Порядок араукариевые (Araucariales)
Семейство араукариевые (Akaucakiaceae) (О. А. Муравьева, В. С. Борхвардт, С. Г. Жилин)
Араукариевые, с которых начинается описание семейств хвойных, часто называют "южными хвойными". Это связано с тем, что большинство видов араукариевых - обитатели южного полушария. Облик этих вечнозеленых растений весьма необычен для жителя северной умеренной зоны. Крупные, широкие, со многими параллельными жилками, листья агатиса (Agathis), одного из двух родов араукариевых, скорее вызывают представление об однодольных, чем о хвойных. Однако и неискушенный в ботанике наблюдатель быстро поймет, что он имеет дело с хвойными, узнав о наличии смолы и шишек у этих растений. Не только агатис, но и некоторые виды араукарий (Araucaria) с их довольно длинными плоскими листьями не ассоциируются с привычными для северянина представлениями о хвойных растениях, которые созданы с детства близким знакомством с елями, пихтами, соснами. Особенно поражает северян араукария чилийская, или араукская (Araucaria araucana), обладающая столь жесткими и колючими листьями, что, говорят, птицы не садятся на ее ветви. С этой особенностью араукарии чилийской связано одно из ее обиходных названий на английском языке - Monkey Puzzle, т. е. "загадка обезьяны", широко употребляемое и вошедшее в ботанические словари. Оно возникло, впрочем, не в Чили, а в Великобритании. В Корнуолле (на юго-западе Великобритании) некий гордый обладатель молодого экземпляра араукарии чилийской (она долгое время покрыта целиком - и ствол и ветви - колючими листьями), показывая его гостям, заметил: "Забраться на это дерево было бы загадкой и для обезьяны!"
Ботаников, путешествовавших по араукариевым лесам, не оставляло ощущение, что они чудом попали в "ископаемый", вымерший лес. И такое ощущение в общем-то не является обманчивым. Араукарии действительно очень древние растения. Недаром их часто называют "живыми ископаемыми". Настоящие же ископаемые остатки араукарий зарегистрированы в земных слоях, возраст которых исчисляется весьма внушительными цифрами - в 150 и даже 240 млн. лет!
Что же создает впечатление древности при виде араукарий? Если иметь в виду молодые растения, то араукария чилийская со стволом, сплошь покрытым колючими листьями, конечно же, напоминает реконструкции вымерших древесных или древовидных растений с облиственными стволами, например лебахии (Lebachia) - одного из древнейших палеозойских хвойных (рис. 185). Кажутся архаичными и взрослые экземпляры араукарий в бразильских и чилийских араукариевых лесах благодаря их высоко поднятым зонтиковидным кронам.
Древесина араукариевых имеет весьма своеобразное строение. Некоторыми чертами она напоминает кордаитовые и древнейшие палеозойские хвойные - вальхиевые (Walchiaceae) и вольциевые (Voltziaceae). У араукариевых, как и у кордаитовых, сердцевина выражена отчетливо. Ствол толщиной 20 см имеет сердцевину диаметром около 2 см. В клетках сердцевины содержатся дубильные вещества - танины, смолы, слизи. Кольца прироста у боль-шинства видов плохо различимы или едва заметны и лишь у отдельных видов отчетливо выражены. Вторичная древесина состоит из очень длинных трахеид, иногда до 10-12 мм.
У араукарии расположение пор на трахеидах одно-двурядное, реже, в зрелой древесине, трехрядное. У агатиса поровость двурядная или трехрядная, реже поры располагаются в один ряд, а иногда в четыре и даже в пять-шесть рядов. Окаймленные поры на стенках трахеид сильно скучены, тесно соприкасаются друг с другом и тогда приобретают шестиугольное очертание. Такой тип многорядной поровости с очередным расположением пор на стенках трахеид получил название араукариоидного типа и является важнейшим диагностическим признаком, отличающим древесину араукариевых от древесины всех других современных хвойных. Правда, как редкое исключение, подобная же поровость, и лишь на концах немногих трахеид, наблюдается у некоторых видов дакридиума (Dacrydium) из подокарповых.
Торус окаймленных пор, хорошо выраженный у других хвойных, в древесине араукариевых отсутствует или, очень редко, имеется в виде небольшого утолщения средней части пленки поры. Древесинная паренхима очень скудная или совершенно отсутствует. Лучи обычно однорядные, редко двурядные; гомогенные (состоящие только из паренхимных клеток, в которых образуется смола, выделяющаяся в смежные с ними трахеиды). В коре стебля, иногда в большом количестве, встречаются смоляные каналы, наполненные смолой.
Некоторые виды рода агатис имеют очень крупные, широкие
листья. Например, у агатиса крупнолистного (Agathis macrophylla) и агатиса Брауна (А. brownii) листья достигают длины 15-18 см и ширины 6 см. Такие листья с многочисленными, параллельно идущими жилками, которые у основания пластинки листа дихотомически ветвятся (рис. 193), внешней формой и величиной напоминают листья некоторых палеозойских кордаитов (Cordaites) и немногих видов современных подокарпов (Podocarpus). До-вольно крупные и широкие плоские листья и у некоторых видов араукарий. Например, листья араукарии Ханстайна (Araucaria hunsteinii) достигают в длину 10 см. Другие араукарии несут мелкие чешуевидные листья. Так, у араукарии Баланса (A. balansae) листья длиной 0,3-0,5 см и шириной 0,2 см. У молодых экземпляров араукарии разнолистной (A. heterophylla) молодые (ювенильные) игловидные листья существуют до 30-40-летнего возраста. После этого растение переходит к репродуктивной фазе и ювенильные листья сменяются взрослыми (дефинитивными), чешуевидными. Растение таким образом совершенно меняет свой облик, становится неузнаваемым (рис. 188).

Рис. 188. Араукария разнолистная (Araucaria heterophylla): 1 - ветвь с молодыми листьями (уменыи.); 2 - ветвь со взрослыми листьями (увел.)
Широкие листья агатисов и некоторых видов араукарий пронизаны многочисленными проводящими пучками (и, соответственно этому, обладают многочисленными жилками). Проводящие пучки в листьях чередуются с проходящими рядом с ними смоляными каналами, что является надежным диагностическим признаком, отличающим араукариевые с широкими крупными листьями от видов подокарпа. Впрочем, среди видов араукарий есть исключение - араукария Руле (A. rulei), у которой, подобно подокарповьгм, смоляные каналы находятся под пучком.
К особенностям араукарий и агатисов относится так называемый веткопад: свойство целиком сбрасывать облиственные боковые побеги или даже ветви с листьями. Веткопад у хвойных не ограничивается араукариевыми. Это свойство имеют также таксодиевые: так содиум (Taxodium), метасеквойя (Metasequoia) и особенно куннингамия (Cunninghamia). Особенностью араукариевых является развитие у них ветвящихся пыльцевых трубок. Пыльцевые зерна араукариевых часто прорастают не на семязачатке, а в пазухе шишечной чешуи. Главные ветви пыльцевых трубок направляются при этом прямо к нуцеллусу, а другие либо растут в сторону микропиле соседних семязачатков, либо врастают в центральный цилиндр оси шишки. Они проникают вдоль проводящих пучков даже в ксилему. Американский ботаник А. Имс, тщательно изучавший этот процесс, писал, что, если такую пыльцевую трубку целиком извлечь со всеми разветвлениями из тканей, в которые она проникла, перед нами будет подобие тонкой сети паутины.
Среди ботаников нет единодушия в оценке этого явления: одни считают его признаком примитивности араукариевых, другие - наоборот, подвинутости. Но, при любом истолковании, ветвистая пыльцевая трубка араукариевых выполняет не только роль органа, транспортирующего гаметы (как у остальных хвойных и у цветковых), но и функцию всасывания питательных веществ (действует подобно гаустории гинкговых и саговниковых). Однако ветвление пыльцевых трубок, по-видимому, наблюдается не всегда. Так, у экземпляров араукарии чилийской, производившей зрелые семена, французский ботаник М. Фавр-Дюшартр (1960) обнаружил неветвящиеся пыльцевые трубки.
