афрамомума (рис. 225, 5), у некоторых видов альпинии (рис. 225, 3) и риделии. Семена чаще овальные или удлиненные с несколькими гранями, одеты твердой оболочкой и окутаны ариллусом, который обычно имеет неправильно изрезанные края. Ариллус по-видимому, играет роль при раскрывании плода. Р. Нордхаген наблюдал интенсивное раскрывание плодов роскои во влажную погоду, когда ариллусы сильно набухали. Створки плода роскои под давлением раскрываются и, выгибаясь, с силой выбрасывают семена на землю. Полное раскрытие плодов наблюдается также у гедихиума, брахихилума (Brachychilum), имбиря, глоббы. Однако не всегда семена разбрасываются. Благодаря слипающимся ариллусам (рис. 225, 4, 7) они иногда остаются лежать в раскрытом плоде или выпадают одним пакетом. Ярко-оранжевые коробочки брахихилума, раскрываясь, обнажают плотно сидящие семена с красным ариллусом. Сочные ароматные плоды и яркие семена привлекают птиц, активно участвующих в распространении семян имбирных. Плоды имбирных поедают обезьяны, кабаны, грызуны и другие животные.
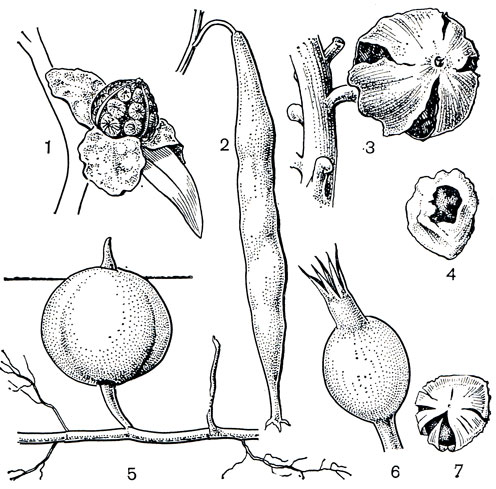
Рис. 225. Имбирные. Котлия желтая (Cautleya lutea): 1 - раскрывшийся плод. Силиквамомум тонкинский (Siliquamomum tonkinense): 2 - плод. Альпиния малаккская (Alpinia malaccensis): 3 - раскрывающийся плод; 4 - семя с ариллусом. Афрамомум атевский (Aframomum atewae): 5 - подземный плод на корневище. Альпиния галангa (Alpinia galanga): 6 - плод; 7 - семя с ариллусом
Большая роль в распространении семян имбирных принадлежит муравьям, которых привлекает сочный, маслянистый, а иногда и сладкий ариллус. В опыте, проведенном Р. Нордхагеном, муравьи за 5 мин утащили 30 семян роскои, почти не тронув семян колокольчика, служивших контролем.
Семена имбирных имеют прямой зародыш, окруженный остатком эндосперма, а запасающей тканью служит перисперм. В отличие от костусовых семена имбирных прорастают под землей.
Многие виды имбирных плодоносят редко. В их жизни большую роль играет вегетативное размножение, главным образом корневищами. Виды рода глобба из Юго-Восточной Азии имеют специальные органы вегетативного размножения - бульбиллы, образующиеся в пазухах нижних кроющих листьев на главной оси соцветия или в завитках наряду с цветками. Бульбиллы (обычно величиной 1-2 см) могут состоять из укороченного стебля, одетого чешуями (рис. 222, б), или из стебля и короткого толстого корня, а у глоббы Холттума (Globba holttumii) из Малайзии бульбиллы еще на соцветии начинают развивать листья и превращаются в готовые маленькие растеньица. Бульбиллы легко опадают при прикосновении, но, попав на землю, могут сохранять жизнеспособность в течение нескольких засушливых месяцев до начала муссонных дождей.
Семейство имбирные подразделяется на 3 трибы: глоббовые (Globbeae), имбирные (Zingibereae) и алъпиниевые (Alpineae). Глоббовые характеризуются одногнездной завязью; в этой трибе 4 рода и более 100 видов; самый большой род глобба (Globba) заключает около 100 видов, распространенных в Индии, Бирме, Южном Китае, на полуостровах Индокитай и Малакка, на островах Малайского архипелага, Филиппинах и Новой Гвинее.
Две другие трибы характеризуются 3-гнездной завязью. Представители трибы имбирных (18 родов и около 300 видов) имеют лепестковидные боковые стаминодии (у имбиря приросшие к губе) и густые соцветия. Растения трибы альпиниевых (25 родов и около 600 видов) имеют мелкие боковые стаминодии или совсем лишены их, им присущи менее густые соцветия. Самый большой род алъпиния (Alpinia, табл, 47, 4) заключает свыше 200 видов, распространенных в тропической Азии, на островах Малайского архипелага, Филиппинах, Новой Гвинее и в Полинезии.
Благодаря содержанию эфирных масел многие имбирные издавна используют как пряные и лекарственные растения. Одним из важнейших пряных растений является имбирь аптечный, происходящий из Южной Азии. Его с древних времен культивировали в Индии, а в настоящее время выращивают во всех тропических странах мира. Корневища имбиря, очищенные или только промытые и высушенные на солнце, содержат 2-3% эфирного масла, имеющего специфический аромат и жгучий вкус. Корневища имбиря применяют для ароматизации пищевых продуктов: кондитерских изделий, варенья, пряных соусов, ликеров, пива, а также используют в парфюмерии. Заменой настоящего имбиря служат корневища некоторых других видов этого рода: имбиря зерумбет (Zingiber zerumbet, рис. 222, 5), имбиря касумунар (Z. cassumunar) из тропической Азии и имбиря японского (Z. mioga).
Большое значение имеет кардамон настоящий (Elettaria cardamomum, табл. 49, 1-2), растущий во влажных горных лесах Южной Индии. Кардамон выращивают главным образом в Индии, на острове Шри-Ланка, полуострове Индокитай и в Южном Китае, закладывая плантации на высоте 600-1500 м над уровнем моря. Семена кардамона содержат 3,5-7% эфирного масла, их используют как пряную приправу к пище, в кондитерской и табачной промышленности, а также в медицине. Заменителем кардамона могут служить семена некоторых других имбирных, чаще всего амомума шиловидного,, так называемого "непальского кардамона" (Amomum subulatum), и алъпинии галанга, или галанги большей (Alpinia galanga). Альпиния галанга распространена в культуре по всей тропической Азии, ее корневища используют как приправу к рису и как источник крахмала. Альпиния лекарственная, или галанга меньшая (A. officinarum), содержит в корневищах 0,7-0,8% эфирного масла и распространена как пряное и лекарственное растение в странах тропической Азии. Корневища этого растения экспортируют в Европу.
Во всех тропических странах возделывают куркуму домашнюю, или культурную (Curcuma domestica), происходящую из Юго-Восточной Азии. В корневищах куркумы 1,3-5,5% эфирного масла, применяемого для ароматизации пищевых продуктов и в парфюмерии. Кроме того, из корневищ получают пищевой крахмал и стойкий желтый краситель для тканей, сливочного масла, сыров. Как пряное и лекарственное растение с древних времен в странах Юго-Восточной Азии культивируют также куркуму цитварную (С. zedoaria), а также кемпферию округлую, клубневидные корневища которой съедобны.
Многие виды имбирных используют как овощные растения и применяют в народной медицине. Из стеблей и листьев гедихиума венценосного и николайи высокой изготовляют плетеные изделия и особую бумагу. Альпинию зерумбет (Alpinia zerumbet), гедихиум венценосный, гедихиум гарднера (Hedychium gardnerianum), гедихиум ярко-красный (Н. coccineum) и другие виды имбирных выращивают в оранжереях ботанических садов как декоративные растения. Красиво цветущие виды роскои можно культивировать на юге СССР, укрывая корневища на зиму.
