Порядок коммелиновые (Commelinales) (В. И. Трифонова)
Семейство рапатеевые (Rapateaceae)
Относительно небольшое семейство рапатеевых насчитывает 16 родов и около 100 видов, распространенных главным образом в тропической Южной Америке. Наиболее полно и разнообразно оно представлено на Гвианском нагорье, где сосредоточена большая часть видов. Лишь представители 2 родов встречаются за его пределами: род эпидриос (Epidryos) распространен в тропиках Южной и Центральной Америки, и монотипный род машалоцефалус (Maschalocephalus) - в тропической Западной Африке,на территории Либерии и Сьерра-Леоне.
Рапатеевые - часто очень высокие (до 3 м), многолетние, похожие на осоковые травы с коротким, довольно мясистым корневищем и двурядными, приземными листьями. Листья узкие, от линейных до мечевидных, со слабо выраженными параллельными жилками. Пластинка листа, как правило, повернута на 90° от хорошо развитого, открытого и сложенного, как правило, вдоль средней жилки влагалища и соединена с ним с помощью черешка (рис. 176). В основании листьев нередко скапливается желатин. Устьица, разных типов. Сосуды обычно имеются во всех вегетативных органах, у большинства они с лестничной и лишь иногда с простой перфорацией. Цветки обоеполые, актиноморфные, собраны в верхушечные головчатые или однобококолосковые соцветия, поднятые на цветоносах. Лишь у рода машалоцефалус цветонос короткий и соцветие почти полностью скрыто в пазухе приземных листьев или слегка выдается из нее наружу. Рапатеевые замечательны наличием у них обертки разнообразной формы из 1-2 или многих кроющих листьев (прицветников), окружающих соцветие. Редко ее нет совсем, как у эпифитного рода эпидриос, или она сохранилась в виде остаточных, узких, пленчатых чешуек, как у многих видов стеголеписа (Stegolepis). У большинства рапатеевых она состоит из 2 расположенных друг против друга, очень крупных или небольших по размеру, кроющих листьев, свободных, как у видов рода рапатея (Rapatea), или полностью сросшихся краями, как у видов рода саксофридериция (Saxofridericia). У представителей рода спатантус (Spathanthus), обертка которого состоит из одного крупного кроющего листа, соцветие удивительно похоже на соцветие аронниковых. Каждый цветок снабжен многочисленными черепитчатыми, маленькими или более крупными и тогда полностью закрывающими цветок, иногда ярко окрашенными прицветничками. Трехчленный околоцветник состоит из четко различимых чашечки и венчика. Твердые, прозрачно-пленчатые, черепитчатые чашелистики свободны или у основания более или менее срастаются в короткую трубку с широко отогнутыми овальными лопастями. Тычинок 6, расположенных в 2 круга, как правило, более коротких, чем лепестки, и становящихся видными, только когда те увянут. Лишь у машалоцефалуса тычинки длиннее венчика и торчат наружу из цветка. Нити очень короткие, у основания сросшиеся друг с другом или свободные и приросшие к трубке венчика. Пыльники крупные, прикрепляются к нитям основаниями. При созревании пыльники открываются 1-2 или 4 верхушечными порами или короткой, верхушечной щелью (рис. 176). Гинецей синкарпный, с длинным столбиком, заканчивающимся маленьким рыльцем. Завязь верхняя, 3-гнездная, с 1-2 или несколькими анатропными семязачатками в каждом гнезде. Плод - локулицидная коробочка. Семена с маленьким линзовидным зародышем, расположенным близ рубчика и с обильным мучнистым эндоспермом. Часто они с верхушечным выростом (рис. 176). Оболочка семени с продолговатыми бороздками или покрыта маленькими колючками. Характерной особенностью рапатеевых является их способность накапливать алюминий. В отличие от коммелиновых у них нет кристаллов оксалата кальция.
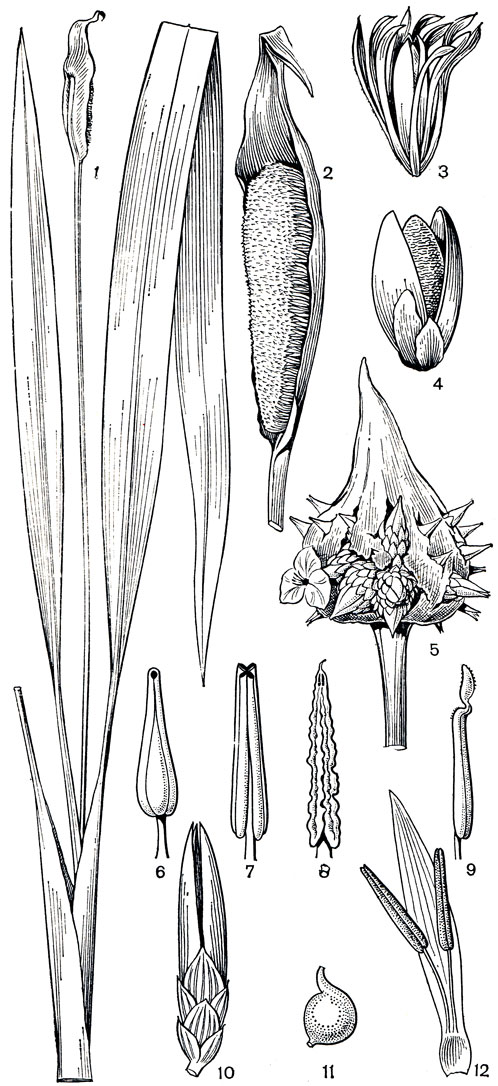
Рис. 176. Рапатеевые. Спатантус односторонний (Spathanthus unilateralism 1 - общий вид; 2 - соцветие с одним кроющим листом, 3 - бутон, окруженный прицветничками; 4 - раскрытый плод. Саксофридериция большая (Saxofridericia grandis): 5 - соцветие. Виндзорина гвианская (Windsorina guianensis): 6 - тычинка. Схеноцефалиум клобучков ый (Schoenocephalium cucullatum): 7 - тычинка. Саксофридериция губчатая (Saxofridericia spongiosa): 8 - тычинка. Рапатея длинноножковая (Rapatea longipes): 9 - тычинка. Схеноцефалиум Мартиуса (Schoenocephalium martianum): 10 - цветок с прицветничками. Спатантус односторонний: 11 - семя с верхушечным выростом. Схеноцефалиум Мартиуса (Schoenocephalium martianum): 12 - лепесток с двумя приросшими к нему тычинками
Рапатеевые составляют важный элемент растительного покрова Гвианского нагорья. Заросли этих высоких и часто довольно мощных трав придают характерный облик его массивам. Большинство рапатеевых приурочено к более или менее влажным местообитаниям. Они растут на горных хребтах, поднимаясь до высоты 1700-2000 м над уровнем моря. Здесь они часто встречаются среди кустарниковых зарослей, в разного рода редколесьях и саваннах, по берегам рек и водных потоков, в периодически затопляемых бамбуковых и пальмовых рощах, на болотах и топях в дождевых низинных лесах. Интересной жизненной формой рапатеевых тропического леса являются эпифитные травы из рода эпидриос. Эти растения с крупными, мясистыми корневищами и слабыми, нитевидными, пониклыми цветоносами селятся в кронах высоких деревьев. Все 3 вида узкоэндемичны, и ареалы их не пересекаются друг с другом. Эпидриос Аллена (Epidryos allenii), внешне напоминающий ирис, найден в тропических лесах холмистой части Панамы, расположенных на высоте 1000 м над уровнем моря, где он вместе с орхидными и папоротниками растет в кронах высоких, десятиметровых, деревьев. Эпидриос мелкопылъниковый (Е. micrantherus) известен только из занимающих более низкие высоты (около 70 м над уровнем моря) дождевых, с большим количеством осадков, тропических лесов Колумбии. Третий вид этого рода - эпидриос гайанский (Е. guayanensis) - был обнаружен в лесах Гайаны совсем недавно, в 1960 г. Несколько позднее он был найден также в соседней Венесуэле, где эти растения образуют довольно большие группы, свисающие с деревьев, растущих по берегам рек и потоков.