Род араукария получил название от провинции Арауко в южной части Чили. Так же называется и город в этой провинции, и залив, омывающий ее с запада. Впрочем, названий с основой "араук" в Южной Америке очень много (например, река Арауко в Венесуэле, интенденсия Араука в Колумбии), что скорее всего объясняется былым более широким распространением группы индейских племен арауканов, населяющих ныне провинции Арауко, Мальеко и более южные провинции Чили, а также пограничные с ними местности Западной Аргентины. И араукария чилийская (A. araucana) не случайно получила свое не только родовое, но и видовое латинское название: ее ареал в значительной степени совпадает с современной областью расселения арауканов.
Именно араукария чилийская была первым описанным видом араукарий. В 1782 г. под названием "сосны" араукской (Pinus araucana) ее описал итальянский ботаник X. Молина. О том, что "сосна" араукская является особым родом, в то время еще не подозревали. Лишь спустя четыре года, в 1786 г., знаменитый ботаник и зоолог Ж. Ламарк описал "сосну" араукскую как особый род, назвав ее в честь путешественника и собирателя растений Дж. Домби - домбеей (Dombeya). Однако несколькими месяцами раньше такое же имя и в честь того же Домби получил род цветковых растений. Поэтому французский ботаник Антуан Жюссье в 1789 г. переименовал хвойное, названное было домбеей, и с тех пор оно зовется араукарией.
Араукарии, как и многие другие хвойные, - растения долгоживущие. Возраст в несколько сот лет для них не диковинка. Считается, что зрелого состояния (в хозяйственном отношении) они достигают не ранее, чем к 300 годам, и именно с этого возраста араукариевые леса пригодны для вырубок. Максимальный возраст деревьев - 2000 лет.
Обычно араукарии - очень крупные деревья (высотой до 60-75 м), преимущественно двудомные. Интересно отметить, что женские экземпляры некоторых видов (например, араукарии чилийской) намного крупнее мужских и достигают высоты 30-60 м с диаметром ствола у основания 2,5 м и более, в то время как мужские не превышают в высоту 15-20 м. У молодых деревьев ветви располагаются вдоль всего ствола, от самого его основания. Они обычно мутовчатые, горизонтально простертые. С возрастом араукарии, как правило, сбрасывают ветви в нижней части ствола, а иногда почти до самой вершины, и стволы оказываются поэтому увенчанными уплощенными зонтиковидными кронами (араукария чилийская и араукария бразильская - A. angustifolia). Крона многих других видов араукарий отличается от описанной. Особенно примечательна она у араукарии колонновидной (A. columnaris), о чем пойдет речь чуть позже (рис. 189).
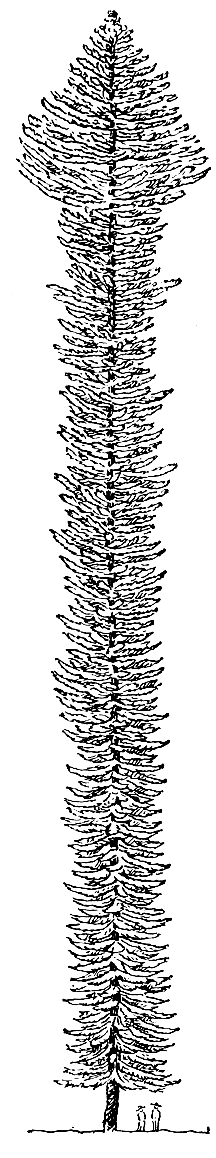
Рис. 189. Араукария колонновидная (Araucaria columnaris)
Поверхность ветвей и стволов молодых деревьев покрыта, как панцирем, выступающими основаниями отпавших листьев. У зрелых деревьев кора толстая, смолистая, бороздчатая, изредка с отшелушивающимися тонкими бумаговидными пластинками. Поверхность стволов зрелых деревьев - с поперечными кольцевыми утолщениями. Этот признак, отличающий араукарии от агатисов, обязан мутовчатому расположению ветвей, с возрастом отмирающих. Как правило, лишенные почечных чешуй почки незаметны, поскольку точка роста защищена мелкими, тесно сближенными листьями, остающимися у основания годичных приростов. Спирально расположенные листья густо покрывают ветви. На молодых боковых ветвях листья иногда двурядные, почти супротивные (араукария Бидвилла - A. bidwillii), кожистые, крупные, длиной 4-10 см, ланцетовидные или даже широкояйцевидные, со многими почти параллельными жилками.
У гетерофильных видов (араукарии разнолистная и колонновидная) листья молодых экземпляров игловидные,, с одной, редко с 3 - 5 (до 13) жилками, достигают в длину см; на вегетативных ветвях взрослых деревьев и на их репродуктивных побегах листья мелкие, длиной до 3-5 мм, чешуевидные, черепитчато налегающие друг на друга (рис. 188). Устьица у плоских листьев араукарий расположены рядами либо на обеих сторонах листа, либо только на нижней. У игловидных четырехгранных листьев молодых растений араукарии разнолистной (A. heterophylla) устьица имеются на всех гранях. Замыкающие клетки погруженные (табл. 49). Надустьичная ямка обычно прикрыта своеобразной пробочкой из воскоподобного вещества (табл. 48).
Микростробилы большинства араукарий очень крупные, самые крупные из всех хвойных, цилиндрической формы, длиной до 20- 25 см и диаметром до 4-5 см. Расположены они либо по одному на верхушках побегов, либо в пазухах побегов; в последнем случае они могут быть одиночными или сближены группами по 2-4-6 на одном побеге (рис. 191).
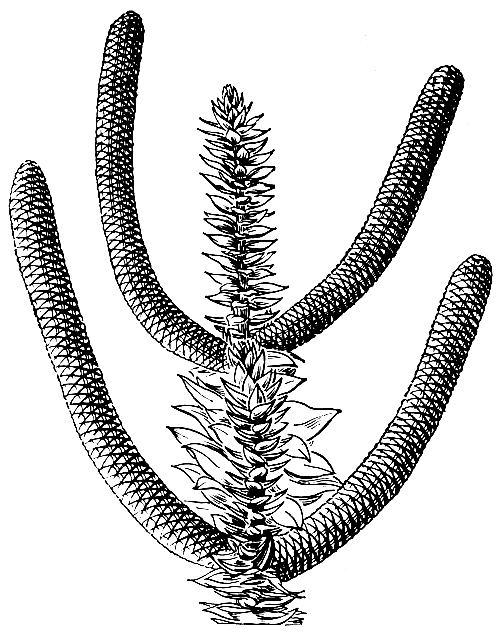
Рис. 191. Араукария Бидвилла (Araucaria bidwillii), ветвь с супротивными мужскими стробилами (схема)
Микроспорофиллы полущитковидные в виде ножки, расширенной, наверху в так называемый надсвязник, имеющий иногда длинный шиловидный придаток. К нижней поверхности надсвязника прикреплено от 3 до 20 свободных микроспорангиев, свисающих параллельно ножке спорофилла. Раскрываются спорангии продольной щелью. Пыльцевые зерна без воздушных мешков.
Шишки шаровидные или эллипсоидальные (рис. 190), диаметром 7-20, а иногда и до 35 см (у араукарии Бидвилла). Семенная чешуя срастается с кроющей на всем протяжении (кроме араукарии Бидвилла, у которой верхний кончик семенной чешуи, называемый лигулой, свободный и чуть приподнят). Такое образование, свойственное также некоторым другим хвойным, называют шишечной чешуей. Кроющие чешуи значительно крупнее семенных, с длинным, позднее обламывающимся остроконечием.