Семейство костусовые (Costaceae)
В пантропическом семействе костусовых 4 рода и около 200 видов, большинство которых распространены в Центральной и Южной Америке. Здесь представлены 3 рода: монотипный род монокостус (Monocostus), род димерокостус (Dimerocostus), в котором 2 вида, и костус (Costus, рис. 226, табл. 47, 5, 50, 4, 5), содержащий около 40 видов,- единственный пантропический род в семействе (число видов его убывает во флоре по мере распространения к востоку). В тропической Африке насчитывают около 25 его видов, в Юго-Восточной Азии - всего 5, а в лесах Северо-Восточной Австралии произрастает единственный вид - костус Потъе (С. роtierae). На Молуккских островах, в Новой Гвинее, на архипелаге Бисмарка и в лесах Квинсленда в Австралии распространено около 20 видов рода тапейнохилус (Tapeinochilus, табл. 50, 1, 3).

Таблица 50. Костусовые. Тапейнохилус колючий (Tapeinochilus pungens): 1 - соцветие; 2 - цветок; 3 - продольный разрез цветка. Костус длинноостроконечный (Costus cuspidatus): 4 - цветущее растение; 5 - тычинка с пыльниками, над ними - рыльце
Большинство костусовых растет куртинами, образуя почти чистые заросли во влажных тропических лесах, на полянах и вырубках, по берегам рек и ручьев, на болотах. К почве они неприхотливы и встречаются и на глинистых и на песчаных почвах. Среди африканских видов костуса есть эпифиты, например костус Ле-Тестю (Costus letestui) и костус голостебелъный (С. nudicaulis). Большую роль играют костусовые во вторичных формациях, на вырубках, плантациях, по обочинам дорог. В Центральной Америке на Атлантическом и Тихоокеанском побережьях костусовые местами образуют заросли вместе с геликониями, каннами, марантовыми и карлюдовикой. В то же время некоторые виды поднимаются в горах на большую высоту над уровнем моря. Так, в тенистых ущельях Коста-Рики костус горный (С. montanus) встречается на высоте 2000 м, а в Перу и Боливии на таком же уровне - димерокостус серебристый (Dimerocostus argenteus). Костусы произрастают и в светлых редколесьях, и в саваннах среди кустарников. В Центральном Перу по реке Уальяга в сухих лесах типа каатинги вместе с кактусами встречается монокостус одноцветковый (Monocostus uniflorus).
Костусовые, как правило, крупные или даже гигантские корневищные травы. Корневище симподиально ветвится в почве и состоит из отдельных частей, заканчивающихся надземным стеблем. Корневище тонкое или чаще клубневидно утолщенное, покрытое чешуевидными листьями и нередко опушенное, несет многочисленные придаточные корни. У костуса почти сидячего (Costus subsessilis), произрастающего в кампосах Бразилии, корни имеют на концах веретеновидные утолщения. Запас воды в корнях помогает этому растению с коротким корневищем и стеблем высотой всего 12-20 см пережить засушливый период. Обычно стебли костусовых хорошо развиты, высотой до 2-3 м, а у некоторых, например у костуса гладкого (С. laevis) из влажных тропических лесов Южной Америки, они высотой до 6 м. Листья у него, как правило, эллиптические, у костуса имбирного (С. zingiberoides) - линейные, злаковидные, а у костуса Малорта (С. malortieanus) - широкоэллиптические, почти округлые. Обычно листья почти без черешка, но с хорошо развитым замкнутым трубчатым влагалищем, которое охватывает стебель. Верхний край влагалища заканчивается лигулой и несет иногда реснитчатое опушение. Располагаются листья на стебле спирально, что сочетается у костуса с вращением оси стебля по мере роста.
Известный немецкий морфолог Карл Гёбель рассматривал стебель костуса как лиану, растущую без опоры. Действительно, стебель не стоит вертикально, а спирально изогнут, подобно винтовой лестнице, что обеспечивает максимально выгодную экспозицию листьев к свету в сумраке тропического леса. Стебли тапейнохилуса ветвятся. У костуса при повреждении точки роста из пазушных почек также развиваются боковые ветви. Интересно, что направление листовой спирали на ветвях обычно обратно направлению на главном стебле. Растения, выросшие из семян одного плода, разделяются примерно на равные группы с листовой спиралью, направленной по часовой стрелке и против нее.
Основание стебля, иногда высотой до 0,5 м, покрыто влагалищными листьями, не имеющими пластинок. У костуса длинноостроконечного (С. cuspidatus, рис. 226, 7-8, табл. 50, 4, 5) из лесов Южной Бразилии в пазухах верхних листьев образуются овальные бульбиллы длиной до 1 см, покрытые густым коричневым опушением. Опадая на землю, они могут переживать засушливый период и прорастать при благоприятных условиях. Все вегетативные органы костусовых нередко имеют опушение из мелких одноклеточных или длинных многоклеточных волосков. Костусовые не содержат в тканях эфирных масел и не имеют аромата.
Цветки костусовых обычно в колосовидных или головчатых соцветиях, венчающих облиственные стебли. У некоторых видов они появляются на специализированных коротких безлистных стеблях, покрытых лишь чешуевидными влагалищными листьями. Репродуктивные побеги вырастают у самого основания вегетативных стеблей, а иногда на значительном расстоянии от них, на другом конце корневища. Ботаники, собиравшие гербарий в лесных зарослях, не всегда могли сопоставить булавовидные, похожие на шишки соцветия с вегетативными стеблями, что подчас порождало путаницу при определении материала.
Сходство с шишкой соцветию придают многочисленные прицветники, плотно расположенные спиральными рядами. В пазухах прицветников расположены цветки, снабженные у основания трубчатым или ладьевидным прицветничком (рис. 226, 2, 3). Особенно крупные соцветия, диаметром до 25 см, имеют костус гладкий и костус гуанайский (С. guanaiensis). У большинства видов прицветники зеленые или желтоватые, у некоторых - ярко-оранжевые или красные. Очень яркие соцветия имеют костус красивый (С. speciosus) и тапейнохилус колючий (Tapeinochilus pungens, табл. 50, 1-5). Иногда прицветники несут на вершине листовидные придатки, особенно крупные в соцветиях костуса длинноостроконечного (табл. 50, 4). У вершины прицветников, а иногда и на прицветничках заметна зона железистой эпидермы, выделяющей нектар. У димерокостусов соцветия колосовидные, достигающие у димерокостуса шишковидного (D. strobilaceus) в длину 40 см. Прицветники димерокостуса имеют широкое воронковидное влагалище, охватывающее ось соцветия, иногда есть и листовидные пластинки на вершине. Только монокостус одноцветковый не имеет дифференцированного соцветия: его цветки располагаются по одному в пазухах верхних вегетативных листьев.
Цветки костусовых образуют трубчатую чашечку с трехзубчатым краем. У димерокостуса 2 чашелистика полностью срастаются и чашечка двулопастная. Лепестки этих цветков белые, желтые, оранжевые или красные, тоже срастаются у основания в трубку. Задний лепесток несколько крупнее боковых. Зигоморфный характер цветка определяет наиболее заметная его часть - направленная вперед крупная лепестковидная губа с большим волнистым или гофрированным отгибом. Известный немецкий морфолог Вильгельм Тролль (1928) пришел к выводу, что губа цветка костусовых является результатом срастания 5 стаминодиев. Края губы в нижней части обычно тоже сомкнуты в трубку, соединенную с трубкой венчика. Единственная фертильная тычинка у костусовых крупная, лепестковидная, принадлежащая к внутреннему кругу тычинок. Она расположена со стороны оси соцветия и, нависая, закрывает вход в цветочную трубку (рис. 226,5). Пыльник расположен в середине или в нижней части широкой тычиночной нити, гнезда его вскрываются продольно. Пыльцевые зерна костусовых покрыты толстой многопоровой экзиной.