Рапатеевые - энтомофильные растения. Цветки их лишены нектара и привлекают насекомых ярко окрашенными лепестками, а нередко и яркими прицветничками и кроющими листьями обертки. Растущая довольно большими группами на заболоченных почвах у водоемов или в низменных лесах рапатея перепончатая (Rapatea membranacea) с небольшими ярко-желтыми цветками посещается маленькими пчелами. У многих рапатеевых цветение не зависит от времени года. Так, например, в популяциях машалоцефалуса цветущие растения можно найти круглый год. Цветки рапатеевых эфемерны: едва распустившись, они увядают. Лепестки темнеют и скручиваются, открывая тычинки с крупными, пустыми, красновато-коричневыми пыльниками. С эфемерностью цветков, вероятно, связано почти полное отсутствие их в гербарии. Редко цветущие растения удается наблюдать и в природе. Так, Г. Глисон (1923) среди десятков и даже сотен растений спатантуса одностороннего (Spathanthus unilateralis) и рапатеи болотной (Rapatea paludosa, рис. 177), растущих в Гвиане, нашел только один экземпляр с распустившимися цветками. Для некоторых рапатеевых, вероятно, характерна клейстогамия. Так, у представителей рода схеноцефалиум (Schoenocephalium) маленькие, почти полностью закрытые прицветничками цветки не открываются совсем. Не открываются чашелистики и у некоторых видов рода стеголепис.
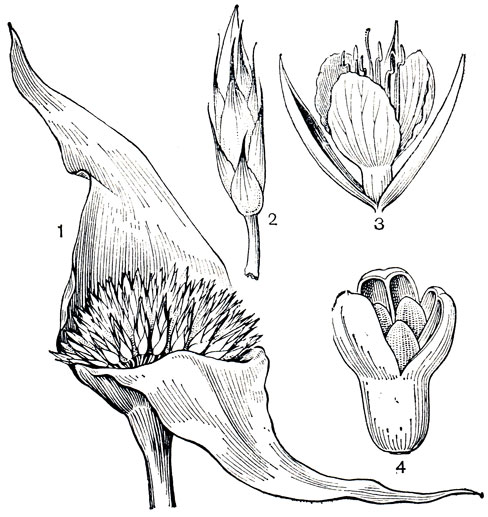
Рис. 177. Рапатея болотная (Rapatea palludosa): 1 - соцветие; 2 - цветок с прицветничками; 3 - цветок; 4 - раскрытый плод
Распространяются рапатеевые семенами и вегетативно с помощью корневых отпрысков. Многочисленные дерновинки фелпсиеллы стеблекрылой (Phelpsiella pterocaulis) - невысокой травы с желтыми цветками, найденной только на открытых вершинах гор в Венесуэле, распространились, вероятно, таким образом. В отраженном свете листья ее приобретают голубоватый оттенок, что характерно и для других рапатеевых. У некоторых представителей этого семейства наблюдается образование луковичек, как, например, в соцветии стеголеписа живородящего (Stegolepis vivipara). Семена у него, как правило, не развиваются, и растение распространяется с помощью многочисленных луковичек, опадающих на землю и прорастающих рядом с материнским растением.
Американский ботаник В. Магуайр (1958), основываясь прежде всего на числе семязачатков, способе прикрепления их к плаценте и форме семени, разделил все семейство на два подсемейства: саксофридерициевые (Saxofridericioideae) и рапатеевые (Rapateoideae). Позднее эта система была в основном подтверждена анатомическими и палинологическими исследованиями Ш. Карлквиста (1961, 1966). В подсемействе саксофридерициевых В. Магуайр выделяет 2 трибы: саксофридерициевые (Saxofridericieae) и схеноцефаловые (Schoenocephalieae).
Первая триба объединяет растения с желтыми, обратнояйцевидными или обратносердцевидными лепестками, заметно выступающими из цветка. К роду саксофридериция относятся 9 видов, представленных часто довольно мощными, высотой до 3 м, многолетними травами. Их единственное соцветие, как правило, полностью закрыто двумя сросшимися краями кроющими 4листьями, имеет форму луковицы. Развивающиеся бутоны прорывают обертку, вынося раскрывающиеся цветки наружу. Лишь у саксофридериции королевской (Saxofridericia regalis) кроющие листья многочисленные, небольшие, наружные длиной до 5 см, пленчатые. Лепестки у саксофридериций ноготковые и срастаются в пленчатую, хорошо различимую трубку только во время цветения. Представители этого рода приурочены к двум различным местообитаниям. Часть из них, относимая к подроду эвсаксофридериция (Eusaxofridericia), ограничена горными районами Гвианского нагорья, где они растут на поросших кустарниками и лесами склонах на высоте 1200-2000 м над уровнем моря. Лишь саксофридериция губчатая (Saxofridericia spongiosa) спускается до высоты 120-200 м над уровнем моря. Эти высокие травы с красивыми, длиной до 3,5 м листьями и крупными ярко-желтыми соцветиями придают характерный облик открытым гранитным откосам. Представители подрода акротека (Acrotheca) встречаются на более низких высотах, где растут на болотах и топях в периодически затопляемых тропических дождевых лесах. К трибе саксофридерициевых относится и самый многочисленный род семейства - стеголепис, насчитывающий 23 вида. Это растения болотистых местообитаний и влажных склонов, встречаются обычно на высоте 1500-2000 м над уровнем моря на песчаниках Гвианского плоскогорья. Более редки они в открытых лесистых местностях. Большинство стеголеписов - наземные растения, но есть среди них и эпифиты. Так, например, стеголепис Штейермарка (S. steyermarkii) или растет на земле, или селится на нижней части стволов крупных тропических деревьев. К этому роду, несомненно, близок полностью эпифитный род эпидриос.
К трибе схеноцефаловых отнесены растения с ярко-красными или красноватыми, заключенными в чашечку лепестками. Лишь у монотипного рода кунгардтия (Kunhardtia) они выступают наружу. Это растение обычно для открытых, поросших кустарником горных террас и саванн, расположенных на высоте 1500-2000 м над уровнем моря. Кунгардтия отличается от остальных членов трибы формой соцветия, вытянутыми лопастями венчика и числом гнезд пыльников. Род схеноцефалиум представлен довольно большими (высотой до 1,5 м и более) травами с красивыми, крупными, ярко окрашенными соцветиями. Растут они на каменистых и песчаных почвах, где доминируют среди остальной растительности. Встречаются схеноцефалиумы нередко и в живописных расщелинах скал и на болотах в долинах рек Ориноко и Риу-Негру.
К подсемейству рапатеевых отнесены растения с одним семязачатком в каждом гнезде завязи и продолговатыми овальными семенами. В трибу рапатеевых (Rapateae) объединены растения с крупными, значительно превышающими соцветие, кроющими листьями и продолговато-бороздчатыми семенами с верхушечным сосочко-колпачковым выростом или без него. Второй по объему род семейства - рапатея - насчитывает почти два десятка видов и имеет самый большой ареал. Эти многолетние травы, внешне напоминающие некоторые осоковые, распространены в Колумбии и Венесуэле и далее на юг - в Перу и Бразилии. Растут рапатеи в болотистых и топких низинах среди дождевых лесов, обычно на небольших высотах - до 700 м над уровнем моря.
Последнее место в системе В. Магуайра занимает триба монотремовых (Monotremeae), наиболее специализированная по ряду анатомических и морфологических признаков. Кроющие листья у растений этой трибы, как правило, более или менее равны или короче соцветия. На примере этой трибы можно проследить эволюцию сосудов в семействе рапатеевых.
Для монотремовых характерно наличие в узких сосудах корней еще лестничной перфорации, а в широких сосудах корней, так же как и в сосудах стеблей, уже только простой перфорации.
К этой трибе принадлежит единственный представитель рапатеевых Африканского континента - род машалоцефалус. Его виды - характерные элементы флоры большей части прибрежной полосы Либерии и растут везде, где находят подходящие для себя условия. Это растение с длинными корнями часто встречается на болотах, во влажных саванновых лесах, затопляемых в период дождей. Более редко он встречается только в отдаленных от моря районах.
Хотя это растение обильно представлено в Либерии, хозяйственная деятельность человека, направленная на осушение болот и изменяющая экологические условия, наносит ему заметный ущерб, вызывая гибель нередко целых популяций. Найден машалоцефалус и в Сьерра-Леоне, где известно пока только одно его местонахождение.
Практическое значение рапатеевых невелико, хотя многие из них весьма декоративны и привлекают внимание ярко окрашенными (желтыми, красными, пурпурными) соцветиями и сочной зеленью листьев. Особенно хороши виды рода схеноцефалиума с нежно-розовыми снизу и белоснежными в верхней части, становящимися пурпурными во время цветения прицветничками.