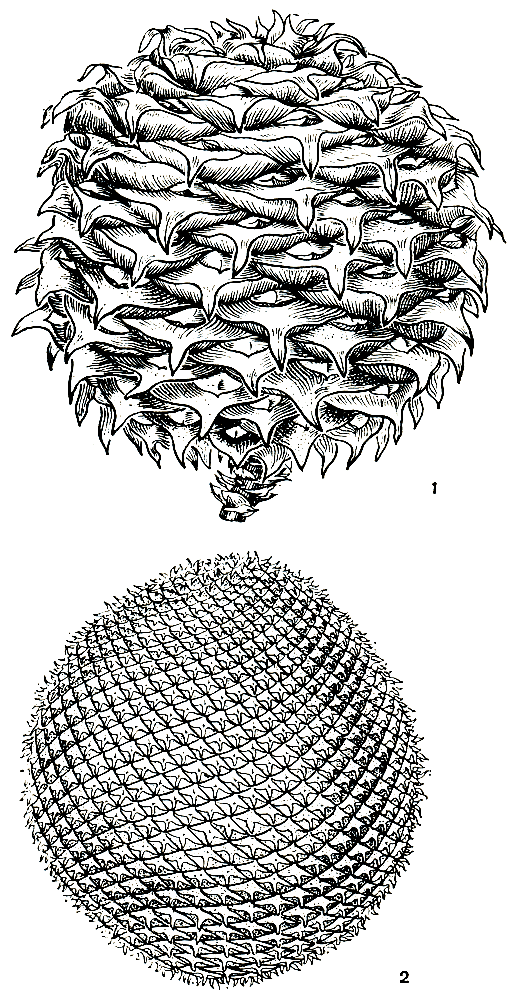
Рис. 190. Шишки араукарий: 1 - араукария Бидвилла (Araucaria bidwillii); 2 - араукария бразильская (A. angustifolia)
Семя полностью погружено в ткань шишечной чешуи и по созревании опадает вместе с нею. Но так происходит у всех видов, кроме араукарии Бидвилла, у которой вследствие разрушения наружного слоя семенной чешуи зрелое семя освобождается, как бы выскальзывая из своего вместилища. Отметим, что зрелые семена появляются у араукарий впервые в 40-50-летнем возрасте.
Семена у разных видов рода бывают двух типов. У араукарий чилийской, бразильской и Бидвилла они ореховидные, крупные (длиной до 5 см), продолговатые или яйцевидно-эллипсоидальные. У других видов араукарий семена мелкие (длиной до 1,5 см), с двумя боковыми тонкокожистыми или пленчатыми крыльями. Для ореховидных семян характерно подземное прорастание, зародыш у них с двумя, очень редко с тремя-четырьмя семядолями и сильно утолщенным гипокотилем. У мелких семян - надземное прорастание, токий гипокотиль и четыре (редко две) зеленые семядоли.
Араукария имеет разорванный ареал (карта 13), что само по себе является указанием на ее древность: ведь видам одного рода, имевшим общее происхождение из одного какого-то места, требовалось некоторое время на расселение. Но в случае с араукарией мы сталкиваемся с обитанием видов одного рода на разных континентах: в Южной Америке и в Австралии (кратчайшее расстояние - 12 тыс. км - через Тихий океан). Несомненно, что два американских вида араукарии отделились от остальных не благодаря дальнему заносу их семян, а в результате расхождения прежде соединенных континентов. Так как в третичное время Южная Америка уже была отделенной от Австралии частью суши, ясно, что разделение ареалов произошло раньше этого времени, вероятно в мезозое.
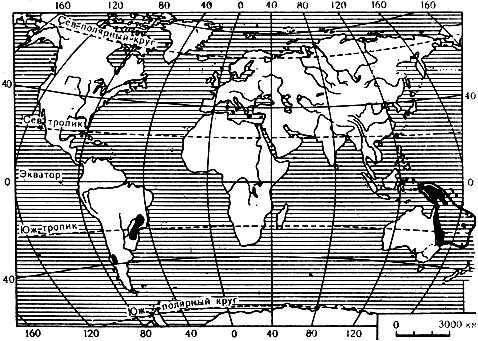
Карта 13. Ареал рода араукария
К тому же араукарии бразильская и чилийская составляют особую, морфологически ясно очерченную секцию араукария (Araucaria) рода араукария, что поддерживает мысль о давнем расчленении ареала рода.
Секция араукария характеризуется крупными плоскими листьями, крупными бескрылыми семенами, которые в зрелом состоянии опадают, так и не освободившись от сросшихся с ними шишечных чешуй.
Чрезвычайно интересны многочисленные находки окаменелых шишек в Патагонии, которые описываются палеоботаниками как араукария удивительная (A. mirabilis). Она относится к секции буния (Bunya), ныне монотипной, т. е. представленной одним видом - араукарией Бидвилла (местное название "буния-буния"), растущей на востоке Австралии, в Квинсленде. Патагония же, где обнаружена вымершая араукария удивительная, расположена на юге Аргентины, как раз между ареалами араукарий бразильской и чилийской, относящихся, как мы уже знаем, к секции араукария. Стало быть, некогда эти две секции (араукария и буния) соседствовали в Южной Америке, потом представители секции буния в Америке вымерли, а сама Америка отделилась от Австралии, По крайней мере так могло быть. К сожалению, возраст ископаемых шишек не установлен с точностью. Наиболее вероятен юрский возраст араукарии удивительной. Следовательно, разрыв ареала рода араукария мог произойти лишь в послеюрское время.
Если мы обратимся к морфологии видов, составляющих секции араукария и буния (последняя характеризуется крупными листьями и крупными семенами, которые, в отличие от семян секции араукария, опадают, освобождаясь от шишечной чешуи), то узнаем, что эти секции наиболее близки друг к другу.
Довольно отчетливо отделена от остальных секция эутакта (Eutacta), которая объединяет 15 видов, обитающих в Австралии, на острове Норфолк, на Новой Гвинее, Новых Гебридах и Новой Каледонии. Причем 12 видов этой секции являются эндемиками Новой Каледонии. Для секции эутакта характерны мелкие игловидные или чешуевидные листья и мелкие крылатые семена.
Наконец, еще одна секция, интермедия (Intermedia) получила свое название именно потому, что слагающие ее виды промежуточны (лат. intermedins означает "находящийся посреди") по своим признакам между секциями араукария и буния, с одной стороны, и секцией эутакта - с другой. У видов секции интермедия семена мелкие (как в секции эутакта), а листья крупные (как в секциях араукария и буния).
Конечно, особенности секций не ограничиваются только внешним строением семян и листьев. Их различают также тип прорастания семян, терминальное или пазушное положение микростробилов на побеге, длина ножек шишек, морфология сеянцев.
Загадочным фактом является то, что остатки побегов, семян, пыльцы араукарий обнаружены не только в южном полушарии (за пределы которого ныне араукарии не выходят), но и в северном (в Америке, Европе и на севере Африки). Может даже оказаться, что центр происхождения араукарий был в северном полушарии. Возраст находок в обоих полушариях весьма солиден - указывают на юру (200-135 млн. лет назад) и триас (240-200 млн. лет). Более древние, палеозойские находки древесины, описываемые, впрочем, под заманчивым названием Araucarioxylon (в переводе это родовое название означает "древесина араукарии"!), по-видимому, не всегда имеют отношение к семейству араукариевых, и скорее всего это не хвойные, а какие-то родственные им голосеменные растения.
Широкое хозяйственное применение араукарий (главным образом древесины, но также и смолы) привело к тому, что, помимо научных названий этих растений, на практике применяют обиходные, происхождение которых весьма различно. Одним из элементов названий араукарий на разных языках, состоящих, как правило, из двух-трех слов, очень часто выступает название одного из хвойных, обычных для северного полушария,- сосны, пихты или ели. Английские, французские и испанские обиходные названия араукарий обычно сопровождаются элементом "сосна" (pine, pin и pino соответственно). Например, "сосна чилийская" (Chile Pine) - араукария чилийская. Другой элемент может быть дан по цвету или назначению древесины: "белая сосна" (White Pine) - араукария Каннингема. Эту араукарию называют также "обручной сосной" (применяют ее в бондарном производстве) и "сосной бухты Моретон" (по месту первой находки - в Восточной Австралии). Есть случаи происхождения обиходного названия от научного: "сосна Клинка" (Klinki Pine) - араукария Клинка. Несомненно, что от обиходного названия араукарии произошло имя одного из островов близ Новой Каледонии - острова Соснового (lie des pins, по-французски), где наряду с араукарией растет также один из видов агатиса. Настоящих сосен в естественном произрастании на этом острове, разумеется, нет.