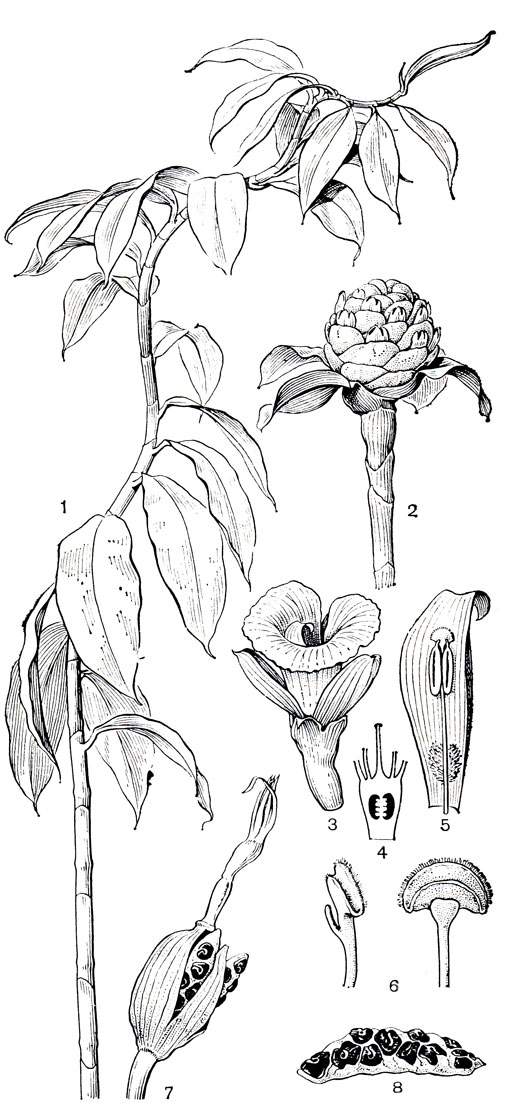
Рис. 226. Костусовые. Костус крупнопр и цветниковый (Costus megalobractea): 1 - вегетативный побег; 2 - соцветие. Костус Лукануса (С. lucanuslanus): 3 - цветок; 4 - продольный разрез завязи; 5 - тычинка с пыльниками, между ними - столбик с рыльцем; 6 - рыльце, вид с двух сторон. Костус длинноостроконечный (С. cuspidatus): 7 - раскрывающийся плод; 8 - семена из одного гнезда плода, слипшиеся благодаря клейким ариллусам
Гинецей костусовых синкарпный и состоит из трех плодолистиков. Завязь нижняя, у костуса 3-гнездная с центральной плацентацией, а у монокостуса, димерокостуса и тапейнохилуса 2-гнездная вследствие недоразвития одного плодолистика, с постенной плацентацией. Семязачатки анатропные, располагаются в один или два ряда.
В тканях завязи, над перегородками, имеются погруженные нектарники, открывающиеся протоком на поверхности. Над задней перегородкой, со стороны тычинки, нектарник развит слабее или совсем редуцирован. Столбик простой, нитевидный, в верхней части зажат между пыльниками. Рыльце воронковидное или из двух полулунных лопастей (рис. 226, б), с реснитчатым опушением по краю, иногда, например у костуса припудренного (С. pulverulentus), имеет с задней стороны вильчатый вырост, который находится между пыльниками и фиксирует рыльце.
Цветки костусовых раскрываются обычно утром, по одному в соцветии, а к вечеру уже вянут. Раскрытие цветков в одном соцветии происходит последовательно, начиная снизу. Эти цветки с крупной губой активно посещают пчелы. Пчела садится на губу и приподнимает нависающую лепестковидную тычинку, пробираясь в трубку цветка. Ее спинка, несущая пыльцу с других цветков, касается прежде всего рыльца. Затем насекомое проходит под пыльниками, вскрывающимися продольно и обсыпающими пчелу пыльцой. Ван дер Пэйл (1941), С. Фогель (1966) и другие биологи наблюдали на цветках димерокостуса и костуса пчел-плотников из рода ксилокопа (Xylocopa latipes и другие виды), а также многочисленных пчел эвглоссин (Euglossinae). В. Бурк (1891) и Ф. Ф. Р. Хайде (1927) наблюдали на Яве цветки костуса красивого с трубкой, поврежденной пчелой ксилокопой, высасывающей нектар снаружи. Однако такому "ограблению" цветков у большинства видов мешают плотные прицветники и муравьи, привлеченные в соцветие нектаром мелких внецветковых железок.
Цветки костуса с длинной и узкой цветочной трубкой и трубчато свернутой губой недоступны пчелам. Из них могут достать нектар только птицы колибри с достаточно длинным клювом. Голландский ботаник П. Мае, подробно изучавший костусовые в тропической Америке, наблюдал (1977) частые посещения соцветий костуса этими птицами. Цветки костуса Малорта опыляют и пчелы, и колибри. Для тапейнохилуса тоже характерно опыление птицами. Интересно, что ареал этого растения совпадает с ареалом райских птиц, опыляющих его цветки.
Плод костусовых - тонкостенная цилиндрическая или овальная 2-3-гнездная коробочка с остатками околоцветника на вершине. У многих видов костуса соплодие падает на землю вместе с отмирающим стеблем и оболочка нераскрывающихся плодов постепенно сгнивает, освобождая семена. Плоды монокостуса, тапейнохилуса, костуса скученноцветкового (Costus congestiflorus), костуса длинноостроконечного и других видов представляют собой локулицидную коробочку, раскрывающуюся сверху вниз. Некоторые виды обнаруживают септифрагное вскрывание коробочек, когда тонкие стенки отделяются от перегородок и крошатся. Так вскрываются плоды костуса арабского (С. аrаbicus) и костуса колосистого (С. spicatus), у которых после выпадения семян в пазухах прицветников торчат пустые трехкрылые колонки,; перегородки плода.
Семена костусовых цилиндрические или овальные, длиной 2-4 мм, коричневые или черные, с белым или желтым неправильно расщепленным ариллусом. У семян димерокостуса ариллус маленький, подушковидный. Поверхность ариллуса покрыта волосками, благодаря которым семена одного гнезда сцепляются и выпадают одним пакетом. Разъединение семян происходит на земле при намокании, когда гигроскопичные волоски становятся скользкими. Прорастание у костусовых надземное. Проросток выносит над поверхностью почвы довольно крупную семядолю с остатками семени на кончике, который является всасывающим органом, поставляющим питательные вещества проростку из семени. К сожалению, неизвестны факты наблюдений в природе за способом распространения семян костусовых, но по характеру ариллуса можно предположить мирмекохорию. Нередко можно обнаружить сеянцы вокруг материнского растения.
Необычная красота костусов привлекла внимание садоводов. Костус красивый (С. speciosus), костус длинноостроконечный (С. cuspidatus или С. igneus, табл. 50, 4-5) и другие виды выращивают в оранжереях ботанических садов. Некоторые виды костуса используют в местной народной медицине.
Семейство канновые (Cannaceae)
Американское семейство канновых включает лишь один род - канна (Саnnа, рис. 227, 228), насчитывающий около 50 видов, которые распространены главным образом в Центральной и Южной Америке, на севере до Южной Каролины и Флориды, а к югу - до северных районов Чили и Аргентины.
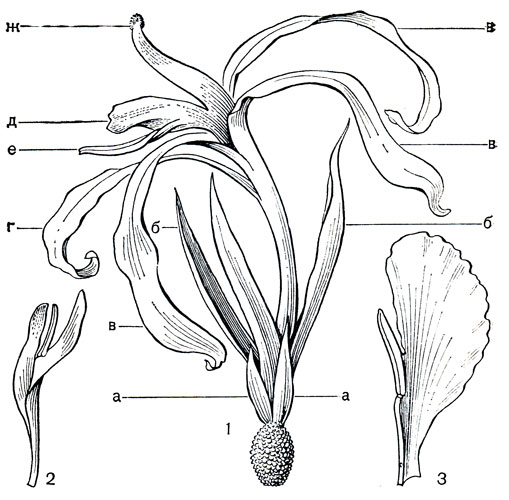
Рис. 227. Канновые. Канна фиолетовая (Canna violacea): 1 - цветок (а - чашелистики, б - лепестки, в - стаминодии, г - стаминодий - губа, д - тычинка, е - пыльник, ж - столбик с рыльцем). Канна многоветвистая (С. polyclada): 2 - столбик (слева) и тычинка с пыльником (справа). Канна повислая (С. flaccida): 3 - тычинка с пыльником
Канны растут по открытым солнечным местам на влажных, богатых гумусом почвах, по берегам рек и ручьев, на приморских равнинах и в горных ущельях. Канна Бриттона (С. brittonii) встречается в Андах Боливии на высоте до 2000 м над уровнем моря, а канна касатикоцветковая (С. iridiflora) была найдена в Перуанских Андах на высоте 2700 м. Некоторые виды канны сопутствуют человеческому жилью, образуя заросли по помойкам и свалкам на богатых азотом почвах, например канна лагунская (С. lagunensis). Некоторые виды канн, такие, как канна индейская (С. indica, рис. 228), канна низкая (С. humilis) и другие, стали неистребимыми сорняками на тропических плантациях. В то же время виды с крупными красивыми цветками подверглись истреблению и стали редкими или, быть может, даже исчезнувшими. К ним относятся канна касатикоцветковая из Перу и канна лилиецветковая (С. liliiflora) из Панамы с белыми душистыми цветками; они плохо приживались в культуре и исчезли из коллекций ботанических садов.
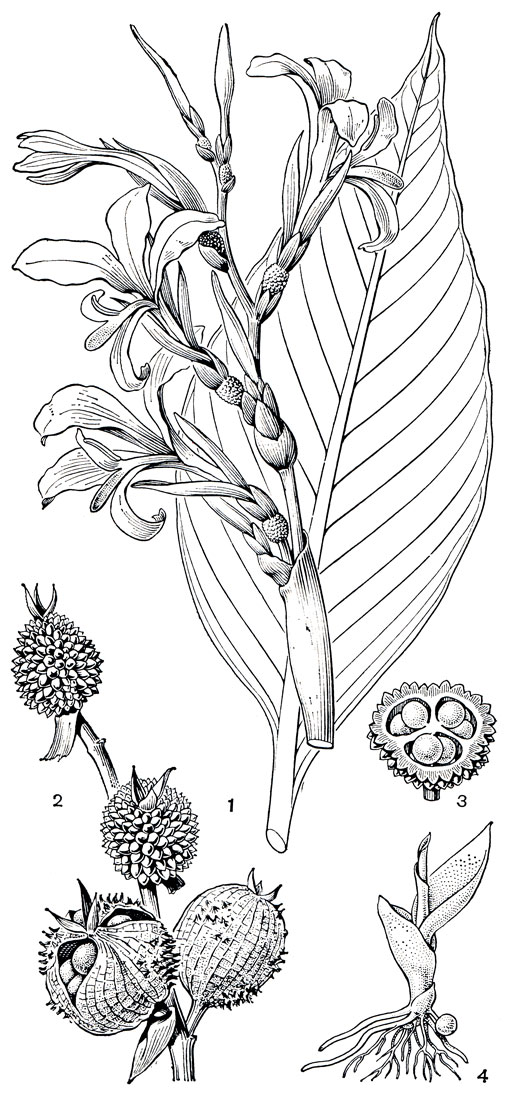
Рис. 228. Канна индейская (Canna indica): 1 - лист и соцветие; 2 - плоды; 3 - поперечный разрез плода; 4 - проросток
Канны - многолетние травянистые растения с симподиально ветвящимися корневищами и крупными листьями, двурядно расположенными на укороченных стеблях. Длинные открытые влагалища охватывают друг друга и образуют ложные стебли нередко высотой 1,5-2 м. Некоторые виды, как, например, канна красивая (С. formosa), растущая в Бразилии и Венесуэле, не превышает в высоту 1 м, а канна широколистная (С. latifolia) достигает 4 м и имеет листья почти метровой длины и шириной около 30 см. Широколанцетные или продолговатые, заостренные на вершине листовые пластинки канн слегка асимметричны, у основания клиновидно сужены и постепенно переходят во влагалище. От крупной главной жилки под острым углом отходят многочисленные боковые жилки со слабым сигмовидным изгибом, сбегающие к краям. Благодаря антоциану листья канн иногда имеют красноватый или бронзовый оттенок, как у канны Варшевича (С. warscewicziana). У канны сизой (С. glauca) и некоторых других видов листья покрыты голубоватым восковым налетом или имеют войлочное опушение, как у канны шерстистой (С. lanuginosa), защищающее растения от палящего солнца и избыточного испарения влаги.
У большинства канн толстые, мясистые, иногда клубневидно утолщенные корневища, содержащие запас питательных веществ и воды. Чешуевидные листья, двурядно расположенные на них, довольно быстро отмирают. Они сохраняются дольше у видов с тонкими корневищами.
Сосуды только в корнях, обычно с простой перфорацией, иногда с лестничной перфорацией и немногими перекладинами.
Первый лист надземного стебля не имеет развитой пластинки и не несет пазушной почки. В пазухах последующих, полноценно развитых листьев закладываются почки. Из них первыми и наиболее интенсивно развиваются почки в пазухах 4-, 5- и 6-го листа, а за ними трогаются в рост почки 7-, 8- и 9-го листа. Развивающиеся почки образуют новые части корневищ с шестью междоузлиями и придаточными корнями. Первые 2-3 междоузлия растут горизонтально, затем точка роста приобретает вертикальное направление и переходит вскоре к формированию надземного стебля. Стебли садовых канн развивают всего около 10 листьев, и над последним листом появляется верхушечное соцветие.
Ось соцветия канн трехгранная и несет спирально расположенные кроющие листья, свернутые в виде влагалища, в котором развиваются парциальные соцветия - завитки. В завитко закладывается 3-5 цветков, но из них развиваются обычно только 2, в редких случаях -3, а у некоторых видов канны завиток редуцирован до одного цветка, и соцветие приобретает характер простого колоса. Такие соцветия характерны для канны повислой (С. flaccida) канны геликониелистной (С. heliconifolia) и других видов. У многих видов главная ось соцветия ветвится. Боковые ветви, расположенные в пазухах крупных кроющих листьев, в свою очередь несут кроющие листья и парциальные соцветия. Нижняя боковая ветвь остается довольно короткой, а вторая превышает по длине главную ось, отклоняет ее в сторону и занимает ее место, принимая вертикальное положение. На боковой оси первого порядка могут образовываться ветви второго порядка. Среди них также вторая снизу имеет преимущественное развитие. У канны многоветвистой (С. polyclada, рис. 227,2) из Южной Бразилии соцветия образуют ветви четвертого порядка. Сильно ветвистые соцветия унаследовали некоторые сорта садовых канн.
Цветки канн резко асимметричны. Они обычно крупные, диаметром 4-8 см, яркие, желтые, оранжевые или красные, лишь немногие виды - канна лилиецветковая и канна Бриттона - имеют белые цветки. Цветки снабжены овальным или ланцетным прицветничком, обоеполые, трехчленные. Чашелистики неравной величины, свободные, черепитчато сложены в бутоне, зеленые или красноватые. Лепестки ланцетные, тоже неодинаковой величины, в нижней расти срастаются и образуют трубку (рис. 227). У большинства видов канны главную роль в облике цветка играют лепестковидно измененные стерильные тычинки - стаминодии. Все члены андроцея в нижней части срастаются с трубкой околоцветника. Единственная фертильная тычинка принадлежит внутреннему кругу андроцея, она тоже имеет лепестковидное строение и при этом асимметрична, с одним пыльником, расположенным у края. Один из стаминодиев внутреннего круга сильно отогнут книзу и выполняет роль губы, служа посадочной площадкой для опыляющих насекомых. У семи видов, составляющих подрод дистемон (Distemon), к которому относится, например, канна метельчатая (С. paniculata), из членов андроцея в цветках присутствуют лишь тычинка и стаминодий - губа. У некоторых видов имеются еще два крупных и ярких лепестковидных стаминодия, как у канны желтой (С. lutea), канны шерстистой и др. Большая часть видов имеет еще один крупный, четвертый стаминодий, расположенный позади тычинки. К таким видам относятся канна сизая, канна индейская, канна повислая. От них унаследовали этот признак и садовые канны.
Гинецей у канн синкарпный из трех плодолистиков; завязь нижняя, трехгнездная, снаружи покрытая мелкими бородавчатыми выростами, с многочисленными анатропными семязачатками. Столбик канн совершенно необычного строения: он плоский, лентовидный, у основания в большей или меньшей степени сросшийся со стерильной частью лепестковидной тычинки, также ярко окрашенный, и на первый взгляд его трудно выделить среди других лепестковидных органов цветка; лопасти рыльца в виде двух валиков выступают по верхнему краю столбика. Известный немецкий ботаник Карл Шуманн (1888), подробно изучавший представителей порядка имбирных, заметил в некоторых цветках канн полоску воспринимающей ткани и на наружном крае столбика. У основания столбика в цветках канн имеются отверстия септальных нектарников, погруженных в ткань завязи над перегородками. Нектар скапливается в трубке венчика.
Цветки канн раскрываются в 5-6 ч утра, только у канны повислой, произрастающей на юго-востоке США,- цветение начинается в 19-20 ч. Порядок раскрытия цветков в соцветии подчинен определенной закономерности. У садовых канн ее проследила в своих наблюдениях сотрудница Никитского ботанического сада Г. Ф. Феофилова (1976). Цветение начинается на главной оси с нижнего цветка первого нижнего завитка. На второй день раскрывается нижний цветок второго завитка и так далее. Интервал между раскрытием цветков в пределах одного завитка обычно 4 дня, иногда он колеблется от 2 до 6 дней. Продолжительность цветения всего соцветия садовых канн в среднем 26 дней. Каждый цветок функционирует 2-3 дня. Цветки дикорастущих видов недолговечны, они цветут лишь несколько часов. Пыльники вскрываются продольно еще в бутоне, примерно за 35 ч до раскрытия цветка, и пыльца высыпается в пространство между тычинкой и лепестковидным столбиком. Пыльцевые зерна канны сферические, с безапертурной шиповатой оболочкой, окончательно дозревают уже в полости бутона. Рыльце готово к восприятию пыльцы уже в бутоне, примерно за 2 дня до раскрытия цветка. У большинства видов канны, в отличие от других представителей порядка имбирных, преобладает самоопыление, происходящее еще в бутоне. Особенно легко оно осуществляется у тех видов, у которых столбик и тычинка одинаковой длины и пыльца легко попадает на рыльце. Самоопыление играет решающую роль у канны повислой и канны касатнкоцветковой, которые имеют повисающие цветки. У последнего вида столбик длиннее тычинки на 1,1 см, но в повисающем цветке рыльце оказывается ниже пыльника и легко опыляется своей пыльцой. Однако к строго самоопыляющимся растениям канны отнести нельзя. Наряду с самоопылением в их ярких, богатых нектаром цветках имеет место и перекрестное опыление. Чарлз Дарвин в работе "Действие перекрестного опыления и самоопыления в растительном мире" (1878) подробно исследовал механизм опыления канны Варшевича и пришел к выводу, что она дает одинаково жизнеспособное потомство как при самоопылении, так и при перекрестном опылении.
Лишь немногие виды канны, как, например, канна широколистная, совсем не могут опыляться без помощи посредника, так как столбик у них длиннее тычинки и своя пыльца не попадает на рыльце. По наблюдениям индийских ботаников И. Мукхерджи и Т. Н. Кхошу, проведенным в 1970 г. в Лакхнау, в цветках садовых канн рыльце на 0,3-1,9 см выше пыльника и на изолированных соцветиях семена не завязываются. Цветки садовых канн опыляют главным образом пчелы, шмели, бабочки. Насекомые садятся на стаминодий, играющий роль губы, и пробираются за нектаром к основанию столбика, касаясь спинкой липкого рыльца и нанося на него пыльцу, принесенную с других цветков. Так же происходит и опыление цветков дикорастущих видов. Лишь в очень широко открывающихся цветках спинка насекомого не достает до рыльца. На желтые цветки канны повислой насекомые вообще не могут сесть, так как его органы не имеют тургора и повисают, словно увядшие. У некоторых видов яркие цветки с длинной и узкой трубкой посещают и птицы, пьющие из них нектар и, возможно, при этом тоже переносящие пыльцу. Вскоре после опыления части цветка увядают и опадают целиком, лишь чашелистики сохраняются на вершине развивающейся завязи.
Плоды канны - 3-гнездные локулицидные коробочки, овальные или цилиндрические, созревают в течение 30-40 дней. Характерные бородавчатые, сочные выросты, покрывающие плод, к концу высыхают и опадают, оболочка становится тонкой, и коробочки медленно растрескиваются в верхней трети или до половины, освобождая круглые, черные семена диаметром 6-10 мм, расположенные двумя вертикальными рядами в каждом гнезде. Вес семян довольно стабилен для каждого вида, так что семена канны Бриттона в Боливии служили некогда эталоном для взвешивания золота. Оболочка семян блестящая, необычайно твердая. Семена канн долго сохраняют всхожесть. Известен случай, когда через тридцать лет проросли семена канны съедобной (С. edulis), собранные исследователями флоры Южной Америки И. Руисом и X. А. Павоном. Зародыш, окруженный тонким слоем эндосперма, лежит в белом и твердом перисперме и имеет несколько зачатков листьев. Прорастание у канн подземное.
Длительное сохранение всхожести семян, быстрое развитие корневищ и возможность вегетативного размножения - все эти факторы сыграли роль в распространении и натурализации некоторых видов канны во многих тропических странах.
Канны с древних времен культивировались индейцами тропической Америки из-за крахмалистых корневищ, используемых в пищу в печеном виде. Древним культурным растением является канна съедобная, называемая "ачирой". В Перу археологами найдены остатки корневищ в захоронениях, датируемых около 2500 г. до н. э. В настоящее время ачиру, канну широколистную, канну сизую и некоторые другие виды продолжают выращивать индейцы Южной Америки. Корневища канны съедобной содержат около 27% крахмала с очень крупными зернами, известного в торговле под названием "квинслендский аррорут". Стебли и листья идут на корм скоту. Кроме стран Латинской Америки, канну съедобную возделывают в Индии, Индонезии, Австралии, на Гавайских островах.
Голландские систематики В. Сегерен и П. Мае (1971) считают, что канна съедобная, по-видимому, представляет собой культурную форму канны индейской, и относят ее к этому виду, широко натурализовавшемуся в тропиках.
В качестве декоративного растения канна индейская была завезена в Европу португальскими мореплавателями в конце XVI в. В XVII в. в каталогах ботанических садов Парижа, Лейдена, Нюрнберга числится уже несколько видов канны. В середине XIX в. на юге Франции появились первые сорта декоративных канн, выведенные Теодором Анне путем скрещивания канны сизой и канны индейской. Впоследствии его сорта были вытеснены завоевавшими славу гибридами Пьера Крози, работавшего в Лионе. Он начал скрещивания канны Варшевича и канны повислой и в короткое время создал сложные гибриды с участием канны индейской и канны сизой, у которых цветки достигали в диаметре 10 см и напоминали по облику гладиолусы. Созданные Крози 180-200 сортов объединяют в сортотип "канн Крози" или под названием "садовые канны" (С. X generalis).
Следующим этапом в истории декоративных канн было создание так называемых орхидеевидных канн (С. X orchioides), созданных в Италии известным селекционером M. Шпренгером и действительно напоминающих цветки орхидеи каттлеи. В создании этих сортов использовались гибриды Крози, опылявшиеся пыльцой канны повислой. Среди орхидеевидных канн преобладают триплоиды, и многие из них стерильны, но зато их нежные цветки с гофрированными стаминодиями достигают в диаметре 16 см (сорт Индиана - Indiana) и даже 21 см (сорт Винтцер'с Колоссаль - Wintzer's Colossal).
В Россию канны были ввезены при Петре I и выращивались в оранжереях первых ботанических садов в Москве и Петербурге. Центром выращивания и селекции канн в нашей стране стал Никитский ботанический сад в Ялте. В настоящее время там выращивают богатую коллекцию видов и сортов канн; имеются сорта отечественной селекции.
Семейство марантовые (Marantaceae)
Семейство марантовых, обнаруживающее черты наиболее высокой организации в порядке имбирных, объединяет около 30 родов и 400 видов, распространенных главным образом в тропической зоне всех континентов, за исключением Австралии. Немногие виды марантовых произрастают в субтропиках и лишь редко встречаются в зоне умеренно теплого климата. Особенно богата марантовыми флора Южной и Центральной Америки, включающая 11 родов этого семейства и среди них самый большой - калатея (Calathea, рис. 229, 230), который объединяет около 150 видов. Наиболее широкое распространение с севера на юг в западном полушарии обнаруживают виды рода талия (Thalia). Талия беловатая (Т. dealbata) встречается в Техасе, Луизиане и юго-восточных штатах США до 37° с. ш., а талия многоцветковая (Т. multiflora) достигает в Уругвае 33° ю. ш. Талия коленчатая (Т. geniculata) растет на болотах Флориды, а к югу распространена до Аргентины; этот вид встречается и в Западной Африке. В Африке марантовые представлены главным образом эндемичными родами и преобладают во влажном тропическом климате западной части континента от Сьерра-Леоне на севере до Анголы на юге. В восточной части материка произрастают лишь немногие виды. На Мадагаскаре из марантовых встречаются лишь галопегия Перръе (Halopegia perrieri) и ктенофриниум односторонний (Ctenophrynium unilaterale), представляющий монотипный эндемичный род. В тропической Азии наибольшее число видов марантовых произрастает в лесах полуострова Малакка и на Яве, немногие виды встречаются в Индии и на острове Шри-Ланка. Среди них наиболее широкое распространение имеет фриниум головчатый (Phrynium capitaturn), растущий в Индии, Бирме, Юго-Западном Китае и доходящий к югу до Явы. С этим видом по величине ареала могут соперничать лишь некоторые виды рода донакс (Donax), распространенные в Индии, Бирме, на полуостровах Индокитай и Малакка, на островах Малайского архипелага и Океании.
Марантовые - характерные обитатели тропических дождевых лесов, где они образуют нередко непроходимые заросли. Они часто встречаются на низких болотистых и даже затопляемых местах, по берегам водоемов, вдоль рек и ручьев. Шуманниантус вильчатый (Schumannianthus dichotomus) произрастает в мангрове восточного полушария вместе с пальмой нипой или поселяется на рисовых полях. Некоторые марантовые обитают в районах с засушливым периодом в листопадных и полулистопадных лесах, переживая засуху в виде корневищ. К таким видам относятся, например, калатея широколистная (Calathea latifolia) и калатея крупночашелистиковая (С. macrosepala) из Южной Америки; последняя особенно засухоустойчива. Марантовые являются частыми обитателями вторичных формаций, образуя сплошные заросли на вырубках или в заброшенных ирригационных каналах. Большинство видов произрастает на высоте до 1000 м над уровнем моря, рекорд высотного распространения представляет калатея Тимоти (С. timothei), растущая по склонам Анд на высоте 2100 м.

Таблица 51. Канновые и марантовые: 1 - канна гибридная (Canna hybrida), экспериментальная база Ботанического института АН СССР в Отрадном Ленинградской области; 2 - канна садовая (С. X generalis), сорт 'Восход', Крым, Никитский ботанический сад; 3 - канна садовая, сорт 'Хамелеон', там же; 4 - канна орхидеевидная (С. X orchioides), сорт 'Фойерфогель', там же; 5 - калатея полосатая (Calathea zebrina), оранжереи Ботанического института АН СССР в Ленинграде; 6 - маранта беложилковатая (Maranta leuconeura), оранжереи Ленинградского государственного университета
Марантовые - многолетние травянистые растения с симподиально ветвящимися корневищами. Корневища их волокнистые и иногда жесткие, деревянистые, покрыты двурядно расположенными чешуевидными, нередко опушенными листьями. У некоторых видов корневища образуют направленные в глубь почвы утолщенные столоны, в которых запасаются вода и питательные вещества. Такие клубневидные столоны образует маранта тростниковидная (Maranta arundinacea, рис. 229, 5). Многие виды маранты, калатеи, саранты (Saranthe) и других родов имеют клубневидно утолщенные корни. У калатеи крупночашелистиковой корневые клубни достигают в диаметре 6 см. У маранты двуцветной (M. bicolor, рис. 230, 4-6) утолщено основание цветоносного стебля. Корневища марантовых обычно имеют короткие междоузлия, так что надземные побеги образуют густые куртины. Стебли марантовых большей частью хорошо развиты, симподиально ветвятся, иногда достигают большой высоты и становятся деревянистыми. Для марантовых характерно образование междоузлий, очень неравных по длине. Внизу стебель развивает обычно влагалищные листья на очень коротких междоузлиях, а междоузлие, несущее первый лист с развитой пластинкой, как правило, длиннее остальных. У видов донакса оно имеет длину около 3 м, а последующие междоузлия так коротки, что листья, собранные густым пучком, венчают его на вершине. Некоторые виды строманты (Stromanthe), миросмы (Myrosme) и других родов образуют над пучком сближенных листьев снова длинное междоузлие, несущее второй ярус густо собранных листьев. Виды африканских родов гипселодельфис (Hypselodelphis), трахифриниум (Trachyphrynium) и некоторые виды южноамериканского рода исхносифон (Ischnosiphon, рис. 230, 9) имеют бамбуковидный габитус. Их стебли растут быстро в высоту, сохраняя почти одинаковый диаметр, на их утолщенных узлах имеются чешуевидные листья, которые вскоре опадают. Из пазушных почек развиваются симподиально ветвящиеся боковые побеги. Для марантовых характерно наличие предлиста в основании каждой ветви. Предлист имеет два килевидных продольных выступа на стороне, прижатой при развитии к главному стеблю. У видов гипселодельфиса предлистья достигают в длину 10 см; большей частью они мелкие, опадающие. После предлиста ветвь обычно несет влагалищный лист, а затем уже листья с развитой пластинкой. Ветви у марантовых отходят под определенным для каждого вида углом, образуя зигзаг или завиток. Такой характер ветвей способствует их закреплению на ветвях соседних кустарников и деревьев. Таким образом стебли гипселодельфиса Ценкера (Н. zenkerianum), растущего в тропических лесах Западной Африки, поднимаются, цепляясь ветвями за окружающие деревья, на высоту до 10 м. Ветви и оси соцветий хауманнии Данкельманна (Haumannia dankelmanniana) оснащены крючковидно загнутыми колючками, которыми растение цепляется за листья и ветви деревьев и создает таким образом непроходимые заросли в лесах Западной Африки. Многие виды калатеи, маранты, фриниума и других родов имеют, напротив, очень короткий стебель с розеткой листьев. Число листьев, развиваемых на стеблях до образования соцветий, как правило, определенное у каждого вида. У некоторых марантовых оно сокращено до одного листа, обычно вертикально стоящего на длинном черешке. Такой габитус имеют виды африканских родов мегафриниум (Megaphrynium), афрокалатея (Afrocalathea) и тауматококкус (Thaumatococcus), единственный вид которого тауматококкус Даниэля (Т. daniellii) имеет черешок листа длиной 2-3 м.
Листья марантовых расположены двурядно на стебле и имеют хорошо развитое незамкнутое влагалище, у некоторых видов с лигулой. На черешке листа около прикрепления пластинки всегда имеется особое утолщенное со-членение (лат. pnlvinus), по наличию которого марантовые легко узнать в вегетативном состоянии. Проводящие пучки, разбросанные в тканях черешка, в области этого утолщения группируются в центре, а по периферии под хлорофиллоносной палисадной тканью располагается слой длинных радиально и косо направленных тонкостенных клеток с водянистым клеточным соком (рис. 229). Колебания тургора этих клеток вызывают изменение угла наклона листа. Такой механизм позволяет регулировать положение пластинки по отношению к свету и имеет большое биологическое значение для растений, живущих в густой тени под пологом тропического леса.
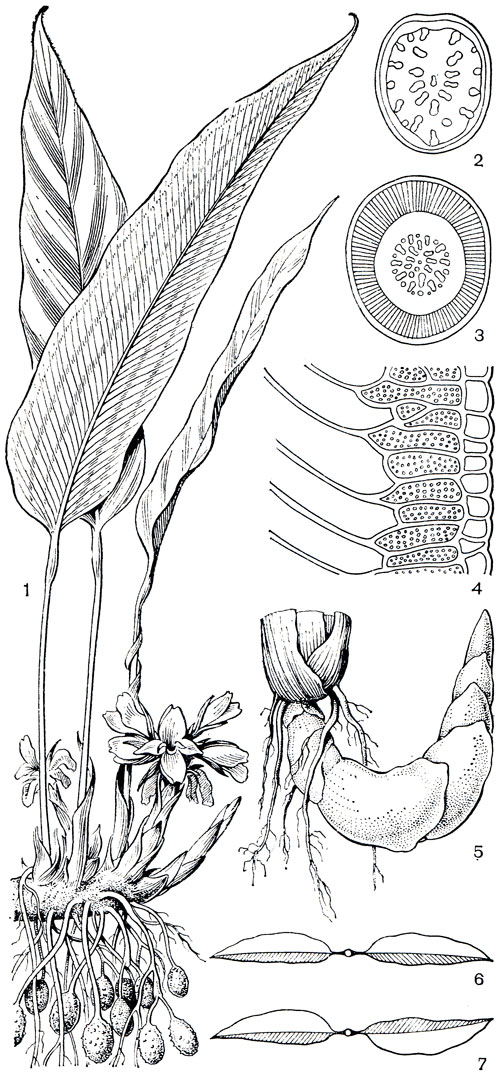
Рис. 229. Марантовые. Калатея Баше (Calathea bachemiana): 1 - цветущее растение; 2 - поперечный разрез черешка; 3 - поперечный разрез утолщения на черешке; 4 - его продольный разрез. 1аранта тростников и дная (Maranta arundinacea): 5 - корневище. Схема расположения листьев на междоузлии: 6 - антитропный тип; 7 - гомотропный тип
В течение суток листья марантовых совершают движения, улавливая солнечные лучи. Многие виды калатеи и маранты, в том числе известное комнатное растение маранта беложилковатая (М. leuconeura) из лесов Бразилии вечером поднимают свои листья вверх и складывают их вместе, то же происходит и за несколько часов перед грозой. Таким образом эти растения предсказывают погоду. Пластинка листа марантовых овальная, продолговатая, ланцетная или почти линейная, с крупной центральной жилкой. Многочисленные боковые жилки слегка изогнуты и соединены через определенные интервалы характерными поперечными анастомозами. Как у всех представителей порядка имбирных, листья марантовых асимметричны в результате развития внутри трубки влагалища, которая стесняет рост наружной половины трубчато свернутого листа. У некоторых марантовых узкая наружная половинка листа имеет постоянное положение справа или слева от главной жилки. Такие листья, называемые гомотропными, преобладают у калатеи, маранты, исхносифона и других родов. У многих марантовых листья с левой узкой половинкой чередуются с листьями, у которых она расположена справа. Такие листья, называемые антитропными, характерны, например, для видов строманты и трахифриниума. Нижняя сторона листьев марантовых иногда окрашена в пурпурный или лиловатый цвет, а верхняя часто имеет пестрый рисунок за счет чередования темных и светлых полос или в виде темных пятен на светлом фоне. Пятнистость иногда сочетается с белыми или розовыми жилками, шелковистым блеском или бархатистой поверхностью эпидермы, клетки которой имеют прозрачные линзовидные утолщения оболочки, концентрирующие световые лучи. Темные пятна на листьях образуются за счет более высокого содержания пластид в клетках, белый цвет связан с образованием воздухоносных межклеточных пространств, а розовый и красный - с наличием антоциана в клетках. Иногда пестрая окраска присуща лишь ювенильным листьям, а взрослые растения развивают однотонные зеленые листья. Характер пестрого рисунка является систематическим признаком, позволяющим определить вид или его разновидность. Листья некоторых марантовых покрыты восковым налетом, как, например, у талии беловатой. Особенно обильный восковой налет на нижней, абаксиальной стороне имеют огромные, полутораметровой длины листья калатеи желтой (С. lutea), растущей по болотистым местам в Центральной Америке и на островах Карибского моря.
Марантовые нередко имеют опушение из одноклеточных волосков с погруженным основанием. В корнях, а иногда и в стеблях и изредка в листьях марантовых имеются сосуды с простой или лестничной перфорацией.
Многие марантовые зацветают в первый год жизни, но отдельные лесные виды цветут исключительно редко. Соцветия марантовых занимают верхушечное положение на облиственных стеблях или на специализированных безлистных цветоносах, вырастающих от корневища. Как правило, марантовые имеют сложные соцветия, состоящие из нескольких элементарных соцветий, которые представляют собой колосовидный тирс с двурядно расположенными кроющими листьями. У некоторых видов калатеи кроющие листья располагаются тесной спиралью, в компактных соцветиях головчатого типа. Кроющие листья зеленые или ярко окрашены, и в этом случае они, несомненно, играют роль в привлечении опылителей. Кроющие листья обычно ладьевидно сложены и содержат в пазухах парциальные соцветия, которые представляют серповидную серию последовательно развивающихся монохазиев. У калатеи в парциальном соцветии может развиваться от 4 до 13 монохазиев, а у исхносифона неравностороннего (Ischnosiphon obliquus) их число достигает 17. В то же время у южноамериканского рода монофилланта (Monophyllanthe) и некоторых видов cap анты парциальное соцветие состоит лишь из одного монохазия. В основании монохазий несет обычно двухкилевой или трехкилевой предлист, два цветка монохазия зеркально-симметричны. Монохазий марантовых, по-видимому, является результатом редукций до двух цветков многоцветкового завитка, столь характерного для всего порядка имбирных. У видов южноамериканского рода строманта можно заметить рудимент третьего цветка, иногда третий цветок бывает хорошо развит. У монотагмы (Monotagma) и монофриниума (Monophryninm) произошла редукция монохазия до одного цветка, причем монотагма имеет парциальное соцветие из 3 или 4 редуцированных монохазиев, а у монофриниума в пазухе кроющего листа находится единственный цветок. В зависимости от длины осей и цветоножек тирс может быть компактным колосовидным или даже головчатого облика, а может иметь и рыхлую кистевидную структуру. У многих марантовых главная ось соцветия симподиально ветвится, благодаря чему образуются сложные соцветия с многими тирсами, венчающими каждую ветвь. В соцветиях марантовых часто имеются прицветники, обычно они пленчатые, слабо окрашенные, у многих видов они быстро опадают. Прицветнички, имеющиеся у многих марантовых, обычно маленькие, пленчатые, а у цветков некоторых видов калатеи, фацелофриниума (Phacelophrynium) и исхносифона они нитевидные, булавовидно утолщенные на конце и, видимо, играют роль амортизаторов для нежных структур цветка, зажатых жесткими кроющими листьями. На прицветничках калатеи портобелъской (С. portobelensis) имеются внецветковьте нектарные желёзки, привлекающие в соцветия многочисленных муравьев.
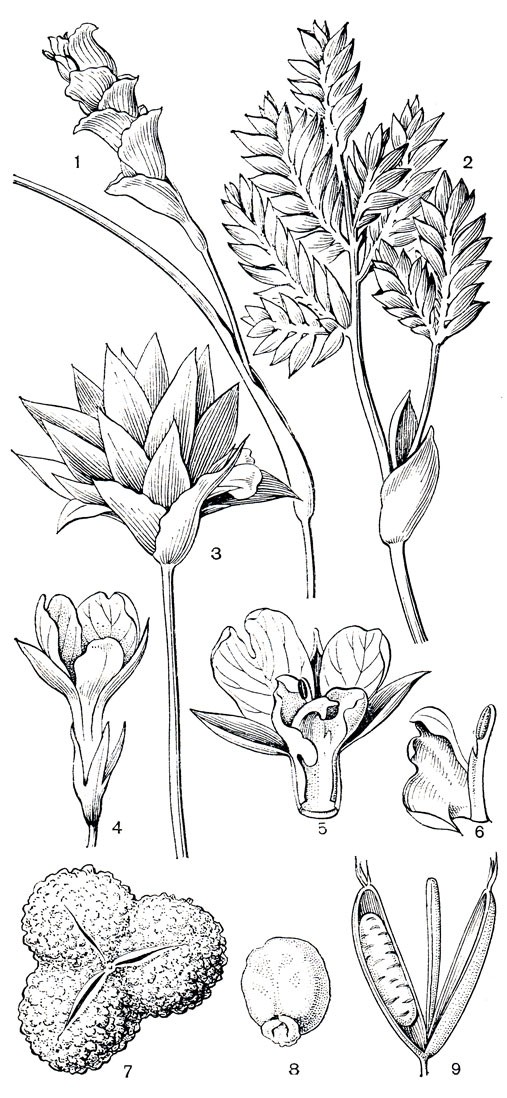
Рис. 230. Марантовые. Соцветия: 1 - калатея Глазиова (Calathea glaziovii); 2 - саранта Риделя (Saranthe riedeliana); 3 - калатея головчатая (Calathea capitata). Маранта двуцветная (Maranta bicolor): 4 - цветок; 5 - раскрытый цветок; 6 - капюшоновидный стаминодий (слева) и тычинка (справа). Хаумания Данкельманна (Haumannia dankelmanniana): 7 - плод. Калатея крупноцветковая (Calathea grandiflora): 8 - семя. Исхносифон неравносторонний (Ischnosiphon obliquus): 9 - раскрывшийся плод с одним семенем
Цветки марантовых обоеполые и, так же как у каиновых, асимметричные. Чашелистики обычно свободные, а лепестки у основания срастаются в трубку. Лепестки неравные по величине (адаксиальный лепесток крупнее остальных и несколько вогнут), большей частью белые или кремовые, редко розовые или голубые. Из двух кругов андроцея у марантовых полностью развит лишь внутренний. Так же как у канновых, все тычинки их имеют лепестковидное строение и срастаются с трубкой венчика у зева. Фертильна лишь одна тычинка, несущая сбоку лепестковидной нити только одно гнездо пыльника. Стаминодий, который у каиновых играет роль губы, у марантовых довольно короткий, упругий, с двумя утолщенными участками, за что получил название мозолистого стаминодия. Другой стаминодий внутреннего круга имеет капюшонообразное строение и в бутоне охватывает столбик и прижатую к нему тычинку. Один край его имеет крючковидный вырост, играющий важную роль в специализированном механизме опыления марантовых. В наружном круге андроцея марантовых всегда отсутствует передний, абаксиальный член и развиты обычно один или два лепестковидных стаминодия. В цветках тауматококкуса, некоторых видов калатеи, строманты и других родов стаминодии наружного круга не развиты совсем.
Единственная половина пыльника марантовых вскрывается продольно еще в бутоне. Пыльцевые зерна гладкие, с безапертурной оболочкой. Гинецей из 3 плодолистиков, синкарпный; завязь нижняя, 3- или 1-гнездная за счет недоразвития двух гнезд. В каждом гнезде почти базально расположен единственный анатропный или кампилотропный семязачаток. В тканях завязи над перегородками всегда имеются крупные овальные нектарные желёзки, открывающиеся у основания столбика. Нектар накапливается в нижней части трубки венчика, которая прикрыта иногда волосками опушения. Столбик короткий, довольно толстый, изогнут на вершине так, что рыльце, находящееся в воронковцдном углублении, направлено несколько вниз и в сторону от тычинки, и пыльца своего пыльника на него не попадает, а высыпается в особое углубление над рыльцем. Клейкие выделения клеток, выстилающих края углубления, не дают пыльце высыпаться в полость бутона. В бутоне лепестки спирально свернуты, а крючковидный столбик приподнят, несколько выпрямлен краем капюшоновидного стаминодия и находится в напряжении.
Цветки марантовых опыляют мелкие виды пчел. Американский ученый Элен Кеннеди (1978), изучающая марантовые в природе, наблюдала на их цветках пчел из родов эвглосса (Euglossa) и эвлема (Eulaema). Пчела обычно садится на стаминодии наружного круга и по мозолистому стаминодию, как по желобу, вводит свой хоботок в цветочную трубку. На пути насекомого, пробирающегося за нектаром, словно взведенный курок, стоит крючковидный вырост капюшоновидного стаминодия. Толчок поэтому триггерному устройству отодвигает этот стаминодий, приподнятый столбик освобождается и, сгибаясь, хлопает рыльцем, по насекомому, забирает имеющуюся на нем пыльцу, принесенную с других цветков, и одновременно стряхивает на него свою пыльцу. По К. Фегри и Л. Ван-дер-Пэйлу (1966), в цветках калатеи, исхносифона и монотагмы пыльца высыпается точно в ямку у основания хоботка, где пчела ее не достает, когда чистится. Виды пчел с длинными хоботками, а также колибри иногда "грабят" нектар в цветках калатеи, не трогая крючковидного выроста и не приводя в действие механизм опыления. Э. Кеннеди удалось проследить узкоспециализированные связи видов калатеи с видами опылителей. Например, крупные цветки калатеи Донелл - Смита (Calathea donell - smithii) опыляет пчела эвлема мериана (Eulaema meriana), а в более мелких цветках с короткой трубкой она пьет нектар, не опыляя их. Среди видов калатеи подрода псевдофриниум (Pseudophrynium) Э. Кеннеди обнаружила довольно обширную группу с неоткрывающимися цветками, в которых, однако, не происходит самоопыления. Их насильственно открывают и опыляют пчелы эвлемы. Пчела жует кончик бутона, чтобы разъединить лепестки, а затем головкой поднимает медианный лепесток и пробирается в цветок. Немногие виды калатеи имеют настоящие клейстогамные цветки, к таким относится калатея панамская (Calathea panamensis). Несколько видов калатеи имеют красные кроющие листья соцветий и желтые цветки, например калатея боковая (С. lateralis), растущая в Андах Колумбии и Перу. Сочетание красного цвета с желтым характерно для птицеопыляемых соцветий геликоний и некоторых бромелиевых. Э. Кеннеди (1977) высказывает предположение о возможном переходе к опылению птицами у калатеи. Крючковидный вырост на капюшоновидном стаминодии у этих видов редуцирован, и обычный для марантовых механизм опыления действовать не может.
Плоды марантовых - трехгнездные или одногнездные локулицидные коробочки или односемянные орешковидные или ягодообразные. У видов галопегии два гнезда завязи остаются стерильными и оболочка единственного семени полностью срастается с оболочкой плода, образуя семянку. Виды саркофриниума (Sarcophrynium) и тауматококкуса Даниэля, растущие в тропической Африке, имеют сочные плоды с желеобразным эндокарпием, у последнего - очень сладкие, их охотно поедают обезьяны и другие животные. Плоды донакса канновидного (Donax cannaeformis), обитающего по морским побережьям Малайзии, Малайского архипелага и Новой Гвинеи, легко плавают в воде благодаря пробкообразному эндокарпию и разносятся течениями.
Семена марантовых большей частью имеют ариллус, микропиле на семени закрыто круглой крышечкой, которую выталкивает корень зародыша при прорастании. Зародыш сильно изогнут и окружен обильным периспермом. Ариллус, несомненно, играет роль в раскрытии коробочек. У некоторых видов калатеи, стахифриниума (Stachyphrynium) и других родов ариллус имеет выросты, которые в сухом плоде сложены компактно, а набухая, распрямляются и, словно пружины, раскрывают плод. У марантовых с нераскрывающимися плодами семена обычно без ариллуса. Ариллус имеет биологическое значение и для распространения семян. Полевые наблюдения и эксперименты американских биологов К. Хорвица и А. Битти (1980), проведенные в тропических лесах на юге Мексики, убедительно доказывают участие муравьев, питающихся ариллусами калатеи, в распространении ее семян. Немалую роль в распространении семян марантовых играют небольшие птицы, живущие в подлесках влажных тропических лесов. Семена видов талии, живущих по берегам водоемов, нередко поедают утки. Семена марантовых обладают способностью покоиться, сохраняя жизнеспособность. По наблюдениям Э. Кеннеди (1978), семена видов калатеи, произрастающих в районах с засушливым периодом, не прорастают до начала новых дождей.
Семейство марантовых делится на две трибы: фриниевых (Phrynieae) и марантовых (Маrаnteae). Для фриниевых характерна 3-гнездная завязь, но у видов фриниума не развит иногда один семязачаток, а у фриниума мелкоцветкового и видов галопегии недоразвитыми остаются два гнезда завязи. Триба марантовых характеризуется развитием лишь одного - переднего, абаксиального гнезда завязи.
К семейству марантовых принадлежит одно из древних культурных растений - маранта тростниковидная, или вест-индский аррорут (Maranta arundinacea, рис. 229, 5). Ее веретеновидно утолщенные корневища используются как источник крахмала, который является ценным диетическим продуктом. Маранта тростниковидная произрастает в тропических лесах Южной Америки и культивируется повсюду в тропиках. Как заменитель аррорута в Гвинее выращивают маранту арума (М. arouma). Стебли видов донакса и некоторых других марантовых расщепляют и используют для плетения корзин и других изделий. Пестролистные виды калатеи и маранты выращивают как декоративные растения в оранжереях и комнатах.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'