В Колумбии эти довольно редкие растения являются предметом выгодной торговли. На рынках Боготы их продают под названием "маленькие звезды с юга". Причудливо окрашенные в красные тона, соцветия некоторых рапатеевых сходны с оперением попугаев ара.
Семейство ксирисовые (Xyridaceae)
Семейство ксирисовых, представленное 4 родами и 270 видами, распространено в тропических и субтропических областях Америки, Африки, Южной и Юго-Восточной Азии, Новой Каледонии, Австралии и Тасмании.
Ксирисовые - невысокие, большей частью многолетние, редко однолетние, внешне похожие на ситниковые травы с волокнистыми, как правило, корнями и приземными двурядными или собранными в розетку листьями. Есть среди них и растения с короткими, утолщенными, иногда ползучими корневищами или луковицами, как, например, у североамериканского ксириса скрученного (Xyris torta). Узкие линейные или нитевидные листья в нижней части переходят во влагалище, вход в которое иногда бывает прикрыт небольшим язычком. Устьица парацитные (ксирис) или аномоцитные (аболъбода - Abolboda). Сосуды имеются во всех вегетативных органах: в корнях, стеблях и листьях. Все они с простой перфорацией, что говорит об определенной эволюционной продвинутости этого семейства.
Ксирисовые - растения главным образом болотистых местообитаний. Обычны они на песчаных кислых и засоленных почвах, на выходах горных пород, где нередко образуют довольно плотные заросли вместе с осоками, росянками, эриокаулонами и ситниками. Растут они и на сфагновых болотах, на торфяниках, вдоль потоков, канав, по берегам рек и озер. Большинство из них растения низин, реже их можно встретить на заболоченных участках в горах, где они могут расти на высоте до 3000 м над уровнем моря.
Обоеполые, 3-членные, актиноморфные или слегка зигоморфные цветки ксирисовых собраны в верхушечные шарообразные или цилиндрические головки, высоко поднятые над землей на цветоносах (рис. 178, 2). Цветоносы цилиндрические или сжатые по бокам, иногда крылатые или ребристые, гладкие или покрытые многочисленными сосочками. Каждый отдельный цветок сидит в пазухе небольшого кожистого или жесткого прицветника слегка вогнутой формы. Прицветники располагаются по спирали, черепитчато налегая друг на друга. Нижние нередко стерильны и формируют обертку соцветия. Чашелистики пленчатые, их, как правило, 3, редко только 2, как у большинства представителей южноамериканского рода абольбода. Два наружных боковых чашелистика лодочковидные, с килем посередине, обычно неопадающие, а третий, внутренний, образует капюшон над венчиком и позднее, по мере развития лепестков, выталкивается ими наружу и опадает. Венчик из 3 желтых, реже голубых или белых лепестков. Лепестки ноготковые, срастаются в короткую или длинную трубку с тремя, как правило, равными и широко отогнутыми лопастями. Зигоморфный венчик характерен только для представителей южноамериканского рода оректанта (Orectanthe). Тычинок 3, приросших к лепесткам, нити их короткие, пыльники раскрываются продольными щелями (рис. 178, 4). У ксирисов фертильные тычинки чередуются с тремя кисточковидными, часто раздвоенными стаминодиями (рис. 178, 5). Нет стаминодиев у монотипного венесуэльского рода ахлифила (Achlyphila) и у оректанты. Среди абольбод встречаются виды как с простыми нитевидными стаминодиями, так и без них. Гинецей паракарпный или у основания 3-гнездный, со столбиком, заканчивающимся простым или 3-лопастным рыльцем (рис. 178, 6). У абольбоды и оректанты столбик с придатком в середине или у основания (рис. 178, 7). Завязь верхняя, 1-гнездная или у основания неясно 3-гнездная, редко явно 3-гнездная, со свободной центральной плацентой (неотропическая секция нематопус - Nematopus рода ксирис). Семязачатков несколько или много, редко 1, ортотропных (ксирис) или слегка кампилотропных или анатропных.
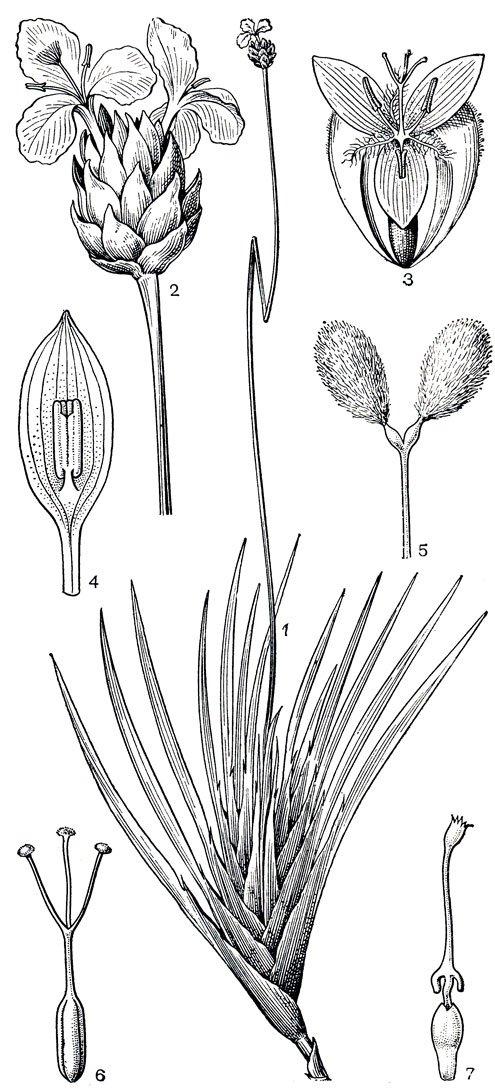
Рис. 178. Ксирисовые. Ксирис витсеноидный (Xyris witsenioides): 1 - общий вид. Абольбода поаршон (Abolboda роаrchon); 2 - соцветие. Ксирис изорванный (Xyris lacerata): 3 - цветок с двумя боковыми чашелистиками (третий, внутренний лепесток удален). Ксирис реснитчатолистный (X. blepharophylla): 4 - лепесток с приросшей к нему тычинкой; 5 -стаминодий; 6 - гинецей. Абольбода Пеппига (A. poeppigii): 7 - гинецей с придатком
Плод - локулицидная коробочка, открывающаяся тремя створками и иногда завернутая в остающийся при плодах венчик. Семена мелкие, продолговато-веретеновидные или шарообразные, с обильным мучнистым или мясистым эндоспермом и маленьким верхушечным зародышем. Оболочка семени обычно с продолговатыми бороздками, реже без них и тогда семена сильно сжатые и ширококрылатые, как у представителей рода оректанта. Созревшие коробочки, растрескиваясь, высыпают многочисленные легкие семена, которые распространяются при помощи ветра и воды. Цветки ксирисовых недолговечны: раскрываясь ранним утром, они вскоре уже опадают.
Семейство ксирисовых довольно неоднородно по составу. Два южноамериканских рода - абольбода и оректанта - некоторыми ботаниками выделяются в самостоятельное семейство аболъбодовые (Abolbodaceae). Эти роды отличаются, как правило, спиральным расположением листьев, более удаленными прицветниками, равными по размеру чашелистиками, сросшимися голубыми или белыми лепестками, безапертурной оболочкой пыльцевых зерен, столбиком с простым рыльцем и придатком в середине или у основания, отсутствием стаминодиев, 3-гнездной завязью с пазушными плацентами, анатропными семязачатками и сжатыми или призматическими Семенами с большим согнутым зародышем.
Самый многочисленный род семейства ксирис (Xyris) насчитывает более 250 видов, широко представленных в субтропических и тропических областях всего земного шара. Некоторые из них, как, например, ксирис капский (X. саpensis) и ксирис индийский (X. indica), распространены от Юго-Восточной Азии до Австралии, но большинство имеют довольно ограниченный ареал. Как правило, это растения низин, растущие по краям мелких болот и топей. Только очень немногие, как, например, ксирис веерообразный (X. flabellata), ксирис большой (X. grandis) и ксирис горолюбивый (X. оrеоphila), поднимаются достаточно высоко в горы, достигая 3000 м над уровнем моря. Одиночные виды, например ксирис индийский, который имеет, вероятно, антропогенное распространение, обычны на заброшенных рисовых полях. Вместе с ксирисом немногоцветковым (X. pauciflora) они растут также в сухих листопадных лесах на высоте 50-100 м над уровнем моря. В Колумбии ксирис каролинский (X. caroliniana) растет среди сфагновых болот и в заболоченных магнолиевых лесах. В Таиланде ксирисы можно найти в болотистых осоковых лугах и на пастбищах, по берегам рек и небольших потоков, вдоль влажных тропинок и в открытых травянистых сосновых лесах на высоте 700-1500 м над уровнем моря. Нередки они и среди мхов на открытых песчаных плато. В Либерии ксирисы обычны как в прибрежной полосе, так и в удаленных от океана районах, где они часто встречаются на выходах горных пород. Обычны они и вдоль потоков и на засоленных почвах низин. Ксирис обманчивый (X. decipiens) образует на открытых заболоченных участках саванн характерные ассоциации вместе с мезантемумом укореняющимся (Mesanthemum radicans).
Род ксирис делится на 3 основные секции. К самой маленькой по объему секции поматоксирис (Pomatoxyris), насчитывающей, вероятно не более 20 видов, отнесены австралийские растения с базальной 3-гнездной коробочкой. К пантропической, доминирующей в Северной Америке секции ксирис (Xyris) отнесено более 100 видов с 1-гнездной коробочкой и париетальной плацентацией. В секцию нематопус (Nematopus) включено большинство неотропических видов, число которых превышает полторы сотни. Это растения также с 1-гнездной коробочкой, но со свободной центральной пла-центой. В 1954 г. Р. Ван Ройен описал новый вид ксириса - ксирис черноостроконечный (X. nigromucronata). Это растение, найденное в Австралии среди зарослей кустарников, настолько отличается от остальных известных видов, что было выделено им в особую секцию австралоксирис (Australoxyris). Прежде всего бросается в глаза наличие 6 лепестков с приросшими к ним 6 тычинками, ярковыраженная бахрома наверху листового влагалища, сросшиеся боковые чашелистики и столбик с простым рыльцем. Отличаются они и 3-гнездной при основании и 1-гнездной в верхней части завязью с более или менее кампилотропными семязачатками. Сросшиеся боковые чашелистики были обнаружены также у растущего в Бразилии ксириса туповатого (X. obtusiuscula), а 6 тычинок найдены были автором у малоазиатского ксириса банкийского (X. baricana).
Второй по численности род семейства абольбода (Abolboda) насчитывает почти 20 видов, полностью ограниченных тропической зоной Южной Америки; нет их только в Андах. Это растения с голубыми или белыми, большей частью актиноморфными цветками, с 2 чашелистиками и столбиком, снабженным 3 отогнутыми вниз небольшими выростами (рис. 178, 7) и с простыми, часто нитевидными стаминодиями или без них.
Близкий к абольбодам род оректанта (Оrесtanthe) отличается отсутствием стаминодиев, формой рыльца, сильно сжатыми крылатыми семенами и сильно вытянутыми лепестками, за форму которых он и получил свое название.
Найденная в горах на юге Венесуэлы ахлифила (Achlyphila) представляет собой многолетнюю траву с горизонтальными корневищами и хорошо развитыми двурядными стеблевыми листьями. Встречаются ахлифилы среди кустарниковой растительности на высоте 2000 м над уровнем моря, на юго-восточных склонах гор, почти всегда закрытых облаками.
Практического значения ксирисовые не имеют, хотя местные жители используют некоторые виды ксирисов для приготовления лекарственных препаратов.
Семейство коммелиновые (Commelinaceae)
Семейство коммелиновых насчитывает 47 родов и около 700 видов, распространенных в тропических и субтропических областях обоих полушарий. Лишь немногие из них заходят в умеренно теплые области Восточной Азии и США. В Европе коммелиновые представлены лишь несколькими одичавшими видами. Особенно богаты коммелиновыми тропики и субтропики Америки и Африки, где сосредоточено подавляющее большинство родов.
Коммелиновые - многолетние, реже однолетние травы обычно с более или менее мясистыми, узловатыми стеблями и листьями и волокнистыми, реже сильно утолщенными и клубневидными корнями. Встречаются среди них лианы и эпифиты. Почти лишены стебля представители африканского тропического рода антерикопсис (Anthericopsis). Очередные, цельные, плоские или желобчатые с параллельным жилкованием листья коммелиновых расширены у основания в пленчатые, почти всегда замкнутые в трубку влагалища. У некоторых родов, как баллия (Ballya), буфоррестия (Buforrestia) и другие, соцветия прорывают листовое влагалище и через отверстие в нем выносят цветки наружу. Листья большинства коммелиновых опушены маленькими, 3-клеточными, железистыми волосками, различимыми обычно только под микроскопом. Густое железистое опушение наблюдается только у представителей рода флоскопа (Floscopa) и монотипного родезийского рода трицерателла (Triceratella). Устьица, как правило, с 4-6 побочными клетками и лишь у трицерателлы и американского рода фиодина (Phyodina) они с 2 побочными клетками. Членики сосудов обычно имеются во всех вегетативных органах. В стеблях и листьях они с простой и частично с лестничной перфорацией, а в корнях только с простой перфорацией и лишь у картонемы (Cartonema, рис. 179, 2), у которой сосуды только в корнях, они бывают изредка и с лестничной перфорацией. Характерным для коммелиновых является также содержание в тканях оксалата кальция, а у некоторых видов и кремнезема. Нет оксалата кальция только у австралийского рода картонема. Цветки коммелиновых небольшие, обычно актиноморфные или более или менее зигоморфные, как правило, обоеполые, собраны в верхушечные или расположенные против листьев пазушные, в основном верхоцветные соцветия, состоящие из более или менее редуцированных одиночных или двойных завитков. Редко цветки одиночные. Встречаются также и полигамные растения, несущие обоеполые и однополые цветки на одном и том же растении или на разных особях одного и того же вида. Примером может служить индокитайский род спатолирион (Spatholirion). Околоцветник состоит из 3 обычно зеленых, но иногда лепестковидных и окрашенных (как у неотропического рода дихоризандра - Dichorisandra) и, как правило, свободных чашелистиков и такого же числа лепестков. Редко чашелистики срастаются в узкую трубку (например, у колеотрипы - Соleotrype, цианотиса - Cyanotis, рис. 179, 7, 8 и зебрины - Zebrina). Лепестки ярко окрашенные, голубые, розовые, белые, реже желтые, свободные или слегка сросшиеся, иногда ноготковые (как у коммелины - Commelina, рис. 179, 4), редко один из них более или менее сильно редуцирован. Замечательной особенностью лепестков многих коммелиновых является их способность при отцветании превращаться в студнеобразную массу, что очень затрудняет работу с гербарием, где цветки ком мелиновых редко полностью представлены. Из 6 тычинок расположенных в 2 кругах, нередко фертильны только 3, а остальные превращены в стаминодии или недоразвиты. Андроцей отличается удивительным разнообразием и иногда, как, например, у мурданнии (Murdannia, рис. 180, 4), варьирует даже в пределах рода. Все 6 тычинок полностью развиты, например, у палеотропического рода цианотис. Редко развита лишь одна тычинка. Нити тычинок свободные или крайне редко частично сросшиеся. У некоторых растений они густо опушены длинными ярко окрашенными волосками, делающими такие цветки заметными и привлекательными для насекомых. Интересной особенностью обладают тычинки коммелиновых: цитоплазма в клетках их волосков циркулирует настолько быстро, что ее движение можно наблюдать под микроскопом. Некоторые виды, как, например, традесканция вирджинская (Tradescantia virginiana, табл. 44, 1), служат для наблюдения и демонстрации этого явления для учебных целей. Самый яркий эффект достигается при исследовании только что распустившихся цветков. Пыльники при созревании раскрываются продольными щелями или реже порами, как у видов американского рода дихоризандра (рис. 179, 6). Необычной формы,, улиткообразно закрученные пыльники характерны для другого американского рода - кохлиостемы (Cochliostema, рис. 180, 2, 3). Гинецей синкарпный, со столбиком, заканчивающимся головчатым или 3-лопастным рыльцем; завязь верхняя, сидячая или слегка приподнятая на короткой ножке, обычно 3-, реже 2-гнездная. В каждом гнезде находится от одного до нескольких ортотропных, гемитропных или анатропных семязачатков. Плод обычно тонкостенная локулицидная коробочка, редко нераскрывающийся (иногда сочный). Нередко плод окружен сильно разросшимися как у буфоррестии, или сочными, как у кампелии (Campelia), чашелистиками. Интересной формы, похожая на стручок крестоцветных удлиненно-линейная коробочка у индокитайского рода этиолирион (ASthiolirion, рис. 179, 10). Семена с обильным мучнистым эндоспермом и маленьким зародышем. Сверху он прикрыт очень маленькой дисковидной или конической крышечкой, так называемым эмбриотегием, отделяющимся при прорастании семени. У большинства коммелиновых он дорсального или латерального типа и только у цианотиса и белосинапсиса (Belosynapsis) горизонтальный, верхушечный (рис. 179, 8). Дж. Бренан (1966) полагает, что это, вероятно, не более чем необычное проявление дорсального эмбриотегия. Семена обычно сетчатые, колючие и ребристые, редко крылатые. У дихосандры черные семена окружены кораллово-красным ариллусом, который, по мнению О. Рохведера (1970), не настоящий ариллус, а лишь сильно разросшаяся часть семенной кожуры, верхних слоев ее наружного интегумента.
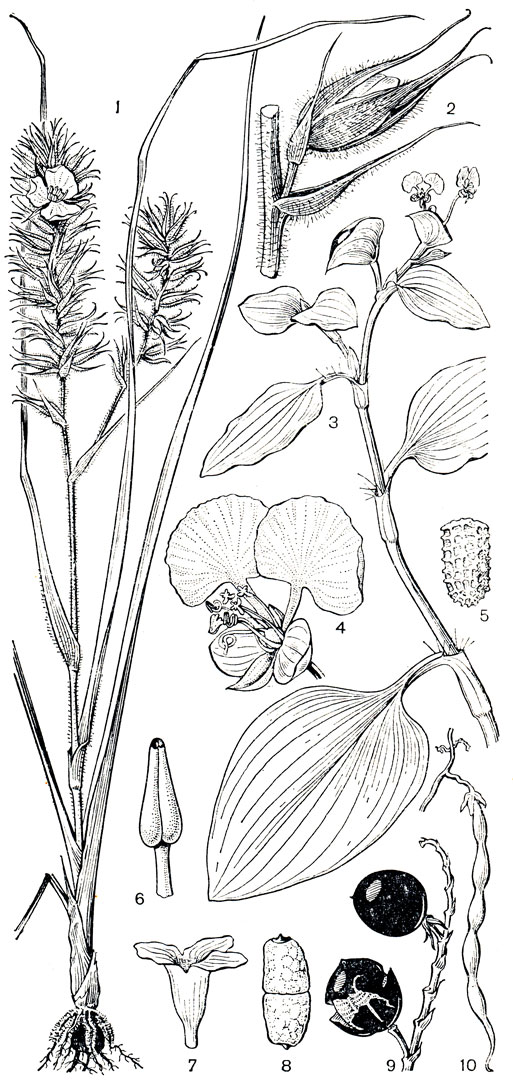
Рис. 179. Коммелиновые. Картонема колосистая (Cartonema spicatum): 1 - общий вид; 2 - цветок. Коммелина обыкновенная (Commelina communis): 3 - верхняя часть растения с двумя цветками; 4 - цветок. Коммелина Седлова (С. sellowiana): 5 - семя с боковым эмбриотегием. Дихоризандра шеститычинковая (Dichorisandra hexandra): 6 - тычинка. Цианотис бородчатый (Cyanotis barbata): 7 - спайнолепестный венчик: 8 - семя с горизонтальным верхушечным эмбриотегием. Поллиясарсогонская (Pollia sorzogonensis): 9 - плоды. Этиолирион узколопастной (Aethiolirion stenolobium): 10 - плод
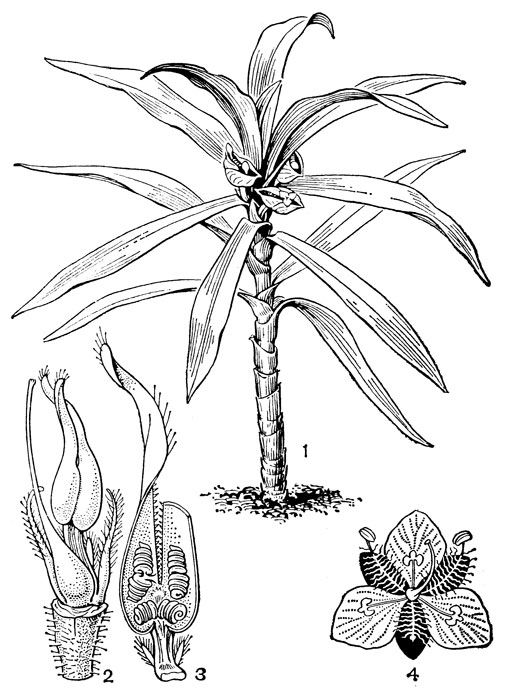
Рис. 180. Коммелиновые. Рео разноцветное (Rhoeo discolor): 1 - общий вид. Кохлиостема ароматнейшая (Cochliostema odoratissimum): 2 - андроцей и гинецей, лепестки удалены; 3 - фертильные тычинки с улиткообразнозакрученными пыльниками; нити двух задних, срастаясь, образуют капюшонообразный вырост с двумя длинными направленными вверх концами, один из которых удален. Сзади виден стаминодий. Мурданния простая (Murdannia simplex): 4 - цветок

Таблица 44. Коммелиновые: 1 - традесканция вирджинская (Tradescantia virginiana), оранжереи Ботанического института АН СССР в Ленинграде; 2 - пирреима Лоддигеза (Pyrrheima loddigesii), оранжереи Ленинградского государственного университета
Большинство коммелиновых опыляются насекомыми. Их небольшие, но ярко окрашенные цветки в густых соцветиях хорошо заметны на расстоянии. Цветки лишены нектарников и привлекают насекомых яркими лепестками и ярко окрашенными стаминодиями, в то время как фертильные тычинки часто бывают неприметными. Являются ли стаминодии источником питания для насекомых или служат лишь для их привлечения, пока не выяснено. Основные опылители - пчелы и более редко шмели, которые садятся на пыльники или лепестки и г собирая пыльцу, невольно задевают рыльце, торчащее в центре цветка. Американский ботаник У. Стивене (1948) высказал предположение о возможности самоопыления в том случае, когда перекрестное опыление почему-либо не произошло. Он считает, что быстро отцветающий и превращающийся в желеобразную массу венчик сжимает в один комок внутренние части цветка, заставляя их соприкасаться друг с другом. Однако, как показали многочисленные исследования, самоопыление у этих растений не происходит и семена не завязываются. Отцветающий венчик сжимает в комок только тычинки, а торчащее на длинном столбике рыльце оказывается снаружи. Кроме пчел и шмелей, распустившиеся цветки традесканций посещают многочисленные мухи, клопы, зеленые и черные жуки, хотя их роль в опылении пока не доказана. Замечено также, что некоторые насекомые, в частности различные жуки, нередко поедают тычинки и другие части цветка, особенно активизируясь с наступлением сухого сезона. Перекрестному опылению нередко способствует дихогамия. Примером протогиничных цветков могут служить цветки коммелины африканской (С. africana). Раскрываясь, они выставляют наружу только столбик; нити тычинок в это время скручены и пыльники расположены в основании цветка. Позднее, когда тычинки выпрямляются, вынося пыльники наверх, столбик сгибается, убирая рыльце с их пути. Ярко-желтые у коммелины африканской и голубые у коммелины бенгальской (С. benghalensis) цветки этих растений, как и у других коммелиновых, недолговечны: раскрываясь утром, они увядают уже через несколько часов. Протандричны цветки большинства традесканций.
Любопытно происходит опыление у мексиканского растения тинантии Прингла (Tinantia pringlei), прекрасно приспособившегося к горным известняковым осыпям. Наряду с обычными для большинства коммелиновых соцветиями, расположенными во влагалищах верхушечных листьев, оно образует цветочные побеги, как правило, с единственным маленьким цветком в нижней части растения. Открывающиеся первыми цветки верхних побегов обоеполые и самоопыляющиеся, а более поздние несут абортивные завязи и, следовательно, не образуют семян. Цветочные побеги второго типа, выходя наружу, пронзают листовые влагалища, самых нижних листьев, расположенных иногда почти полностью под землей или возвышающихся над ней не более чем на 3- 5 см. Эти цветки не открываются совсем (клейстогамия) или приоткрываются только частично, и плоды развиваются под землей или на ее поверхности. Лепестки подземных цветков, как правило, сильно редуцированы. Геокарпными среди коммелиновых являются также коммелина бенгальская и коммелина Форскола (С. forskalaei).
Семена коммелиновых распространяются главным образом различными травоядными животными, поедающими их сочные побеги. Попавшие в желудок вместе с зеленой массой, семена не претерпевают изменений и беспрепятственно выходят наружу. Снабженные двумя воздушными мешками семена коммелины африканской, вероятно, переносятся водой или ветром, а хорошо заметные на расстоянии, окруженные кораллово-красным ариллусом семена дихоризандры распространяются птицами.
В семействе коммелиновых наблюдается значительное разнообразие жизненных форм. Наряду с обычными, столь привычными в комнатной культуре ампельными видами традесканций, зебрин и коммелин встречаются и настоящие травянистые лианы. В Старом Свете они представлены тремя индокитайскими родами: стрептолирион (Streptolirion), спатолирион и этиолирион с довольно широкими черешковыми листьями и компактными пирамидально-метельчатыми соцветиями. К лазающим или цепляющимся лианам относится и большинство видов тропического американского рода дихоризандра. Лишен суккулентных черт напоминающий по облику тростник род трицерателла, который вообще во многом отличается от остальных коммелиновых. Многие коммелиновые произрастают во влажных низинных и горных лесах, где они обычны по берегам рек и потоков, в глубоких ущельях и на склонах холмов. Некоторые из них приспособились к жизни и в более влажных условиях. Они встречаются в заболоченных лесах и на затопляемых равнинах, на аллювиальных террасах рек и глинистых берегах прудов, на заброшенных рисовых полях и в лужах. Нередки они и на болотах, и в неглубокой воде в прибрежной части водоемов. В тропических болотах Западной Африки можно встретить маленькое изящное растеньице с желтыми цветками - флоскопу желтоватую (Floscopa flavida) и флоскопу приручейную (F. rivularis) с мелкими голубыми или розовыми цветками. Растут коммелиновые и на песчаных морских побережьях. В Мексике виды гибасиса (Gibasis) приурочены к известняковым холмам и открытым каменистым горным склонам, где они растут среди зарослей колю-чих кустарников и кактусов. Часты традесканции и в сосновых и дубовых лесах. В Эфиопии под пологом светлых акациевых лесов нередко можно обнаружить среди многочисленных травянистых лиан коммелину огненно-реснитчатую (С. pyrroblepharis). Растущие во влажных тропических лесах на стволах старых крупных деревьев коммелиновые развивают длинные воздушные корни. Свисающие вниз наподобие веревок, они в нижней части покрыты многочисленными корневыми волосками, придающими им бархатистый вид. С помощью этих волосков происходит улавливание влаги из воздуха. Некоторые виды можно встретить как в сухих, так и во влажных, в затененных и на открытых солнцу местах. Так, коммелина африканская в Западной Африке растет на опушках леса, нередка она и на открытых просторах саванн, где прекрасно выживает, несмотря на ежегодные пожары. Немало коммелиновых широко расселились как сорные растения.
Английский ботаник Дж. Бренан (1966), основываясь прежде всего на типе соцветия, выделяет в семействе коммелиновые 15 триб (которые он называет, однако, "группами"). Он рассматривает завиток как основную структурную единицу соцветия. Среди одиночных завитков, по Дж. Бренану, основным примитивным типом следует считать верхоцветное соцветие на более или менее облиственном стебле. Интересно отметить, что у большинства родов Старого Света встречаются соцветия с одиночными завитками, а у представителей Нового Света - с двойными завитками. Производные этих типов соцветий показывают удивительный параллелизм в развитии соцветий. Среди растений Старого Света двойной завиток встречается лишь у африканского рода атерикопсис, что указывает скорее на дивергентное развитие, чем на наличие сходства с американской группой. Созданная Дж. Бренаном система была в основном подтверждена анатомическими исследованиями Р. Томлинсона (1969).
Триба, которую можно было бы назвать трибой мурданниевых (Murclannieae), объединяет растения с моноподиальными прямостоячими или стелющимися стеблями и более или менее вытянутыми завитками, обычно сгруппированными в пирамидально-метельчатые соцветия, и нераскрывающимися коробочками. У мурданнии завиток редуцирован до пучка, состоящего из одного или нескольких цветков. К этой трибе иногда относят и американский род геогенантус (Geogenanthus), по типу опушения края лепестков близкий тропическому американскому роду кохлиостема.
Необычный по облику род картонема Дж. Бренан выделяет в особую трибу картонемовые (Cartonemeae), некоторые ботаники выделяют этот род иногда даже в отдельное семейство (Cartonemataceae). Эти растения, своеобразные внешне, имеют ограниченный ареал (кроме тропической Австралии, они были найдены еще на одном из островов Ару - острове Транган, лежащем к юго-западу от Новой Гвинеи). Картонемы довольно высокие многолетние травы, отличающиеся от большинства коммелиновых отсутствием суккулентных черт, сильной опушенностью железистыми волосками, колосовидной кистью, плохо развитым зародышем и эмбриотегием, отсутствием в тканях оксалата кальция и рядом незначительных анатомических признаков. Дж. Бренан считает соцветие картонемы колосовидной кистью, состоящей из редуцированных до одного цветка одиночных завитков, что, впрочем, можно наблюдать и у других коммелиновых, например у мурданнии тончайшей (М. tenuissima).
Три рода: поллия (Pollia), палисота (Palisota) и аклисия (Aclicia), распространенных в Африке, Азии и Австралии и объединенных в отдельную трибу поллиевых (Pollieae), очень сходны между собой по облику. Все они близки к мурданниевым, отличаясь от них главным образом раскрывающимися плодами (рис. 179, 9). Сюда же Дж. Бренан относит и псевдопарис (Pseudoparis), выделяемый М. Пишоном (1964) в отдельную трибу. К трибе стрептолирионовых (Streptolirioneae) отнесены три индокитайских рода: стрептолирион, спатолирион и этиолирион - с лазающими стеблями и черешковыми листьями. Видам рода колеотрипы, относимой к монотипной трибе колеотриповых (Coleotryреае), свойственны пазушные, прорывающие листовые влагалища пучковидные соцветия и цветки со сросшимися в нижней части лепестками и 6 равными тычинками. К этой трибе близки представители трибы цианотовых (Cyanotideae) с 2 широко распространенными в Старом Свете родами: белосинапсис и цианотис, отличающимися от колеотриповых цельными листовыми влагалищами и необычным для коммелиновых горизонтальным верхушечным эмбриотегием.
Видам африканского тропического рода антерикопсис, относимого к трибе антерикопсисовые (Anthericopsideae), присуще почти полное отсутствие стебля и состоящее из 1 или нескольких ложных зонтиков (каждый из двойного сидячего завитка) соцветие. Необычайны внешне и представители другого африканского рода - трицерателлы (монотипная триба трицерателловые - Triceratelleae), несколько напоминающей по облику картонему. Эти похожие на тростник, лишенные суккулентных черт растения сочетают симподиальный, сильно укороченный стебель с соцветием в виде удлиненного завитка со свободнолепестными цветками и 6 одинаковыми или почти одинаковыми тычинками.
Самый большой в семействе (150-200 видов) пантропический род коммелина объединен в одну трибу коммелиновые (Commelineae) с 2 близкими к нему родами с зигоморфными цветками, собранными в 1-2 завитка и более или менее завернутыми в покрываловидный прицветник. К. Линней назвал не без определенного смысла этот род коммелина: зигоморфный венчик состоит из 3 лепестков, 2 из которых крупные, ярко-голубые, а третий много меньше и более бледный, иногда белый. И назван этот род в честь братьев Коммелин, двое из которых были известными учеными, а третий ничем особенным не прославился. Это была одна из многих остроумных шуток великого натуралиста.
Группу американских, главным образом тропических родов, выделенных Р. Вудсоном (1942) в отдельную трибу традесканциевые (Tradescantieae), Дж. Бренан оставляет почти без изменений, исключая из нее только коммелину и 2 близких ей рода. Очень разнообразные внешне, они все объединены особым устройством соцветия, состоящего из конечных или пазушных, двойных, сидячих завитков, сросшихся с цветоножкой. Сюда относится названный в честь английского ботаника Дж. Традесканта - отца род традесканция, представленный более 60 видами прямостоячих или стелющихся многолетних трав, распространенных в Северной и Южной Америке. Любопытно, что виды этого рода служили объектами космических исследований на кораблях "Восток" и "Восход".
К трибе зебриновых (Zebrineae) относятся 4 американских рода: зебрина, сепаротека (Separotheca), сеткризия (Setcreasea) и велдения (Weldenia), достаточно близкие к традесканциевым, но отличающиеся от последних сросшимися в нижней части в трубку лепестками. Неясно положение необычно выглядящего рода велдения с цветами и листьями, собранными в плотную розетку.
Представленный в тропических областях Америки несколькими десятками видов род дихоризандра отличен от остальных коммелиновых способом раскрытия пыльников и семенами с мясистым ариллусом. Еще М. Пишон (1946) выделил этот род в монотипную трибу дихоризандровые (Dichorisandreae). Несмотря на явную обособленность, этот род довольно близок к 3 американским родам: кохлиостеме, сидеразису и геогенантусу, которые Дж. Бренан помещает в разные трибы. С геогенантусом дихоризандру сближает такой чрезвычайно редкий у американских видов признак, как разорванное листовое влагалище. Кроме того, многие из них часто очень схожи внешне. О несомненной близости этих родов говорит и сходный по морфологии и их числу кариотип (2n = 38), отличимый от остальных коммелиновых. У растущей в Бразилии дихоризандры продырявленной (D. perforans), несущей как верхушечные, так и боковые соцветия, последние прорывают листовые влагалища. Подобная картина встречается лишь у африканского рода полиспата, один из видов которого - полиспата метельчатая (Polyspatha paniculata) также несет на одном и том же растении как обычные, так и разрывающие листовые влагалища соцветия.
К трибе кохлиостемовых (Cochliostemateae) отнесен южноамериканский род кохлиостема, характеризующийся оригинально устроенным андроцеем. Оба вида этого рода представляют собой суккулентные бесстебельные или с довольно длинным (до 2 м) полулазающим стеблем эпифитные растения с крупными, обычно до метра и более длиной, собранными в розетку листьями. Выставленная впервые на майской Парижской выставке 1867 г. кохлиостема ароматнейшая (С. odoratissimum, или С. jacobianum, рис. 180, 2, 3) вызвала сенсацию среди ботаников своим необычным видом. Но еще большее изумление вызвало появление в сентябре многочисленных пурпурных цветков со сложно устроенным андроцеем. Пурпурные или бледно-голубые со слабым приятным запахом цветки несут 3 фертильные тычинки. Два перистоопушенных длинными желтыми волосками стаминодия располагаются по сторонам завязи, а третий, более мелкий, иногда почти редуцированный, находится позади фертильных тычинок. Нити двух тычинок разрастаются в сторону и вверх от улиткообразно закрученных пыльников, образуя капюшоновидный вырост с двумя длинными, вверх направленными концами; в нижней, расширенной части его и находятся пыльники всех трех тычинок (рис. 180, 2, 3).
Многие коммелиновые (особенно роды рео - Bhoeo, рис. 180, 2, зебрина, цианотис, коммелина, традесканция и др.) очень декоративны и широко культивируются как в открытом грунте, так и в оранжереях многих стран. Благодаря необычайно окрашенным (фиолетовым, зеленым, пятнистым, полосатым), красивым сочным листьям, непритязательности, легкости и быстроте размножения вегетативными частями и семенами коммелиновые (например, зебрина висячая - Zebrina pendula) являются излюбленными объектами комнатного и оранжерейного цветоводства.
У нас на Кавказе, Украине и Дальнем Востоке растет единственный представитель этого теплолюбивого семейства - коммелина обыкновенная (Commelina communis, рис. 179, 3, 4), опасный сорняк, далеко распространившийся за пределы своего естественного ареала. На Дальнем Востоке коммелина обыкновенная встречается не только среди посевов, но и на старых паровых полях и залежах, где образует сплошные заросли красивого голубого цвета. В прошлом веке ее специально возделывали на Амуре для получения из лепестков пигмента для окрашивания рыбьих кож в особо любимый местными народами (эвенками и нанайцами) голубой цвет. В сентябре на Черноморском побережье Кавказа вдоль дорог и тропинок можно видеть это небольшое стелющееся растение с удлиненно-овальными листьями и ярко- голубыми цветками. За чистую окраску цветков ее еще называют и синеглазкой. За недолговечность цветков, не увядающих, как обычно, а превращающихся в желеобразную каплю, коммелины на родине называют дневной красавицей.
Менее известны лекарственные свойства коммелиновых. Их используют в народной медицине при лечении различных заболеваний. Корни некоторых коммелин, например коммелины клубневой (С. tuberosa) и коммелины голубой (С. coelestis), содержат питательные вещества и употребляются в пищу, как и корни коммелины обыкновенной.
Семейство майяковые (Mayacaceae)
Семейство майяковых, представленное одним родом майяка (Мауаса), распространено главным образом в Америке. Лишь один вид - майяка Баума (М. baumii) - растет в тропиках Западной Африки, на территории Анголы, Заира и Замбии. На Американском континенте майяковые встречаются от юго-востока Соединенных Штатов Америки (где граница проходит от Северной Каролины до Флориды и на западе до Восточного Техаса) и дальше через Центральную Америку до Боливии, Северо- Восточной Аргентины и Уругвая. Растут майяковые и на Багамских, Бермудских и Антильских островах (см. карту 9). В роде майяка 4 полиморфных вида, но некоторые авторы доводят число видов до 10 или даже 12.
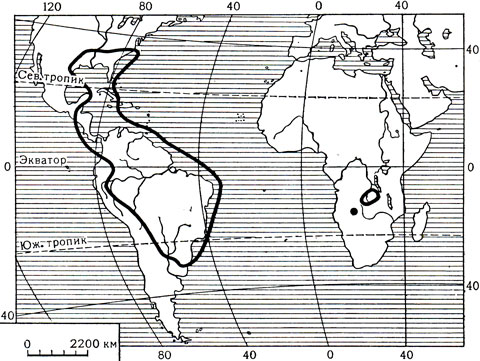
Карта 9. Ареал семейства майяковых
Майяковые пресноводные, как правило, плавающие или полностью погруженные в воду травы с простым или ветвистым слабым стеблем, покрытым многочисленными маленькими линейными или нитевидными листьями, и с очень тонкими нитевидными корнями. Внешне эти маленькие, изящные растения удивительно похожи на знакомый северному жителю мох кукушкин лен (Polytrichum, рис. 181). Лишь когда на них появляются небольшие нежноокрашенные цветки, становится ясно, что это цветковые растения.
Растут майяки и на суше по берегам прудов, озер, канав, на болотах и трясинах, где они предпочитают богатые гумусом почвы, в то время как водные формы укореняются прямо в береговом песке. Майяковые плохо переносят соседство других растений и только в отсутствие их образуют на влажных открытых местах довольно плотные дерновинки. Тщательный анализ таких дерновинок обнаружил среди майяк лишь единичные стебельки мха, печеночника, мелких элеохарисов (Eleocharis) и листья щитолистника буэнос-айресского (Hydrocotyle bonariensis).
Так же как и у других водных растений, наземные формы майяк значительно мельче, чем водные. Так, например, длина стебля майяки речной (M. fluviatilis) у экземпляров, растущих на берегу, не превышает 2-2,5 см, только в затененных местах иногда достигая 5-8 см, в то время как среди погруженных в воду растений нередки экземпляры, чей слабый, вялый стебель вырастает в длину до 1 м и более.
Приспособление к водному образу жизни нашло отражение и в своеобразном анатомическом строении всего растения. В стеблях, листьях и корнях находятся воздушные каналы с поперечными перегородками из звездчатых клеток. У майяковых отсутствуют механические ткани, функции которых выполняет окружающая редуцированную стелу эндодерма. Иногда она сильно утолщена, например, в стебле. Отсутствуют также волоски, за исключением рано опадающих, развивающихся в пазухах листьев, и секреторные элементы. У водных форм все эти признаки выражены сильнее, чем у наземных.
Листья майяковых очередные, сидячие, с одной срединной жилкой, лишены влагалищ и часто оканчиваются на верхушке двумя зубцами. У наземных форм майяки речной они очень мелкие, длиной всего 2-3 мм, а у водных длина достигает 1 -1,8 см. Интересно, что в листовых пазухах очень молодых листьев развиваются нитевидные 2-3-клеточные волоски, сходные с волосками эриокаулоновых. Развиваясь очень рано, они образуют густое опушение, служащее защитой для листовых зачатков (примордиев) и верхушек молодых побегов. Выполнив свою роль, волоски отмирают и на старых побегах, как правило, отсутствуют.
Маленькие, обоеполые и актиноморфные цветки располагаются по одному в пазухах листьев или, реже, собраны на верхушке побега в зонтиковидное соцветие (рис. 181). Прозрачные пленчатые прицветники часто отгибаются вниз после цветения. Закладываясь как один, прицветник довольно рано, часто еще до того, как цветоножка достигнет своей нормальной длины, расщепляется продольной щелью на две части. Двойной околоцветник состоит из 3 почти створчатых чашелистиков, 3 черепитчатых лепестков и чередующихся с ними тычинок. Обратнояйцевидные, чисто-белые, розовые или фиолетовые лепестки лишь ненамного превышают чашелистики. Тычинки с тонкими, иногда слегка расширенными у основания нитями и пыльниками, прикрепленными к нитям своими основаниями, составляют при-мечательную черту семейства. Особенно важен способ открывания пыльника, являющийся основным признаком при определении видов. Пыльники раскрываются верхушечными или почти верхушечными порами или поровидными щелями и высыпают многочисленную пыльцу прямо наружу или в вытянутую в виде трубки или кубка верхушечную часть пыльника. Некоторое представление о разнообразии строения андроцея майяковых дает рисунок 181. У майяки Баума поровидное отверстие, через которое высыпается пыльца, прикрыто в незрелых пыльниках мозолистым утолщением, позднее растворяющимся и освобождающим выход из пыльника (рис. 181). Для майяковых характерен паракарпный гинецей, состоящий из 3 плодолистиков. Имеются указания на случайное нахождение 4 и 5 плодолистиков. Завязь верхняя, с нитевидным столбиком, за-канчивающимся цельным или 3-лопастным рыльцем, часто сохраняющимся до раскрытия , плода. В каждом гнезде завязи по нескольку ортотропных семязачатков. Число их колеблется от 6 до 30 у разных видов. Плод - трехгранная коробочка, раскрывающаяся тремя створками. Мелкие, шаровидные или яйцевидные, темноокрашенные семена с сетчато-ямчатой поверхностью. Маленький верхушечный зародыш наверху с небольшим чашевидным эмбриотегием, образующимся, вероятно, из клеток внутреннего интегумента. При прорастании семени эмбриотегий разрушается, образуя канал, через который 4зародыш проникает наружу.
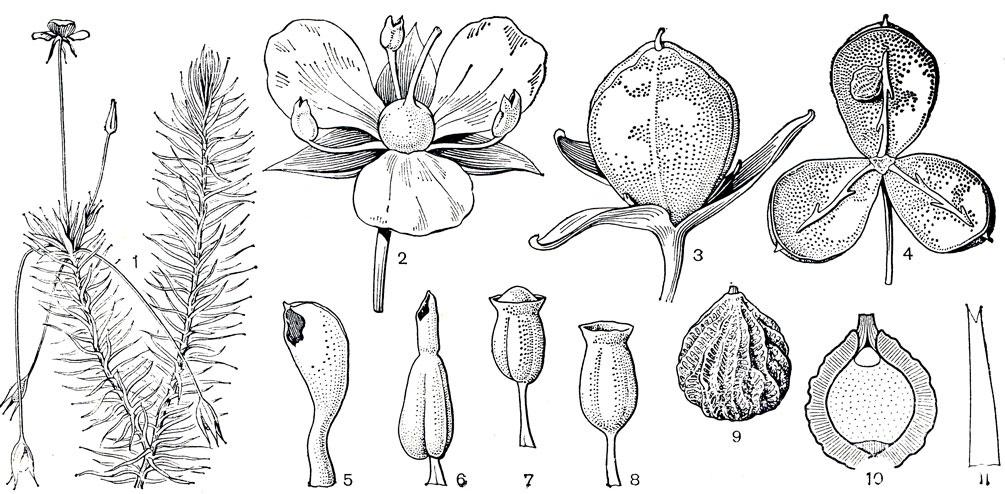
Рис. 181. Майяковые. Майяка длинноножковая (Mayaca longipes): 1 - побег с цветками. Майяка речная (М. fluviatilis): 2 - цветок. Майяка Ванделла (M. vandellii): 3 - молодой плод; 4 - раскрытый зрелый плод. Майяка речная: 5 - тычинка (вид сбоку). Майяка Селло ва (M. sellowiana): 6 - тычинка. Майяка Баума (M. baumii): 7 - молодая тычинка, видно мозолистое утолщение, закрывающее пору; 8 - зрелая тычинка. Майяка Ванделла: 9 - семя. Майяка речная: 10 - схематический разрез семени, сверху виден эмбриотегий. Майяка Ванделла: 11 - лист с двумя зубцами на верхушке
Интересен процесс опыления у майяковых. Строение цветка позволяло предполагать, что оно осуществляется насекомыми. Однако И. Упхофу (1933), в течение 7 лет наблюдавшему цветение майяки речной, не удалось обнаружить насекомых на ее маленьких, диаметром не более 4 мм, неприметных цветках. Те же случайные насекомые, которые крайне редко оказывались на этих растениях, не могли считаться их регулярными опылителями. Тем не менее на растениях постоянно образовывались плоды даже в холодную погоду, когда цветки остаются закрытыми.
Во Флориде майяка речная цветет почти весь год, за исключением лишь самого холодного времени или разгара летней жары. Цветки ее настолько мелки и неприметны, что только в момент массового цветения их можно разглядеть на фоне темно-зеленых мохообразных дерновинок. Открываются они утром, большей частью между 11 ч утра и часом пополудни. К вечеру большинство из них увядает. Лепестки становятся бурыми и засыхают, а зеленоватые чашелистики поднимаются кверху и смыкаются вокруг завязи, лишь позже расходясь и открывая молодой плод. Самоопыление осуществляется благодаря особому устройству цветка: трехлопастное рыльце находится на одном уровне и в непосредственной близости с пыльниками, и таким образом высыпающаяся пыльца попадает на рыльце. Однако, как показали наблюдения, на рыльце попадает немного пыльцы, в результате чего развивается незначительное количество семян. В редких случаях можно встретить цветки, у которых самоопыления не происходит из-за того, что тычинки их несколько короче рыльца. Достаточно разницы в 0,5-1 мм, чтобы самоопыление стало невозможным. При отцветании тычинки поникают или опадают, а побуревшее и сморщенное рыльце нередко сохраняется до раскрытия плода. Плоды развиваются быстро, и уже на следующий день становится заметным увеличение оплодотворенной завязи. Постепенно плоды приобретают треугольную форму, подобную плодам гречихи. Стоящая вертикально цветоножка все больше наклоняется к земле и все ниже опускает созревающие плоды. Через 4-6 недель плоды растрескиваются тремя отгибающимися назад створками и созревшие семена легко выпадают на землю, где и прорастают рядом с материнским растением.
Особенно любопытно опыление у подводных форм майяки речной, образующей плотные заросли на глубине 1 - 1,2 м. Полностью клейстогамные, т. е. никогда не раскрывающиеся, цветки ее располагаются на цветоножках, свободно торчащих во все стороны от подводного стебля. Они состоят из недоразвитых коричневатых или прозрачных лепестков и обычных чашелистиков, тычинок и завязи. Так же как и у наземных форм, пыльники располагаются в непосредственной близости от рыльца. Внутри плотно закрытого цветка сухо. Вода проникает в него крайне редко и в незначительных количествах, и даже в этих случаях она не препятствует опылению. Как показали исследования, попадающая в воду пыльца сохраняет способность к прорастанию. Развитие плодов идет так же, как и у наземных форм, от которых они отличаются только несколько более угловатой формой, хорошо сохранившимся столбиком и более светлоокрашенными семенами. Высыпающиеся семена распространяются водой на большие расстояния, чем у наземных форм.
Любопытно отметить, что семена майяк легче переносят высыхание, чем нахождение под водой. Как показал опыт, семена, подвергшиеся высушиванию в течение 6 недель, при намачивании прорастали очень быстро, а те, что выдерживались под водой, не дали всходов и через 12 недель.
Экономическое значение майяковых неизвестно, и в культуре они не встречаются. Вероятно, они могут быть использованы как изящные и оригинальные аквариумные растения.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'