Рассмотрим наиболее примечательных представителей четырех секций рода араукария.
Араукария Бидвилла (секция буния) - единственный представитель араукариевых с совершенно самостоятельными проводящими пучками кроющей и семенной чешуй. Семенная чешуя при этом имеет даже два проводящих пучка, что характерно для пазушных побегов. У этой араукарии были обнаружены кроющие чешуи не с одной, а с двумя и тремя семенными чешуями, каждая со своим семязачатком. На такой кроющей чешуе образуются соответственно 2-3 семени. Все это свидетельствует о древности, примитивности араукарии Бидвилла. Это дало повод Р. Флорину высказать идею о происхождении араукариевых от предполагаемых палеозойских предков современных хвойных из семейства вольциевых, через промежуточный род схизолепис (Schizolepis), у которого кроющая чешуя несет трехлопастную фертильную чешую - каждая лопасть с одним семязачатком. В ходе эволюции средняя лопасть оказалась редуцированной, а две боковые слились. Следовательно, единственное семя у араукарий является остатком от первоначальных трех.
Араукария Бидвилла - растение обычно двудомное. Женские экземпляры достигают 40-50-метровой высоты, со стволом диаметром до 125 см. Молодые деревья с широкопирамидальной кроной, боковые ветви по 8-12 в мутовках, мало ветвящиеся, нередко на концах повислые. У взрослых деревьев ствол почти до половины высоты освобожден от ветвей. Кора толстая, смолистая, темная.
Почки на концах молодых побегов окружены значительно более мелкими листьями, которые впоследствии остаются на побегах; благодаря этому хорошо заметны границы одногодичных приростов побега. Листья в верхней части кроны расположены спирально, яйцевидные, с остроконечием. На боковых же побегах оно двурядно расположены в одной плоскости. Листья у этой араукарии крупные (длиной от 4 до 7,5 см и шириной до 1 см). Верхняя поверхность листьев блестящая, со многими параллельными жилками, но без киля, устьичные линии имеются на всей нижней поверхности листа. К обоим концам годичного побега размер листьев уменьшается.
Микростробилы пазушные, иногда супротивные (рис. 191), цилиндрические, очень длинные (14-20 см, при ширине 1,2-1,5 см). Надсвязник микроспорофиллов яйцевидцый с островатой верхушкой, микроспорангиев 6 и более.
Шишки располагаются на концах коротких боковых облиственных веток, вегетативные листья которых постепенно переходят в чешуи шишки. Зрелые шишки широкоэллиптические или шаровидно-яйцевидные, с толстой осью и очень крупные, в природных условиях размеры шишек достигают в диаметре 35 см и массы до 3 кг. Семена длиной 5,5 см при ширине 3 см. Среди араукарий самые крупные шишки и семена именно у араукарии Бидвилла. :
В 1843 г. английский натуралист и путешественник Дж. Бидвилл, в честь которого и назван этот вид, передал несколько экземпляров араукарии в Ботанический сад Кью (Англия). С этого началась интродукция араукарии Бидвилла в Западную Европу и Россию. В Батуми одно дерево в возрасте 80 лет имело ствол высотой 25 м и примерно в диаметре 60 см. В суровую зиму 1949/50 г., когда температура понизилась до - 8,5 °С, оно сильно пострадало, отмерли ветви, но затем дерево восстановило свою крону. В некоторые годы на нем образуются шишки, но семена в них не достигают зрелости.
Теперь обратимся к двум видам секции араукария, обитающим в Южной Америке. Араукария чилийская растет не только в Чили, но и в западной части Аргентины. Ареал этой араукарии расчленен на два изолированных участка. Меньший расположен на западном склоне Береговой Кордильеры на высоте около 700 м над уровнем моря. Протяженность этого ареала с 37°20' по 37°50' ю. ш. Более обширная часть ареала находится в Андах, Она тянется здесь вдоль хребтов между 37 и 40° на высоте 1600-1800 м, а на восточном склоне не спускается ниже 800-600 м над уровнем моря. Высказано предположение, что не все заросли араукарии чилийской естественного происхождения. Есть данные, позволяющие считать, что часть их приурочена к древним стоянкам индейцев. Широкое использование семян араукарий в пищу могло привести к распространению араукарий при посредстве человека, случайном или даже намеренном.
Араукария чилийская образует чистые или смешанные с южным буком (Nothofagus) леса вплоть до верхней границы леса. Лучше всего развиваются араукариевые леса на вулканических почвах. Наиболее густые заросли встречаются именно по склонам вулканов.
Араукария чилийская, как и араукария Бидвилла, - очень крупные деревья, достигающие (женские экземпляры) высоты 60 м при диаметре ствола до 1,5 м. Крона молодых деревьев широкопирамидальная, причем нижние ее ветви лежат прямо на земле. С возрастом нижние ветви обычно опадают. Боковые ветви взрослых деревьев расположены по 6-7 в мутовках, они горизонтально простертые или немного свисающие у старых деревьев; крона становится плоско-зонтиковидной, расположенной лишь на самой вершине ствола. Кора смолистая, толстая, продольно-трещиноватая.
Прирост молодых растений (до 30-150-летнего возраста) может достигать 45 см ежегодно, а затем уменьшается до 10-15 см. Этим араукарии резко отличаются от чрезвычайно медленно растущих агатисов.
Листья араукарии чилийской жесткие, колючие, темно-зеленые, спирально расположенные, покрывают ветви очень плотно друг к другу. Длина листьев 2,5-4 см, ширина 1,5- 2,5 см, по форме они яйцевидные с остроконечной верхушкой, к побегу прикреплены широким основанием, снизу - слегка килеватые. Живут листья до 40 лет.
Характерной особенностью араукарии чилийской являются ее микростробилы. Они пазушные, одиночные (но очень часто сближены в группы у самой верхушки ветви по 2-4-6), прямые, цилиндрические, иногда почти овальные, у основания окружены вегетативными листьями.
Микроспорофиллы с относительно длинной ножкой (до 8 мм). Надсвязник длиной до 15 мм, с сердцевидным основанием, на конце шиловидно суженный, вначале направленный кверху, а затем отогнутый вниз, поэтому поверхность микростробила кажется щетинистой; К нижней поверхности надсвязника прикреплены 12-16 двурядно расположенных микроспорангиев.
Шишки на коротких боковых ветвях, вегетативные листья которых постепенно переходят в чешуи шишек. Зрелые шишки коричневые, шаровидные, диаметром 12-18 см (они достигают массы 1,6 кг), вначале покрыты длинными, вытянутыми до 3 см и загнутыми кверху остриями кроющих чешуй, которые впоследствии обламываются.
Взрослые деревья дают 20-30 шишек, в каждой из которых содержится до 200-300 крупных семян. По созревании шишка рассыпается. Зрелое семя продолговатое, слегка сжатое, длиной 2,5-4 см, по краям семени заметны узкие каемки, остатки крыла.
Араукария чилийская светолюбива, растет в условиях влажного климата, на равномерно влажных, но незаболоченных, достаточно богатых питательными веществами почвах. Хорошо переносит и засушливые условия, а также небольшие морозы.
Крупные семена араукарии чилийской питательны и вкусны. Арауканы употребляют их в пищу в сыром или поджаренном виде. Древесина этой араукарии желтовато-белого цвета, ее используют в строительстве. Смола используется в народной медицине арауканов. Араукария чилийская - весьма ценное декоративное растение. Размножают ее как семенами, так и черенками.
В 1796 г., т. е. через 16 лет после открытия араукарии чилийской в Америке, она уже была интродуцирована в Англии (первое выращенное дерево просуществовало почти сто лет). Позднее эта араукария широко распространилась в Западной Европе (вплоть до Норвегии!), а в России - только в ботанических садах Крыма и Кавказа.
Другим южноамериканским видом является араукария бразильская, или узколистная (А. angustifolia). Растет она в горах Юго-Восточной Бразилии и Северо-Восточной Аргентины, образуя хотя и вырубленный в значительной степени, но все же довольно крупный массив чистых араукариевых лесов на высоте 500- 2000 м. По периферии сплошных араукариевых лесов араукария встречается и в других типах леса, но никогда не спускается до океанического побережья, хотя ее ареал почти точно повторяет - на некотором расстоянии - береговую линию.
Леса из араукарии бразильской (табл. 50) называют в Бразилии пинэйросами (португальское слово все от того же латинского корня "pin" - сосна), занимают значительную часть штата Парана (древесину этой араукарии так и называют "паранской сосной"). Здесь араукариевые леса соседствуют (по вертикали) с тропическими дождевыми лесами и сами содержат в подлеске некоторые типичные виды этих лесов. Однако собственно араукариевые леса в Бразилии нельзя рассматривать как тропические в климатическом отношении. На значительных площадях светлого (редкостойного) араукариевого леса его подлесок составляет сплошные заросли парагвайского чая, или мате (Ilex paraguariensis). Считается, однако, что распространение этого растительного сообщества обязано антропогенному фактору, а именно выпасу домашнего скота, уничтожившего первичный подлесок.

Таблица 50. Араукариевый лес в Бразилии (Рио-Гранде)
Араукария бразильская - важнейшее лесопромышленное дерево в Бразилии. Ее древесину широко использует местная промышленность как строительный материал, а также для изготовления мебели и всевозможных поделок и даже как сырье для бумаги. Используют ее при производстве музыкальных инструментов в Бразилии и других странах, в том числе в СССР (например, на Ленинградской фабрике).
К секции интермедия относят три новогвинейских вида араукарий. Наиболее примечательным видом является араукария Ханстайна (A. hunsteinii). Это - самая высокая араукария. Ее обычная высота 65-75 м. Крупнейший экземпляр араукарии Ханстайна, измеренный в 1941 г., имел высоту 88,9 м! Таким образом, это самое высокое дерево новогвинейских тропических лесов (если не самое высокое дерево тропиков!). Араукария Ханстайна входит в состав тропического дождевого леса в почетной роли дерева-эмергента - ее кроны возвышаются на 30 м и более над пологом леса, состоящего из цветковых растений. По существу, эмергенты составляют особый надъярус ("лес над лесом") дождевого леса. Араукария Ханстайна обитает на высоте от 500 до 2800 м над уровнем моря. При столь большой высотной амплитуде эта араукария входит также в состав других сообществ, в том числе и в "по-лувечнозеленые" леса, включающие листопадные виды кордии (Cordia), эритрины (Erythrina), терминалии (Terminalia), тооны (Тооnа) и других цветковых растений. С подъемом в горы араукария Ханстайна оказывается в составе горного дождевого леса с доминирующими кастанопсисом (Castanopsis) и дубом (Quercus). Выше этого леса араукарию Ханстайна часто замещает в лесных сообществах араукария Каннингема (A. cunninghamii), входящая в в секцию эутакта. Эта араукария, достигающая 60-метровой высоты и 3-метровой толщины ствола, выглядит весьма внушительно.
Араукария Каннингема встречается по всей Новой Гвинее и на острове Фергусон у северного берега Папуа. В Австралии растет на восточном побережье до реки Хастингс (33° ю. ш.) в северной части Нового Южного Уэльса. Ареал араукарии Каннингема протягивается больше чем на 30°. Это наибольшая широтная протяженность ареала среди видов араукарий. И одновременно араукария Каннингема наиболее северный вид араукарии; она растет у самого экватора, однако в северное полушарие нигде не заходит. Широка и климатическая амплитуда араукарии Каннингема. Она встречается на высоте от 90 до 2800 м над уровнем моря, обитая в тропических дождевых лесах, а в Австралии входит в состав субтропических дождевых лесов. На Новой Гвинее араукария Каннингема образует также почти чистые насаждения по хребтам, а на горе Дейман встречается во мшистом верхнегорном дождевом лесу (на высоте 2050-2450 м над уровнем моря) с участием ксантомиртуса (Xanthomyrtus) из миртовых и фикуса (Ficus).
По последним данным, араукария Каннингема прекрасно восстанавливает свои древостой в тех местах, где полог леса, состоящий из цветковых растений, несколько изрежен, например на крутых склонах, скалистых хребтах и в местах с нарушенным растительным покровом. Эти новые исследования, пожалуй, опровергают широко бытовавшее прежде мнение об араукариях, как о вымирающих растениях. В вымирании слишком явно повинен человек. Араукария Каннингема - долгоживущее растение; доживает она до 400-500 лет и, как многие долгожители, поздно достигает зрелого состояния. Поэтому процесс воссоздания уничтожаемых лесов очень долог. Он гораздо длительнее, чем при обычной лесоводческой практике. Для того чтобы ощутить результаты все расширяющейся деятельности лесоводов по созданию плантаций араукарии, на них надо смотреть глазами наших внуков или даже правнуков.
Местные жители на Новой Гвинее издавна использовали кору араукарии Каннингема в качестве строительного материала для своих хижин. Такие, лишенные коры экземпляры, естественно, погибали. Другой причиной гибели араукарий в лесах считаются пожары, в которых повинны люди. Несмотря на значительную влажность араукариевых лесов, пожары все же случаются, особенно в засушливые годы. Большая часть сохранившихся араукариевых лесов на Новой Гвинее расположена в ущельях, где огонь, вероятно, останавливал свое победное шествие. Сохранились леса и на нейтральных землях - участках леса, пограничных между враждующими племенами, а также там, где действуют табуистические запреты (в местах обитания "злых духов").
Другим и тоже знаменитым представителем секции эутакта является араукария разнолистная (A. heterophylla). Она широко известна как комнатное растение и в садоводстве под другим названием - араукария высокая (А. excelsa). Ее часто называют также "норфолкской елью". Это гигантские деревья, достигающие 70-метровой высоты при диаметре ствола 1,5-2 м. Крона узкопирамидальная или широкоцилиндрическая, с горизонтально простертыми ветвями по 4-7 в раздвинутых мутовках. Листья двух типов (рис. 188): игловидные - у молодых растений и густо черепитчато расположенные, чешуевидные - у взрослых (30-40 лет и старше). Перемена наряда (игловидных листьев на чешуевидные) так резко сказывается на внешнем облике этой араукарии, что во взрослом состоянии ее обычно не опознают.
Араукарию разнолистную размножают как семенами, так и черенками. Интересно, что из черенков, взятых с боковых ветвей, не образуется вертикального стебля и побеги растут только в горизонтальной плоскости. Она популярна и как декоративное растение, почему и распространена широко по оранжереям Европы.
Араукария колонновидная, или араукария Кука (A. columnaris), - примечательна во многих отношениях. Она обитает в южной тропической зоне на Новых Гебридах и на острове Сосновом (Новая Каледония). Впервые встречена была в 1774 г. на острове Сосновом капитаном Куком (тогда же была открыта и сама Новая Каледония). В 1786 г. немецкий натуралист Г. Форстер, участник экспедиции Кука, описал этот вид под названием "кипарис" колонновидный (Cupressus columnaris).
Стволы этих величественных деревьев равномерно одевает, от самого основания до вершины, узкая крона, отдаленно напоминающая крону пирамидального кипариса. Она образована сравнительно короткими ветвями, собранными в мутовки и отходящими от ствола почти под прямым углом (у кипариса ветви прижаты к стволу). На острове Сосновом араукария колонновидная образует прибрежные густые древостой, поражавшие своим видом первых путешественников, которые сравнивали их то с базальтовыми колоннами, то с дымящими фабричными трубами. На самой вершине дерева крона обычно несколько расширена (рис. 189).
Шишки араукарии колонновидной длиной до 10 см имеют щетинистый вид благодаря верхушкам чешуй, оттянутым в длинный (5-6 мм) шиловидный придаток, который отогнут книзу. По многим признакам араукария колонновидная похожа на араукарию разнолистную. У нее тоже происходит изменение формы листьев при переходе от юного состояния во взрослое. Но можно заметить и различия по листьям, на первый взгляд очень сходным: у араукарии колонновидной листья мягче, более выпрямленные, с постепенно суженной верхушкой (у разнолистной - внезапно суженные).
Араукария колонновидная широко распространена в культуре, но лишь в странах с тропическим, реже субтропическим климатом (в Австралии, Индонезии, Африке, даже в Италии). Ее можно увидеть и в оранжереях, например в Ботаническом институте в Ленинграде.
Этому роду араукариевых долгое время уделяли значительно меньше места и внимания в ботанической и популярной литературе, чем араукарии. Однако постепенно накапливался материал, и оказалось, что число реально существующих видов агатиса по крайней мере не меньше, чем видов араукарий, и по новейшим данным достигает двадцати. Впрочем, многие виды еще недостаточно изучены.
Сравнительно с араукарией род агатис еще очень молод: его ископаемые остатки не встречены в отложениях более древних, чем олигоценовые. Можно думать, что агатис возник около 30 млн. лет назад.
Все представители рода агатис - деревья, за незначительным исключением, крупные, достигающие высоты иногда 70 м и большой толщины ствола (2-3 м и более). Два вида агатиса имеют значительно меньшие размеры. Агатис желтеющий (A. flavescens) - дерево высотой обычно не более 12 м, иногда даже карликовюе, распространено в дождевых лесах центральной части полуострова Малакка (карта 14). Экземпляры агатиса яйцевидного (A. ovata), встречающегося в Новой Каледонии, редко превышают высоту 9 м.
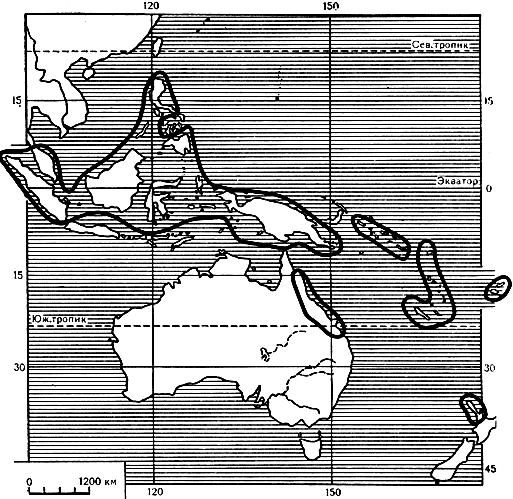
Карта 14. Ареал рода агатис
Ствол агатиса обычно колонновидный, лишь слегка утончающийся к вершине, на большом протяжении лишенный боковых ветвей. На уровне нижней трети или чаще середины общей высоты дерева ствол агатиса разветвляется на несколько крупных распростертых ветвей, образуя большую раскидистую крону. Ветви у молодых деревьев горизонтально простертые, часто мутовчатые.
В отличие от араукарий агатисы имеют хорошо выраженные округлые почки с крепкими, плотно прижатыми чешуями. У основания годичных побегов хорошо заметны рубцы от опавших почечных чешуй.
Листья на вертикальных побегах агатисов спирально расположенные, на боковых двурядные, супротивные или почти супротивные. Сохраняются листья до 15-20 лет. Молодые листья нередко розовые или красноватые, мягкие; позднее они приобретают темно-зеленый цвет и становятся кожистыми.
Зрелые листья обычно крупные, длиной 5- 18 см и шириной 1-6 см (рис. 192). До недавнего времени, пока еще не был открыт самый крупнолистый вид подокарпа, именно агатис по величине листьев считался чемпионом среди хвойных. Форма листьев у разных видов агатиса различна. Они могут быть и эллиптическими, и широколанцетовидными, и продолговатыми, и яйцевидными. Жилки листьев многочисленные, почти параллельные, у основания пластинки дихотомически ветвящиеся (рис. 193). Черешок короткий и плоский, или листья сидячие, без черешка. После опадения листьев на побегах остаются подушковидные рубцы.

Рис. 192. Ветвь агатиса белого (Agathis alba)
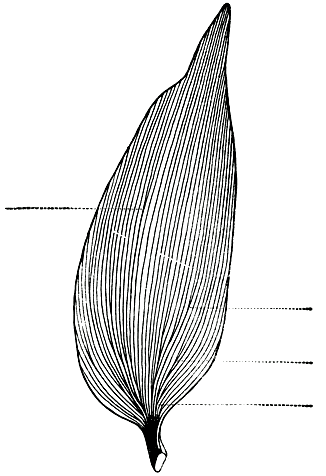
Рис. 193. Дихотомическое ветвление жилок листа агатиса белого (Agathis alba). Пунктиром показаны точки дихотомического ветвления
Устьица на листьях агатиса расположены рядами (рис. 194). Замыкающие клетки погруженные. Надустьичная ямка большей частью прикрыта, как и у араукарий, пробочкой из воскоподобного вещества (табл. 48).
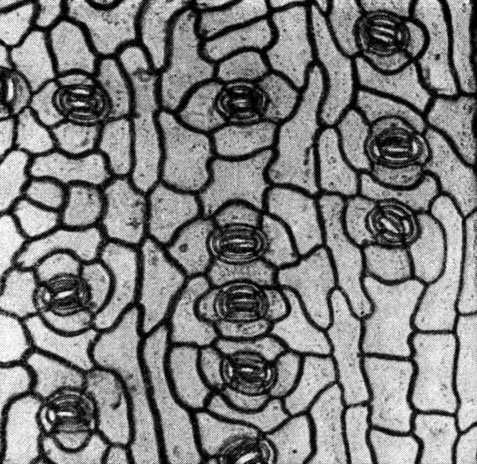
Рис. 194. Нижняя эпидерма листа агатиса Брауна (Agathis brownii)
Микростробилы пазушные, в том числе и те, которые кажутся надпазушными. Ложное впечатление "надпазушности" создается благодаря тому, что ножка микростробила, выходя из пазухи листа, в нижней своей части срастается со стеблем и лишь верхняя ее часть отклоняется от него. Форма микростробилов цилиндрическая, длина 2-6 см, ширина до 1 см, у основания - стерильные чешуйки.
Микроспорофилды на микростробилах расположены очень близко друг к другу, по спирали, сидят на коротких (длиной 3-5 мм) ножках. К нижней стороне надсвязника, имеющего полущитковидную форму с закругленным и отогнутым кверху краем, прикреплены свободно свисающие микроспорангии (их обычно 3-15).
Шаровидные шишки (считается, что латинское название агатиса, означающее в переводе с греческого "клубок ниток", дано растению именно из-за формы шишек) располагаются на концах коротких веточек. Форма шишек не всегда правильно шаровидная, она может быть даже широкоцилиндрическая (у агатиса туполистного - A. obtusa). Величина шишек у разных видов различна, их диаметр от 6 до 15 см.
Семенные чешуи агатисов полностью редуцированы. Кроющие чешуи, густо сидящие по спирали на толстой оси, плотно прилегают друг к другу, чем и объясняется гладкая поверхность шишки. Впрочем, агатис южный (A. australis) представляет исключение, так как его кроющие чешуи имеют короткие остроконечия, выступающие на поверхности шишки. В отличие от араукарий семязачатки агатиса не срастаются с чешуей. Зрелые шишки, как и у араукарий, рассыпаются по созревании. Семена агатисов довольно крупные (длиной 1-1,5 см), имеют одно хорошо развитое по боковой стороне крыло, образующееся из интегумента. С другой стороны семени иногда образуется небольшой пленчатый вырост, который может в редких случаях развиться во второе крыло, меньших размеров, нежели основное. Наличие таких приспособлений для полета, несомненно, способствует расселению агатиса (рис. 195).
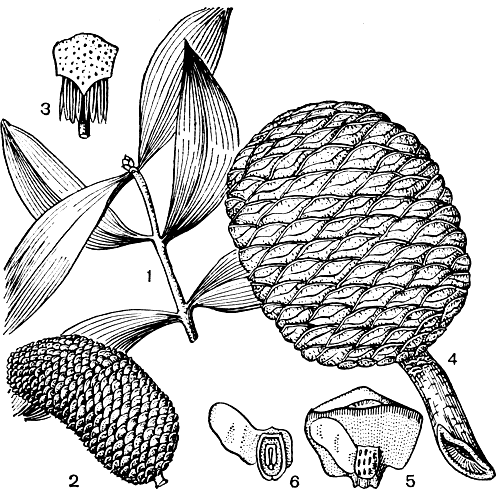
Рис. 195. Агатис крупноколосковый (Agathis macrostachys): 1 - ветвь; 2 - микростробил; 3 - микроспорофилл; 4 - шишка; 5 - семенная чешуя с семенами; 6 - семя с крылом в продольном разрезе
Древесина агатиса обладает весьма высокими техническими качествами и ценится даже выше древесины араукарий. Она прекрасно поддается обработке, очень эластична, в ней мало сучков. Поэтому область ее использования была и остается весьма широкой: кораблестроение, особенно во времена парусного флота, бондарное производство, строительство, изготовление мебели и т. д. На первом месте по ценности древесины среди видов агатиса, несомненно, стоит агатис южный - единственный вид рода, обитающий в Новой Зеландии. Его древесина не подвержена повреждению жуками-точильщиками. Именно поэтому она завоевала рынки многих стран. Но это все в основном в прошлом. Нынешний экспорт не может сравниться с экспортом середины прошлого века, когда еще строились деревянные корабли, а значительная часть Северного острова Новой Зеландии была покрыта лесами из агатиса южного.
В наши дни древесину агатиса южного применяют главным образом в тех случаях, когда трудно найти ей замену. Например, в производстве мебели ценится не только обычная древесина этого растения за ее приятную на ощупь текстуру, за прочность, долговечность и очень красивый цвет (от желто-белого до золотисто- коричневого), но даже в большей степени пятнистые и волнистые разновидности, обязанные этими качествами неравномерному накоплению смолы и неправильностям роста (довольно обычным в горных условиях).
Кроме древесины агатиса южного, промышленное значение имеет древесина других видов, но преимущественно на местных рынках. Ценна древесина австралийских агатисов Палмерстона (A. palmerstonii) и мелкоколоскового (A. microstachys), новокаледонских агатисов ланцетовидного (A. lanceolata) и Мора (A. moorei), а также обитателя Соломоновых островов, Новых Гебрид и островов Санта-Крус - агатиса крупнолистного (A. macrophylla).
Агатис может быть назван островным родом, так как его ареал, заходя лишь на края двух континентов (на полуостров Малакка в Юго-Восточной Азии и в штат Квинсленд в Австралии), охватывает преимущественно острова (Филиппины, Суматра, Калимантан, Сулавеси, Новая Гвинея, Соломоновы, Санта-Крус, Новые Гебриды, Новая Каледония, Фиджи и остров Северный в Новой Зеландии).
Несмотря на то что южная часть ареала агатиса заходит в Новую Зеландию, находящуюся в южной умеренной зоне, агатис по справедливости считают наиболее тропическим родом среди хвойных. Многие ботаники, правда, указывают, что хвойные в тропиках занимают главным образом горные местообитания. В отношении агатиса это верно лишь в том смысле, что тропические леса со значительным участием агатиса или даже с его доминированием в них и чистые агатисовые леса располагаются на средних и больших высотах в горах (до 2100 м). Но рассеянные одиночные экземпляры или небольшие группы деревьев агатиса встречаются и на морском побережье, исключая, впрочем, Малакку, Филиппины и Квинсленд, где агатис белый (A. alba) и агатис Брауна (A. brownii) отмечены в горах, лишь начиная с 200-500 м и выше.
Приуроченность многих агатисов к горным местообитаниям вряд ли можно свести к климатическим причинам (т. е. объяснить приверженностью этих растений, как и других хвойных, к более прохладным условиям в горах тропиков).
Старые экземпляры агатисов благодаря своим огромным размерам выступают над пологом дождевого леса, образуя как бы высокие зеленые холмы на равнине: равнина - это сплошной полог леса, а холмы - гигантские раскидистые, очень густые кроны агатисов, основание которых скрыто под лесным пологом, образованным цветковыми растениями. Например, на Ваникоро (острова Санта-Крус) высота сомкнутого полога леса составляет 15-20 м, тогда как самые высокие кроны растущего здесь агатиса крупнолистного достигают высоты 35-45 м.
Наличие таких эмергентов, т. е. гигантских деревьев, выступающих над высоким лесным пологом, вообще свойственно тропическому дождевому лесу.
Чаще всего в роли эмергентов оказываются различные цветковые растения, особенно представители семейства диптерокарповых (Dipterocarpaceae). Но местами хвойные в дождевых лесах являются либо полноценными компаньонами цветковых, либо доминируют в растительных сообществах, а самые крупные, просто гигантские хвойные (араукарии и агатисы) занимают в надъярусе такого леса места столь же крупных цветковых.
Растут агатисы чрезвычайно медленно. Зрелого состояния (с лесохозяйственной точки зрения) разные виды агатисов достигают не ранее 100-200 лет. Светолюбивые, буйно развивающиеся растения молодого вторичного дождевого леса быстро обгоняют в росте сеянцы агатисов, обрекая их на гибель. Каким же образом агатисы оказались в роли эмергентов первичного, т. е. зрелого, тропического леса?
Оказывается, для того чтобы стать полноправным членом дождевого леса, агатисы должны вступить в зрелый лес. Это удалось установить благодаря тщательному обследованию вулканического острова Ваникоро, произведенному английским ботаником Т. Витмором. На низкой высоте, где остров полого спускается к океану, в изобилии растут сеянцы и молодые растения агатиса крупнолистного. На средней высоте, где на крутых склонах развит дождевой лес, агатис представлен почти нацело эмергентными довольно редкими экземплярами или небольшими группами. Витмор объясняет это тем, что агатис лучше развивается на нарушенных почвах. Остров Ваникоро лежит в зоне ураганов, во время которых крупные деревья в лесу, особенно на горных склонах, могут быть вырваны с корнем. Кроме того, на Ваникоро ощущаются постоянные подземные толчки, ускоряющие оползневые процессы. Обильные дожди способствуют размыву обнаженных участков. Почва, таким образом, оказывается нарушенной, конкуренция агатису со стороны других растений ослабевает, и агатисы получают возможность партиями заселять зрелый дождевой лес.
Агатисы можно различать, например, по листьям. Так, у агатиса южного они серо-зеленые, на старых экземплярах толстые, кожистые и тесно расположенные на побеге длиной 3-7 см, шириной до 1 см, а у сходного с ним агатиса туполистного (A. obtusa) листья сверху блестящие, темно-зеленые, продолговато-эллиптические, с округлой верхушкой и гораздо крупнее (длиной до 12 см и шириной до 3 см).
Сходные между собой агатисы ланцетовидный и агатис Мора имеют некоторые отличия по величине и форме листьев. У первого из них листья заостренные, у второго - притуплённые. Агатис ланцетовидный имеет листья шириной до 1,5 см при длине их 4-12 см. У агатиса Мора ширина листьев 1,5-4 см, длина до 10 см. Но особенностью агатиса ланцетовидного являются все же не листья, а супротивные цилиндрические микростробилы (длиной 2,5 см и диаметром 0,8 см) на коротких толстых деревянистых нож-ках. Большинство остальных видов агатиса, в том числе и агатис Мора, имеют одиночные микростробилы.
У разных видов агатиса существенно различно число микроспорангиев. Так, у южного их 10-12, у белого - 5-12, а у агатиса Брауна микроспорофиллы несут только 3 микроспорангия.
К агатису крупнолистному, наиболее хорошо изученному в лесах острова Ваникоро, близок агатис белый, широко разводимый в оранжереях мира (чаще всего он значится под названием либо агатиса даммара - А. dammara, либо агатиса ремнецветниколистного - A. loranthifolia). Агатис белый растет в тропических лесах на полуострове Малакка, на Филиппинах, Суматре, Сулавеси, Калимантане, на Молуккских островах, на Новой Гвинее. Это наиболее широко распространенный вид агатиса, его ареал лежит между 10° с. ш. и 8° ю. ш., т. е. в экваториальной зоне. Именно агатис белый обладает наиболее широкими листьями.
В пределах ареала агатиса белого указывается еще несколько видов агатиса, но только часть их вполне самостоятельны, остальные же, например агатис целебесский (A. celebica), агатис борнеоский (A. borneensis), агатис филиппинский (A. philippinensis), являются лишь разновидностями агатиса белого. Уже был упомянут агатис желтеющий, названный так потому, что его экземпляры, растущие на открытых плоскогорьях полуострова Малакка, бросаются в глаза желтым цветом своей листвы. В лесах его листья приобретают зеленый цвет.
Особняком в экологическом отношении стоит низкорослый агатис яйцевидный. Этот агатис чаще встречается как одиночное дерево или образует небольшие древостой по сухим горным склонам от 450 м до уровня океана на юге Новой Каледонии и примыкающем к ней острове Сосновом. В северной части Новой Каледонии обитает агатис Мора, а в южной - агатис ланцетовидный, близкие друг к другу лесные виды. Еще очень слабо изучены два других новокаледонских вида - агатис горный (A. montana) высотой 15-20м и агатис Корбассона (A. corbassonii) высотой до 40 м.
Три вида - агатис Брауна, близкий к нему агатис Палмерстона (A. palmerstonii) и агатис мелкоколосковый (A. microstachys) - обитают на востоке Австралии, в Квинсленде. Последний занимает прибрежные местообитания в тропическом нижнегорном лесу Северного Квинсленда. Все австралийские агатисы - очень крупные деревья (высотой 35-60 м).
Кроме того, в Австралии, в ботанических садах Сиднея и Мельбурна и просто в городских посадках, широко культивируются с декоративной целью различные виды агатиса, например агатис Мора с его пятнистой, как у платана, корой, а далеко на юго-западе Австралии, в Перте, - восточноавстралийский агатис Брауна.
Самый восточный вид - агатис фиджийский (A. vitiensis) - достигает границы восточного и западного полушарий (на острове Вануа-Леву). Он растет в тропическом дождевом лесу островов Фиджи, выступая в качестве дерева, доминирующего в верхнем пологе. Обычная высота его 15-18 м, но отдельные экземпляры достигают высоты 30 м. На Фиджи древесину этого вида издавна использовали для различных строительных целей (лодки, прибрежные сооружения и дома). Черную краску, получаемую из сажи этого растения, помимо ее самостоятельного значения, фиджийские женщины используют в смеси с красной землей как коричневый пигмент для татуировки. В Австралии агатис фиджийский разводят в качестве декоративного растения.
Наиболее известным видом агатиса является самый южный, так и называемый агатисом южным, или каури (A. australis). Он обитает на севере Северного острова Новой Зеландии, в районах Нортленд и Сентрал-Окленд. Название "каури" новозеландское, точнее, маорийское, распространилось впоследствии, по мере познания ботаниками рода агатис, и на другие его виды, которые стали называть "каури" с прибавлением того или иного географического эпитета: фиджийский каури, ваникоро-каури, квинсленд-каури (агатис Брауна), северный квинсленд-каури (агатис мелкоколосковый), северный каури (агатис Палмерстона) и др.
Голландский мореплаватель Абель Тасман, в середине XVII в. открывший Новую Зеландию, увидел острова, сплошь покрытые величественными лесами. Вероятно, такой же страна предстала и Куку, вторично открывшему ее в 1769 г. Коренные жители Новой Зеландии, полинезийское племя маори, за двести (или более) лет до Тасмана переселившиеся сюда с островов Полинезии, были мореплавателями, строили свои суда из местных деревьев, в том числе из каури, строили жилища, добывали смолу каури для освещения. Но процесс сведения лесов был очень медленным до середины XIX в., пока Новая Зеландия не была включена официально в состав Британской империи (1840). Колонизация Новой Зеландии с этого времени стала очень бурной (ей не помешали и маорийские войны 1843-1872 гг.). Новопоселенцы расчищали лесные площади под пашни и пастбища. Лесных великанов - каури - добывали выборочно из-за прекрасных качеств их древесины. Поэтому уже в конце XIX в. появилась необходимость в возобновлении лесов, и тогда же были созданы первые плантации каури.
Каури - "царь новозеландских лесов" - величественное дерево, достигающее высоты 60 м. Двух- и даже трехметровая толщина ствола каури обычна для крупных экземпляров. Максимальной указывается толщина в 7 и 10 м. Многим это кажется преувеличением, но ниже мы приведем некоторые конкретные данные о размерах стволов каури, с тем чтобы читатель мог яснее представить себе облик этого удивительного растения.
Стволы каури поднимаются как мощные гладкие сизо-серые колонны, лишь незначительно утончающиеся в месте первого ветвления. Лишенные ветвей колоннообразные стволы достигают высоты 18-20 м, а в исключительных случаях и более. Крона взрослого каури очень обширна, раскидиста. Молодые растения имеют иной облик. Даже в возрасте 20-40 лет это еще невысокие деревца с тонкими боковыми ветвями, отходящими от ствола по всей его высоте. А достаточная для лесоводческой практики толщина ствола достигается лишь в возрасте 150-200 лет. Трехсотлетние экземпляры могут иметь ствол 1,5-метровой толщины.
У каури самые мелкие и узкие листья среди агатисов. У молодых экземпляров и на молодых ветвях листья длиной 3-7 см и шириной до 1 см, с тупой верхушкой, суженные к основанию, серовато-зеленые. На старых деревьях листья толстые и короткие (длиной до 5 см), чуть более широкие (но редко превышают 1 см), зеленого цвета.
В настоящее время в Новой Зеландии сохранились лишь остатки субтропических дождевых лесов, в которых каури был доминирующим растением. Эти леса располагаются на высоте до 600 м. Выше их сменяет широколиственный лес с подокарпом тотара (Podocarpus totara) в качестве доминирующего растения.
Известны три резервации агатисового леса (леса каури) в Новой Зеландии. Это лес Ваитекере в окрестностях Окленда и два массива в Нортленде, на западном берегу острова Северного: севернее 36-й параллели - Парк-Траунсон и самый большой лес - Ваипоуа. Лес Ваипоуа, имеющий статус арборетума, занимает площадь 11 тыс. га собственно леса каури, а вместе с плантациями - 15,5 тыс. га.
Наиболее знаменитые экземпляры каури носят в Новой Зеландии личные имена, и к ним, как и к мамонтовым деревьям (Sequoiadendron) в Калифорнии, прикрепляют дощечки с этими именами. Как раз в лесу Ваипоуа есть дерево, носящее маорийское имя Тане Махута - "Первое воплощение Тане": маори считают, что это дерево - лесной бог Тане. Но каури не случайно связан с именем божества. По представлениям маори, Тане, сын Ранги (Неба-отца) и Пэпы (Земли-матери), оказался самым могучим среди своих братьев и самым "человечным". Он настоял на том, чтобы лишь оторвать небо от земли, не убивая их: ведь это были его родители. Он сделал так, и потоки света и воды (слезы Ранги) обрушились на богов. