Порядок рестиевые (Restionales)
Семейство флагеллариевые (Flagellariaceae) (Т. Г. Леонова)
Монотипное семейство включает один род флагеллария (Flagellaria, рис. 186, 187) и 3 вида, распространенных в тропиках Старого Света, но преимущественно на островах Тихого океана. По пыльце и своему анатомическому строению, особенно устьиц и эпидермальных клеток, оно обнаруживает большое сходство со злаками.
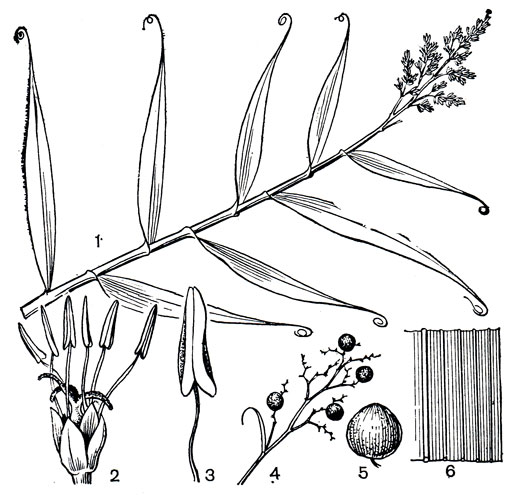
Рис. 186. Флагеллария гвинейская (Flagellaria guineensis): 1 - верхняя часть стебля с соцветием; 2 - цветок; 3 - тычинка; 4 - веточка соцветия с плодами; 5 - плод; 6 - сегмент листа
Флагеллариевые представляют собой многолетние лесные лианы с коротким, покрытым чешуйками симподиальным корневищем, в котором запасается сахароза, а не крахмал, как у большинства растений; каждый конец корневища несет только по 2 почки возобновления. Стебли у них одиночные, мощные, длиной от 2 до 15 м; они вальковатые, травянистые, но твердые и крепкие (напоминают виноградную лозу), в основании более или менее деревенеющие. Стебли простые или равно дихотомически ветвятся только в верхней части, не разделены на узлы и междоузлия и не имеют вторичного утолщения. Листья густо расположены по всему стеблю, очередные, двурядные, жесткие и напоминают листья злаков, в почкосложении улиткообразно свернутые. Они линейные или имеют форму от яйцевидных до ланцетовидных, нескладчатые, изолатеральные или дорсивентральные, цельнокрайные, с густым продольным и параллельным жилкованием и с короткими, косыми и тонкими поперечными жилочками; крупные жилки дугообразно сходятся в основании пластинки. Длина листьев колеблется от 3 до 50 см, а ширина - от 0,5 до 6,5 см. В основании листья закругленные или слегка сердцевидные, сидячие или же внезапно переходят в дорсально уплощенный черешок длиной 3-10 мм. Этот черешок или основание пластинки у сидячих листьев переходят в вальковатое, трубчатое и замкнутое почти до самой усеченной верхушки влагалище. Влагалища плотно охватывают стебель и в месте перехода в пластинку или черешок имеют, как у злаков, ушки в виде узкого и тонкого ободка; язычок отсутствует. К верхушке листья суживаются и, кроме самых нижних, заканчиваются длинным и тонким, дорсально уплощенным твердым усиком, который закручивается наподобие часовой пружины (рис. 186,187). С помощью этих усиков флагелларии цепляются за деревья и кустарники, по которым они высоко поднимаются вверх. Как только усики захватывают опору, они утолщаются и становятся еще более твердыми и крепкими. Листья, как и все растение, голые. Устьица парацитные (М. А. Баранова, 1975), замыкающие клетки их граминоидного типа. Сосуды имеются в стеблях и листьях, членики их с простой или лестничной перфорацией. Стенки клеток всего растения, особенно эпидермиса, обильно пропитаны кремнеземом, а некоторые клетки содержат скопления кремнезема в виде округлых тел. В паренхиме стебля имеются рассеянные секреторные клетки, а в мезофилле листьев наблюдаются секреторные каналы и маленькие гроздья кристаллов оксалата кальция.
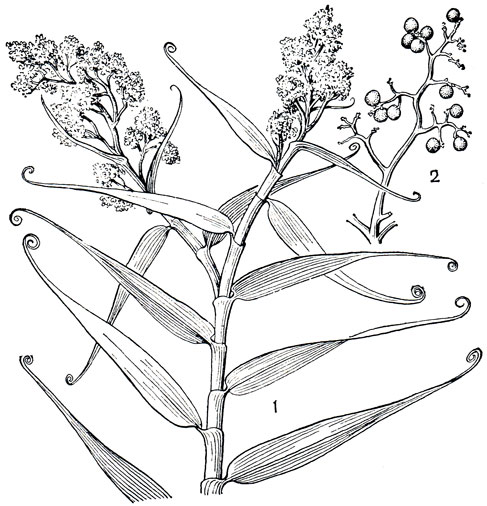
Рис. 187. Флагеллария индийская (Flagellaria indica): 1 - верхняя часть стебля с соцветиями; 2 - ветвь с плодами
Цветки флагеллариевых актиноморфные, мелкие и малозаметные, обоеполые, трехчленные, сидячие, с запахом. Они многочисленные, с прицветничками, собраны в конечные, более или менее густые, сильно ветвистые, снабженные опадающими кроющими листьями метелки длиной 3-50 см. Прицветнички мелкие, широкие и вогнутые, с ушками. Околоцветник, остающийся при плодах, состоит из 6 перепончатых продолговатых и тупых, беловатых или желтых, расположенных в 2 круга сегментов, наружные из которых в 2 раза короче внутренних. Тычинок 6. Они свободные, располагаются в 2 круга, выступают из околоцветника и имеют нитевидные тычиночные нити, к которым пыльники прикрепляются своим основанием.
Пыльники линейные или линейно-продолговатые, в основании стреловидные, интрорзные, открываются продольной щелью. Пыльцевые зерна однопоровые, граминоидного типа. Гинецей синкарпный, из 3 плодолистиков; завязь верхняя, 3-гнездная и трехгранная, содержит в каждом гнезде по 1 висячему семязачатку. Столбик короткий или отсутствует; рыльца в числе 3, вальковатые, с сосочками, довольно длинные, как и тычинки, выступают из околоцветника. Пыльник и рыльца созревают одно временно (гомогамия). Цветут и плодоносят флагелларии одновременно, опыляются ветром. Т. Ньюел (1969) считает, что в опылении принимают участие и муравьи.
Плоды костянковидные, нераскрывающиеся, полушаровидные, остроконечные, розовые или ярко-красные, содержат по 1, реже по 2-3 семени. Экзокарпий плодов тонкий и мясистый; эндокарпий очень твердый, напоминает кость, гладкий или бороздчатый (так называемая косточка). Семена шаровидные или несколько сплюснутые и уплощенные, с корковидной оболочкой; эндосперм обильный, мучнистый, содержит, кроме того, запасной крахмал в виде простых зерен; зародыш маленький, чечевицевидный, слабодифференцированный, окружен эндоспермом. Плоды флагеллариевых обладают некоторой плавучестью благодаря наличию в эндокарпии двух небольших полостей, заполненных воздухом. На этом основании Г. Ридли (1930) считает, что плоды флагелларии разносятся главным образом морской водой и морскими течениями как на близкие, от одного берега к другому, так и на значительные расстояния - от острова к острову. Подтверждение своей точки зрения Ридли видит и в экологии флагелларии. Все 3 вида этого рода растут в тропических сырых, но не заболоченных прибрежных лесах преимущественно на морских побережьях, иногда каменистых, и на влажно-грязных берегах тех рек, по которым поднимается морской прилив,- чаще по окраинам лесов и вдоль внутреннего края мангровых зарослей, где они карабкаются на деревья и кустарники. Реже флагелларии обитают вдоль берегов рек в горах на высоте до 1500 м над уровнем моря. По мнению Т. Ньюела, плоды флагелларии расселяются также и человеком, поскольку она широко им используется и довольно часто растет вдоль дорог. Возможно также, что в какой-то степени плоды переносят и птицы.
Наиболее широко распространена и известна флагеллария индийская (F. indica, рис. 187), встречающаяся в Индии, Таиланде, на полуострове Малакка и на островах Мадагаскар, Маскаренские, Сейшельские, Шри-Ланка, Суматра, Ява, Калимантан, Сулавеси, Хайнань, Тайвань, Филиппины, Новая Гвинея, Соломоновы, а также на севере Австралии. Этот вид характеризуется дихотомически ветвящимся в верхней части стеблем, наиболее крупными, длиной до 50 см, листьями с сердцевидным основанием, внезапно переходящим в черешок длиной 3-10 мм. Цветки у него белые, густо собраны в неравновершинную метелку длиной около 30 см, с короткими извилистыми и поникающими ветвями. Плоды розовые, длиной около 6 мм, имеют гладкий эндокарпий. Близкий вид - флагеллария гигантская (F. gigantea) - растет на островах Фиджи, Самоа, на архипелаге Бисмарка (остров Новая Ирландия) и, возможно, на Новой Гвинее. Отличается от фла-гелларии индийской более широкосердцевидным основанием листьев, рыхло расположенными цветками в длинной (до 50 см) равновершинной метелке с длинными и прямыми ветвями. Плоды длиной 12 мм, с бороздчатым эндокарпием. Третий вид - флагеллария гвинейская (F. guineensis, рис. 186) - эндемичен для западного и восточного побережий тропической Африки; в Западной Африке он известен от Берега Слоновой Кости до Южной Нигерии, а на восточном побережье - от прибрежных лесов Кении до Ист-Лондона на востоке Капской провинции ЮАР. Стебли у этой флагелларии не ветвятся. Листья у нее сидячие, длиной 10-20 см и шириной 1,5-3 см. Цветки желтые, собраны в неравновершинную метелку, длиной 6-10 см, с прямыми ветвями. Плоды ярко-красные.
Флагеллария, особенно азиатские виды, довольно широко используется местными жителями. Стебли ее идут на изготовление корзин как менее ценный заменитель ротанговой пальмы. Молодые стебли и ветви поедаются в сыром виде, а наряду с листьями употребляются для мытья волос. Листья и корни применяются как лекарственное средство. По сообщению Е. Христоферсена (1935), на острове Самоа стебли флагелларии иногда используют при сооружении домов в качестве опоры, к которой прикрепляется солома.
Семейство жуанвилеевые (Joinvilleaceae) (Т. Г. Леонова)
Монотипное пантропическое семейство жуанвилеевых состоит из одного рода жуанвилея (Joinvillea) и 2 видов, широко распространенных в тропических лесах Юго-Восточной Азии от острова Суматра через полуостров Малакка и северную часть острова Калимантан до островов Палаван и Холо (Филиппины), а также на Новых Гебридах, атолле Каролайн, Соломоновых островах, в Новой Каледонии, на Фиджи,, в Западном Самоа и на Гавайских островах.
Жуанвилеевые прежде объединялись с семейством флагеллариевых, но в 1970 г. английские ботаники А. Смит и П. Хомлинсон на основании морфологии и анатомии вегетативных органов, а также строения пыльцы предложили выделить жуанвилею в особое семейство, что было принято современными учеными (А. Л. Тахтаджян, 1980; А. Кронквист, 1981, и др.). Жуанвилеевые отличаются от флагеллариевых прежде всего прямостоячим, разделенным на узлы и междоузлия стеблем, складчатыми листьями без усиковидной верхушки.
Жуанвилеевые представляют собой многолетние травы с коротким и толстым симподиальным корневищем, покрытым чешуйками и несущим на своих концах по нескольку почек возобновления. Своим внешним обликом и некоторыми особенностями строения, как, например, эпидермальных клеток листа, устьиц и пыльцевых зерен, жуанвилеевые напоминают злаки. Стебли у них мощные и крепкие, прямостоячие, неветвистые, вальковатые или билатерально сплюснутые, голые, часто в числе нескольких, высотой 1,5-5,5 м и диаметром 0,4-1,4 см. Они густо облиствены, состоят из полых междоузлий длиной 2-14 см и заполненных тканями узлов (как у злаков) и не имеют вторичного утолщения. Нарастание стебля в высоту идет от его основания. Стебель почти полностью, как футляром, покрыт длинными (2-14 см), незамкнутыми, жесткими и голыми влагалищами листьев. Влагалища внезапно расширяются в пластинку листа, и в месте перехода наблюдаются маленький пленчатый язычок и 2 ушка (наподобие злаков). Ушки тоньше влагалищ, продолговатые, с закругленной или острой верхушкой, или они шиловидные, длиной 0,6-9 см. Пластинка листа линейно-ланцетная, дорсивентральная, на абаксиальной стороне вогнутая, имеет в длину 35-100 см и в ширину 4,5-20 см. Она цельнокрайная, складчатая благодаря наличию нескольких сильно выступающих с обеих сторон, крупных и густо расположенных жилок параллельного жилкования - наблюдается по 14-30 продольных "складок" на пластинке. В почкосложении пластинки листа также складчатые. На верхушке листья острые или довольно часто сужены в длинное остроконечие, но лишены усиков. Листья и стебли жуанвилеи грубые и жесткие, что вызвано обильным содержанием в них кремнезема. Листья, кроме того, с одной или обеих сторон густо покрыты крепкими щетинками и жесткими нитевидными волосками. В корнях же кремнезем образует значительные скопления в виде тел, заполняющих всю полость клеток, расположенных снаружи от механических волокон, окружающих тесно сближенные проводящие пучки. Устьица жуанвилеи парацитные (М. А. Баранова, 1975), замыкающие клетки их граминоидного типа. Сосуды с простой или лестничной перфорацией наблюдаются во всех органах растения. Секреторные клетки отсутствуют.
Цветки актиноморфные, мелкие и малозаметные, сидячие. Они обоеполые и трехчленные, снабжены ланцетовидными, рано опадающими пленчатыми прицветничками длиной 1-4 мм. Многочисленные и расставленные цветки собраны в конечные, широкие и пирамидальные метелки длиной 10-40 см. Метелки сильно ветвятся и имеют относительно тонкие, извилистые и отклоненные ветви, густо опушенные короткими и жесткими волосками. Соцветия снабжены рано опадающими треугольными или линейными пленчатыми кроющими листьями. Кроющие листья, окружающие главный цветонос, достигают в длину 25 см, прицветники на боковых ветвях - 0,5-3 см. Цветки сидят на утолщениях конечных веточек метелки. Околоцветник чашечковидный, голый, состоит из 6 пленчатых, черепитчато расположенных в 2 круга сегментов почти равной длины; они свободные или в основании коротко сросшиеся, по краю иногда мелкореснитчатые; наружные из них уплощенные, ланцетные или от яйцевидных до округло-яйцевидных, остроконечные или острые; внутренние - продолговатые или округло-яйцевидные, вогнутые. При плодах сегменты околоцветника сохраняются.
Тычинок 6, свободных, в 2 кругах; нити тонкие, длиной 2-4 мм; пыльники продолговатые, прикреплены к нитям своим основанием, открываются латрорзно продольной щелью; в основании они стреловидные, на верхушке вдавленные, во время цветения выступают из околоцветника. Пыльцевые зерна однопоровые, граминоидного типа. Гинецей синкарпный, из 3 плодолистиков; завязь верхняя, 3-гнездная, с 1 семязачатком в каждом гнезде. Столбиков 3, коротких, свободных или в основании сросшихся; рыльца линейные, перистые, довольно длинные, во время цветения выступают из околоцветника и сохраняются при плодах.
Цветут и плодоносят жуанвилеи одновременно. Опыляются они ветром перекрестно, но может произойти и самоопыление, благодаря тому что пыльники и рыльца созревают одновременно (гомогамия) и находятся в это время на одном уровне, превышая околоцветник. Кроме того, тычиночные нити и рыльца у них довольно слабые, так что под воздействием ветра они наклоняются друг к другу и пыльники при этом прикасаются к поверхности рылец, осуществляя автогамию. Т. Ньюел (1969) считает, что опылителями ее, кроме ветра, могут быть и пчелы, которых он находил на цветках этого растения.
Плоды костянковидные, нераскрывающиеся, более или менее трехгранные, они тускло- или ярко-красные, красно-оранжевые или коричнево-черные, содержат по 2-3 семени в каждом гнезде; экзокарпий плодов тонкий и мясистый, эндокарпий очень твердый и напоминает кость. Семена шаровидные или яйцевидные, диаметром 1,5-2 мм, с перепончатой оболочкой; они снабжены обильным мучнистым эндоспермом, окружающим маленький чечевицевидный зародыш. По мнению Г. Ридли (1930), семена распространяются птицами. Т. Ньюел (1969), однако, пишет, что он никогда не видел птиц на жуанвилее и не знает из сообщений в печати, чтобы их видел там кто-либо другой. С его точки зрения плоды жуанвилеи распространяются с помощью штормовых ветров. Плоды не имеют специальных приспособлений к анемохории, но они, как считает Ньюел, достаточно легкие для переноса ветром, дующим с большой силой. Семена хорошо прорастают в сухой период через 6 недель после созревания; в воде они сразу же тонут.
Обитают жуанвилеи в тропических дождевых лесах и светлых лесах на влажных, но не заболоченных почвах, а также по окраинам лесов, преимущественно в удаленных от моря районах, и в горах - на хребтах и водоразделах, где поднимаются до высоты 1600-1920 м над уровнем моря. Жуанвилея борнейская (J. borneensis) растет в северной части острова Суматра, на полуострове Малакка, на севере острова Калимантан и на Филиппинах (острова Палаван, Холо), возможно, на атолле Каролайн. Эта мощная трава встречается вдоль дорог, на окраинах лесов и в других довольно открытых местах с хорошо дренированной почвой, на высоте 50-1920 м над уровнем моря. Она часто является пионером в зарастании нарушенных человеком растительных сообществ. По облику она напоминает тростник. Листья ее на верхушке суживаются в длинное остроконечие, по краям, а часто и по обеим поверхностям они шероховатые от густо усаженных коротких щетинок. Близкий вид жуанвилея Годишо (J. gaudichaudiana) известен на Соломоновых островах, Новых Гебридах, Новой Каледонии, Фиджи, Западном Самоа, Гавайских островах и на атолле Каролайн. Это довольно редкое растение встречается во влажных лесах и затененных местах на горных хребтах и вдоль пересыхающих (временных) рек и потоков на высоте 300-1600 м над уровнем моря. Листья этой жуанвилеи снабжены редкими щетинками только на адаксиальной стороне; плоды имеют в диаметре 4,1-5,8 см, столбик при них не сохраняется.
Семейство рестиевые (Bestionaceae) (Н. Н. Цвелев)
В некоторых районах Южной Африки и Австралии место осоковых и злаков в растительных группировках занимают травянистые растения, по строению соцветий очень похожие на осоковые, но имеющие оригинальный внешний облик, который является следствием полной или почти полной редукции листовых пластинок и большой жесткости прутьевидных стеблей, взявших на себя функцию фотосинтеза и транспирации. Это представители семейства рестиевых, к которому принадлежит около 300 видов из 30 родов. Рестиевые особенно характерны для внетропических стран южного полушария и потому мало известны жителям северного полушария. Лишь один вид этого семейства - лептокарпус разъединенный (Leptocarpus disjunctus) встречается к северу от экватора - во Вьетнаме и Малайзии; другой вид этого же, преимущественно австралийского рода - лептокарпус чилийский (L. chilensis) является единственным представителем рестиевых в Америке (Чили и Южная Аргентина).
В пределах общего ареала семейства выделяют две области, наиболее богатые родами и видами рестиевых. Одна из них - внетропическая Австралия, включая Тасманию, где известны 13 эндемичных родов этого семейства. Лишь немногие виды рестиевых заходят на остров Чатам и в Новую Зеландию, во флоре которой имеется и один эндемичный род рестиевых - спорадантус (Sporadanthus). Второй основной областью распространения семейства является южная оконечность Африки, выделяемая, несмотря на небольшие размеры территории, в особое Капское флористическое царство. Здесь представлены 12 эндемичных родов рестиевых, и только немногие виды этого семейства заходят отсюда в тропическую Африку (до Малави) и на юг Мадагаскара. Наиболее богатый видами (около 125) род рестио (Bestio) встречается в обеих основных областях распространения семейства, однако в последнее время выявлено много существенных анатомических, палинологических и биохимических различий между южноафриканскими и австралийскими видами, и они, по-видимому, будут отнесены к разным родам.
Рестиевые - многолетние травянистые растения с длинными ползучими корневищами или короткокорневищные и тогда образующие густые дерновины. Корневища обычно несут кожистые чешуевидные листья, у основания которых по 1-2 отходят придаточные корни. Лишь у западноавстралийской дилъсии (Dielsia) корневища лишены чешуй, но покрыты густым буроватым войлоком, подобно корневищам некоторых папоротников. Надземные побеги большинства рестиевых представлены только у основания разветвленными жесткими цилиндрическими стеблями, несущими соцветия и обычно достигающими в высоту 25- 100 см. Лишь у немногих видов (например, у капской элегии мутовчатой - Elegia verticillaris) они достигают в высоту 2 м и в толщину 4 (8) см. Виды очень обособленного австралийского рода анартрия (Anarthria) имеют сильно сплюснутые с боков безлистные стрелкообразные стебли, а у капского рода антохортус (Anthochortus) они острочетырехгранные. Удлиненные междоузлия стеблей могут быть как полыми, так и заполненными сердцевиной. У видов некоторых родов, кроме неразветвленных репродуктивных побегов, имеются еще более или менее разветвленные вегетативные побеги (например, у рестио остроконечного - Bestio cuspidatus, рис. 188). У других видов стебли могут ветвиться во всех узлах, и в этом случае из пазух влагалищеобразных листьев могут выходить или пучки фертильных и стерильных ветвей, или только стерильные ветви. У элегии мутовчатой последние располагаются в узлах ложными мутовками и весь побег напоминает по облику побег гигантского хвоща или харовой водоросли.
Хорошо развитые мечевидные прикорневые листья, напоминающие листья ирисов, но более жесткие, имеются лишь у видов анартрии, например у анартрии шероховатой (Anarthria scabra, рис. 188). У других рестиевых листья обычно видоизменены в кожистые или перепончатые, долго сохраняющиеся на растении или опадающие влагалища, края которых обычно перекрываются, но, в отличие от влагалищ осоковых, не срастаются друг с другом. У некоторых родов (например, у австралийского онихосепалума - Onychosepalum) все узлы и расположенные на них влагалища тесно сближены у основания надземных побегов, но обычно на побегах имеется несколько расставленных узлов и влагалищ. Более или менее редуцированные листовые пластинки, обычно шиловидной формы и часто колючие на верхушке, есть у многих рестиевых, особенно на вегетативных ветвях. У рестио остроконечного (рис. 188) и некоторых других видов этого рода на границе влагалища с пластинкой расположен перепончатый вырост - язычок, сходный с язычком злаков и некоторых водных однодольных. По строению устьиц, которые здесь так называемого граминоидного (злакового) типа, рестиевые более сходны со злаками, чем с осоковыми. Расположение влагалищеобразных листьев на стеблях у рестиевых всегда двурядное, как у злаков.
Цветки рестиевых почти всегда однополые и двудомные, хотя рудимент гинецея в мужских цветках и стаминодии в женских нередко присутствуют. Мужские и женские цветки в одних и тех же соцветиях имеются только у монотипного капского рода филокомус (Phylocomns), а всегда обоеполые цветки - у австралийского вида лепиродия обоеполая (Lepyrodia hermaphroditа). Подобно злакам и осоковым, для рестиевых очень характерны элементарные соцветия - колоски, которые обычно, в свою очередь, собраны в колосовидные, кистевидные, метельчатые или зонтиковидные общие соцветия, имеющие у своего основания, а часто и у основания своих ветвей влагалищеобразные верхушечные листья. Внешне колоски представителей этого семейства особенно похожи на колоски некоторых осоковых, например видов камыша (Scirpus). Соцветия с мужскими и соцветия с женскими цветками в пределах одного и того же вида обычно настолько отличаются по внешнему виду, что их нетрудно принять за принадлежащие разным видам и даже родам. Женские соцветия меньше разветвлены, число цветков в женских колосках обычно уменьшено, а у ряда родов (например, харперия - Harperia, гиполена - Hypolaena и др.) они имеют только один цветок. Каждый цветок в колоске располагается в пазухе перепончатой или кожистой кроющей чешуи - прицветника, а у двух родов - анартрии и лепиродии - по бокам цветков имеются еще по 2 прицветничка. Самые нижние в колоске кроющие чешуи обычно не несут в своих пазухах цветков.
Околоцветник у рестиевых перепончатый или пленчатый, обычно состоящий из 4 или 6 расположенных 2 кругами бесцветных или слабоокрашенных сегментов более или менее одинакового строения. Сегменты внутреннего круга могут срастаться у своего основания. Кроме того, у некоторых видов один из сегментов, а иногда и все сегменты этого круга могут редуцироваться, и тогда околоцветник состоит из 3 или 5 сегментов. У австралийского рода локсокария (Loxocarya) околоцветник в женских цветках полностью отсутствует. У большинства родов семейства цветки актиноморфные или почти актиноморфные, однако у некоторых родов они становятся зигоморфными. Так, у капского тамнохортуса колосоноского (Thamnochortus spicigerus) боковые сегменты сросшегося при основании околоцветника сильно увеличены и снабжены килем, переходящим в широкий крыловидный вырост (рис. 188). У этого вида околоцветник сохраняется при плодах, играя роль летучки.
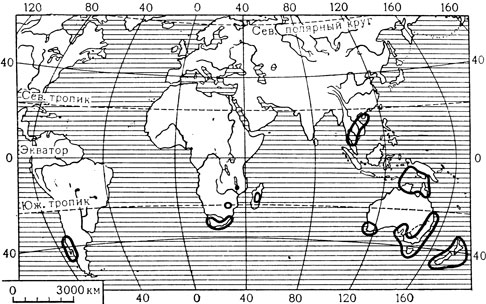
Карта 10. Ареал семейства рестиевых
Мужские цветки рестиевых имеют 3, реже 2 тычинки, расположенные супротивно внутренним сегментам околоцветника. Тычиночные нити обычно свободные, тонкие, с одногнездными, реже двугнездными (у примитивных и в других отношениях австралийских родов анартрии, лигинии - Lyginia и хопкинсии - Hopkinsia) пыльниками, прикрепленными к нити спинной стороной и вскрывающимися интрорзно поперечной щелью. Лишь у лигинии (рис. 188) нити тычинок срастаются в нижней части, образуя колонку. У этого последнего рода, как и у некоторых других родов, имеются надсвязники в виде шипиковидного выроста. Пыльцевые зерна рестиевых имеют однопоровую оболочку и обычно очень сходны с пыльцевыми зернами злаков.
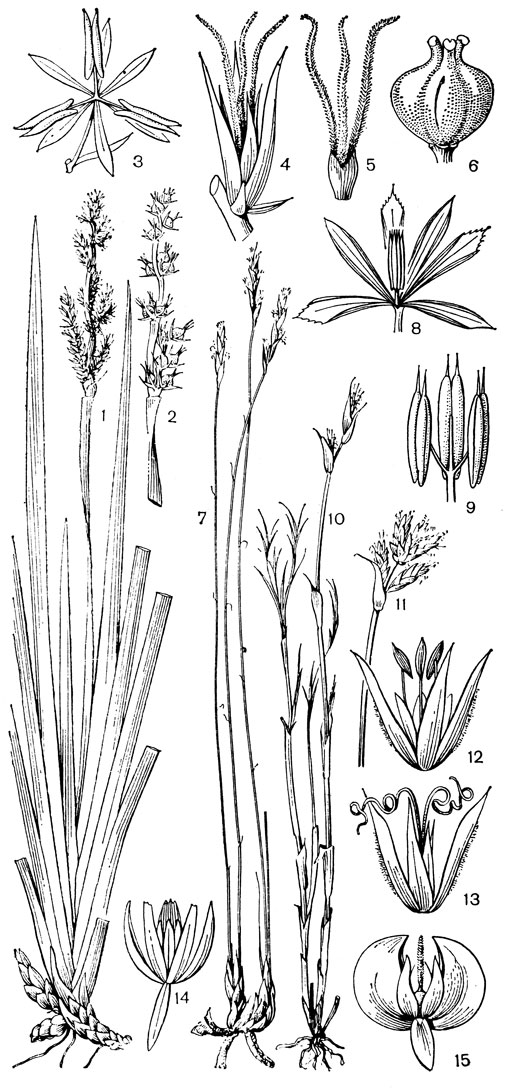
Рис. 188. Рестиевые. Анартрия шероховатая (Anarthria scabra): 1 - общий вид мужской особи; 2 - женское соцветие; 3 - мужской цветок; 4 - женский цветок; 5 - гинецей; 6 - плод. Лигиния бородатая (Lyginia barbata): 7 - общий вид мужской особи; 8 - мужской цветок; 9 - тычинки (их нити в нижней части сросшиеся). Реетио остроконечный (Restio cuspidatus): 10 - общий вид женской особи; 11 - мужское соцветие; 12 - мужской цветок; 13 - женский цветок. Тамнохортус колосоносный (Thamnochortus spicigerus): 14 - мужской цветок (один из сегментов наружного круга околоцветника отогнут); 15 - женский цветок (передний сегмент наружного круга околоцветника отогнут)
Синкарпный или паракарпный гинецей женских цветков рестиевых обычно состоит из 3 или 2 плодолистиков, лишь у родов тамнохортус и лепидоболус (Lepidobolus) он псевдомономерный в результате редукции переднего и одного из двух боковых гнезд первично-3- гнездной завязи. В двучленных женских цветках (например, у многочисленных видов рестио) два гнезда завязи располагаются супротивно двум наружным сегментам околоцветника. Ветви столбика обычно соответствуют числу гнезд завязи и в разной степени срастаются друг с другом. Волосистая рыльцевая поверхность находится на их внутренней стороне. Однако у некоторых родов гинецей имеет только один неразветвленный столбик, что связано или с редукцией двух гнезд завязи (у австралийского рода тамнохортус), или с полным срастанием всех ветвей столбика (например, у капской харперии). У гиподискуса (Hypodiscus) и вилъдеповии (Willdenowia) завязь, а позднее и плод располагаются на короткой дисковидной ножке, а у дильсии основание двухраздельного столбика пирамидообразно утолщено, как это бывает у некоторых родов осоковых. В каждом развитом гнезде гинецея имеется только один виеячий ортотропный семязачаток.
У большинства рестиевых плоды -2-3-семянные коробочки, раскрывающиеся продольной щелью по выступающим в виде ребер средним жилкам плодолистиков, т. е. локулицидно. Лишь немногие роды, например элегия и лепидоболус, имеют нераскрывающиеся орехообразные плоды с одним семенем. Семена всех рестиевых сходны по строению; они содержат обильный мучнистый эндосперм, у верхушки которого располагается маленький зародыш.
Большинство рестиевых обитает на бедных питательными веществами, обычно кислых, песчаных или торфянистых почвах. Это растения преимущественно влажных открытых местообитаний, однако более или менее пересыхающих во время сухого периода года. Среди капских видов много обитателей междюнных западин на морских побережьях, а некоторые австралийские виды могут расти на солончаках и даже в условиях настоящей пустыни при почти полном отсутствии осадков. Однако ксероморфный облик свойствен всем рестиевым, даже растущим на непересыхающих болотах, что, по-видимому, объясняется историческими причинами (предки рестиевых могли расти в еще более влажных условиях) или физиологической сухостью местообитаний.
Рестиевые являются ветроопыляемыми (анемофильными) растениями, причем возможность самоопыления у большинства видов исключается за счет двудомности - расположения мужских и женских цветков на разных особях. Специальных приспособлений для распространения семян у большинства рестиевых не отмечается. Выпадающие из коробочек семена могут переноситься ветром на некоторое расстояние от материнского растения, а затем разносятся или водными потоками, или на ногах животных вместе с комочками почвы. Лишь мелкие односемянные плоды рода тамнохортус, окутанные околоцветником с ширококрылатыми боковыми сегментами околоцветника, могут разноситься ветром на большие расстояния.
Система рестиевых пока еще недостаточно разработана, хотя их обычно делят на две группы по числу гнезд в пыльниках: двугнезднопылъниковые (Diplantherae) - более примитивная группа и одногнезднопылъниковые (Haplantherae), к которым принадлежит большинство родов семейства. В последнее время рестиевые, исключая выделяемый в особое монотипное семейство род экдейоколея (Ecdeiocolea), делят на 2 подсемейства: собственно рестиевые (Restionoideae) и анартриевые (Anarthrioideae), к которому принадлежит только оригинальный австралийский род анартрия (рис. 188), обладающий рядом примитивных признаков (хорошо развитые листья, присутствие прицветничков и др.).
Хозяйственное значение рестиевых незначительно. Их жесткие, безлистные или почти безлистные побеги не поедает или плохо поедает скот. Подобно тростнику, их можно использовать как покрытие для легких построек (особенно капский хондропеталум кровельный - Chondropetalum tectorum). В последнее время некоторые южноафриканские виды из родов элегия, хондропеталум, рестио и тамнохортус вводят в культуру в качестве декоративных растений, интересных своим своеобразным обликом.
Семейство экдейоколейные (Ecdeiocoleaceae) (Т. Г. Леонова)
Это небольшое семейство, состоящее из одного монотипного рода экдейоколея (Ecdeiocolea), эндемично для Юго-Западной Австралии. Прежде этот род включался в состав семейства рестиевых, но в 1965 г. английские ботаники Д. Катлер и Г. Эйри Шоу придали ему ранг самостоятельного семейства.
Экдейоколея одноколосая (Е. monostachya) представляет собой многолетнюю, похожую по облику на ситник, ксерофильную, голую траву с ползучим корневищем, покрытым чешуевидными влагалищами и многочисленными нитевидными придаточными корнями. Стебли у нее многочисленные, прямостоячие, крепкие, простые и тонкие, высотой от 40 до 60 см (редко до 1,5 м), внутри полые, типа соломины. Они серовато-зеленые, цилиндрические, в основании несколько утолщены и по всей длине имеют довольно глубокие продольные борозды, практически лишены листьев и выполняют функцию фотосинтеза. В основании стебли несут несколько коричневых, черепитчато расположенных, замкнутых листовых влагалищ длиной 7,5- 10 см, верхнее из которых удлиняется и поднимается почти до верхушки стебля. Листовая пластинка редуцирована до чешуйчатого или листовидного дорсивентрального зачатка, большею частью длиной 0,5-1 см и шириной от 2 мм и более. В стеблях экдейоколеи хорошо развита склеренхима в виде волокон и склереид, а также аэренхима. Во влагалищах листьев наблюдаются воздухоносные каналы, расположенные между проводящими пучками. Устьица парацитные, причем их замыкающие клетки граминоидного типа. Сосуды встречаются во всех органах, членики их с простой или преимущественно лестничной перфорацией. Соцветие расположено на конце стебля и имеет вид одиночного, густого, конического или цилиндрического колоса длиной большей частью 1,8-2,5 см и шириной 0,7 - 1 см. Колос состоит из многочисленных, густо черепитчато расположенных, твердых, темно-коричневых и блестящих чешуевидных брактей, или прицветников. Прицветники яйцевидные, тупые или закругленные, длиной около 5 мм, по виду напоминают колосковые чешуи злаков. В пазухе каждого прицветника, кроме 2-3 самых нижних, располагается по 1 сильно уплощенному цветку. Цветки однополые, собраны в однополые колосья, находящиеся на разных растениях. Экдейоколея считается двудомной, но английский ботаник Г. Вентам (1878) сообщает, что он находил в одном колосе как мужские, так и женские цветки; по-видимому, она может быть и однодомной. Околоцветник простой, чашечковидный, не превышает прицветника, состоит из 6 свободных, пленчатых, голых или близ верхушки слабоволосистых сегментов, расположенных в 2 круга. Наружные сегменты околоцветника уплощенно лодочковидно согнутые, широкие; внутренние - узкие и вогнутые. В мужских цветках имеются 3-4 тычинки, супротивные внутренним сегментам околоцветника; внешний круг тычинок почти полностью утрачен. Тычинки не превышают околоцветник; нити их тонкие, свободные, относительно короткие. Пыльники состоят из двух отдельных, не соединенных вместе гнезд, прикрепляющихся к тычиночной нити на середине спинки; они интрорзные, раскрываются продольной щелью. Пыльцевые зерна однопоровые, граминоидного типа. В мужских цветках имеется рудиментарная завязь, а в женских цветках наблюдаются 3 очень маленьких стаминодия. Гинецей синкарпный, состоит из 2 плодолистиков; завязь верхняя, 2-гнездная, содержит в каждом гнезде по 1 висячему семязачатку. Столбиков 2, коротких и свободных, переходящих в длинные рыльца. Экдейоколеи опыляются, вероятно, ветром. Ее плоды до сих пор неизвестны. Обитает экдейоколея в речных долинах.
Семейство центролеписовые (Centrolepidaceae) (Т. Г. Леонова)
Небольшое семейство центролеписовых объединяет 3 рода и около 35 видов, распространенных главным образом в Австралии, Тасмании и Новой Зеландии. Около 6 видов этого семейства распространено на Новой Гвинее, Зондских и Филиппинских островах, в Индокитае и на острове Хайнань, а 1 вид известен на юге Южной Америки (Огненная Земля) и на Фолклендских островах. Самый крупный и наиболее распространенный род центролепис (Centrolepis) насчитывает 26 видов, встречающихся почти по всему ареалу семейства, кроме Южной Америки. Наибольшее количество - 20 эндемичных видов этого рода - сосредоточено в Австралии и Тасмании, 3 вида - в Новой Зеландии и только 3 вида встречаются на остальной части ареала рода. Второй род - гаймардия (Gaimardia) - включает 2 вида, один из которых - гаймардия щетинолистная (G. setacea) - растет в Тасмании, Новой Зеландии и Новой Гвинее, второй вид - гаймардия южная (G. anstralis) - известен только на Фолклендских островах и Огненной Земле. Единственный вид третьего монотипного рода афелия (Aphelia) - афелия сытевидная (А. суperoides) - встречается в Южной Австралии и Тасмании.
Карта 11. Ареал семейства центролеписовых
Центролеписовые - мелкие многолетние или однолетние травы высотой до 10, реже 15, иногда всего 0,3-0,8 см. По внешнему облику они похожи на злаки, осоковые и ситниковые, а некоторые даже напоминают мхи. Однолетняя афелия сытевидная и однолетние виды центролеписа, например центролепис наименьший (Centrolepis minima), образуют небольшие пучкообразные розетки, состоящие из прикорневых листьев и одиночного безлистного стебля - цветочной стрелки, которая обычно выше листьев. Довольно часто цветущие стебли окрашены в ярко-красный цвет. У самых маленьких,- похожих на мох центролеписа карликового (С. hnmillima) из Западной Австралии и центролеписа Муррея (С. mnrrayi) из Южной Австралии стрелка отсутствует и сидячие соцветия полностью скрыты дуговидно изогнутыми шиловидными листьями длиной всего 3-8 мм. Многолетняя гаймардия, как и многолетние виды центролеписа, например новозеландские центролепис бледный (С. pallida) и центролепис реснитчатый (С. ciliata), а также широко распространенный в горах Австралии и ряда островов Малезии центролепис пучковатый (С. fascicularis) имеют многочисленные ветвистые, густо облиственные стебли, ветви которых могут укореняться. Эти виды образуют подушки диаметром обычно 10-30 см, аугаймардии щетинолистной - до 1 м. Увеличение размера подушек идет за счет центробежного роста стеблей. При этом центральная, более старая часть подушки может отмирать и разрушаться, в результате чего подушка приобретает необычную форму кольца (например, у центролеписа пучковатого и центролеписа филиппинского - С. philippinensis), напоминающего "ведьмины кольца" некоторых европейских грибов и плаунов.
Листья центролеписовых узколинейные, обычно нитевидные, щетиновидные или шиловидные, длиной 0,3-6,7 см и шириной 0,1-0,7 см. Они довольно жесткие, вальковатые, сплюснутые или желобчатые, голые или опушенные белыми многоклеточными волосками. На верхушке листья могут быть тупыми или острыми, иногда с длинным прозрачным острием. При основании они расширены в хорошо заметное открытое перепончатое влагалище, иногда снабженное в месте перехода в пластинку язычком или ушками. У однолетних видов все листья прикорневые и собраны в виде розетки. У многолетних видов они очередные и довольно густо расположены на стеблях. Устьица парацитные, причем замыкающие их клетки граминоидного типа. Сосуды имеются во всех органах растения; членики их с лестничной перфорацией.
Цветки центролеписовых однополые и однодомные. Они очень мелкие, сидячие, лишены околоцветника. Мужские цветки представлены одной тычинкой с длинной нитью и продолговатым или линейным одногнездным пыльником. Пыльники прикреплены к нити спинной стороной, обычно близ ее середины (качающиеся), но у афелии - почти у основания пыльника; раскрываются они интрорзно продольной щелью. Пыльцевые зерна имеют однопоровую оболочку и обычно граминоидного типа. Строение женских цветков еще недостаточно изучено. Гинецей, по-видимому, псевдомономерный. Завязь 1-гнездная, со столбиком, заканчивающимся часто ярко-красным (у центролеписа) или желтоватым (у афелии) односторонним линейным рыльцем. Однако у многих центролеписовых в результате срастания завязей двух или трех соседних женских цветков может образоваться сложная "завязь" с 2-3 гнездами; столбики при этом могут также срастаться друг с другом в нижней части или оставаться свободными, причем их рыльца иногда скручиваются друг с другом или завиваются в виде спирали. В 1-гнездной завязи или в каждом гнезде такой сложной "завязи" имеется только один висячий ортотропный семязачаток. Цветки центролеписовых собраны в обоеполые, реже однополые парциальные (элементарные) соцветия, расположенные в пазухах супротивных или очередных чешуевидных кроющих листьев. Эти элементарные соцветия, в свою очередь, собраны в общее (сложное) соцветие, которое у видов центролеписа имеет форму головки, а у гаймардии и афелии колосовидное. Наиболее сложно устроены соцветия у центролеписов (рис. 189, 190). Их головчатое соцветие в основании окружено двумя, редко тремя почти супротивными, неравными по величине, линейными, яйцевидными или обратнояйцевидными кроющими листьями длиной 2-5 мм. На верхушке они тупые или с длинным, иногда по килю зазубренным остевидным окончанием. В пазухе каждого такого кроющего листа располагаются по 1-8, редко до 20 элементарных соцветий, обычно снабженных бахромчатыми чешуевидными прицветниками (редко без них). В редких случаях самый нижний кроющий лист лишен элементарных соцветий. У центролеписа каждое такое соцветие состоит из одного (иногда отсутствующего) мужского цветка и 1-20 (обычно более одного) женских цветков. Женские цветки располагаются в 1-2 ряда друг над другом или спирально, прирастая боковой частью завязи к оси соцветия. У центролеписа карликового в основании головки имеется только один кроющий лист, в пазухе которого располагается одно редуцированное до 1 мужского и одного женского цветка элементарное соцветие. На оси колосовидного общего соцветия афелии такие соцветия располагаются по одному в пазухах кожистых, тесно сближенных, очередных, в основании расширенных кроющих листьев. Самые нижние 1-2 кроющих листа голые, отличаются от остальных большими размерами и длиннозаостренной верхушкой. В пазухе каждого из них развивается по одному однополому элементарному соцветию, состоящему из 1 или 3 мужских цветков. Остальные кроющие листья (их 5-16) зеленые, жестко- и коротко-волосистые, на верхушке острые. Они заключают или по одному однополому элементарному соцветию, редуцированному до одного женского цветка, или по одному обоеполому соцветию, состоящему из одного мужского и одного женского цветка. Каждое такое соцветие у афелии обычно снабжено 1-2 пленчатыми чешуевидными прицветниками.
У гаймардии (рис. 189) общее колосовидное соцветие более упрощено, и на его оси имеется всего 3 очередных, последовательно уменьшающихся (длиной от 3 до 1 мм), чешуевидных, кроющих листа, в пазухе каждого из которых помещается по одному обоеполому элементарному соцветию. У гаймардии щетинолистной чаще всего верхний и реже средний кроющие листья лишены элементарных соцветий, а у гаймардии южной во всем общем соцветии имеется только 1 элементарное соцветие, находящееся в пазухе нижнего кроющего листа. Элементарное соцветие гаймардии содержит 2 свободных мужских цветка и 2 женских цветка, полностью сросшихся завязями друг с другом. Плоды центролеписовых мелкие, сухие, с пленчатым околоплодником (перикарпием), раскрывающиеся продольно наружной
щелью или двумя створками (у гаймардии южной). Семена длиной 0,5-1 мм, эллипсоидальные, яйцевидные или обратнояйцевидные, с ячеистой поверхностью. В семенах имеются обильный мучнистый эндосперм и маленький обратноконический или щитковидный зародыш.
Центролеписовые обитают преимущественно в субальпийском или альпийском поясе гор на высоте от 2000 до 4000 м над уровнем моря, реже в низкогорьях и на равнинах. Обычно они встречаются группами на открытых и более или менее влажных местообитаниях: скалах, болотах, болотистых лужайках, песчаных и глинистых склонах, берегах водоемов (в том числе морских лагун), иногда на рисовых полях. Некоторые виды центролеписа растут в песчаных пустынях Австралии.
Биологические особенности центролеписовых еще недостаточно изучены. Все они - ветроопыляемые растения, хотя, вероятно, могут и самоопыляться. Одни центролеписы начинают цвести в сентябре, другие - в декабре - марте и плодоносят, соответственно, в ноябре или с января по апрель. Афелия цветет в октябре - декабре, гаймардия - в ноябре - декабре.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'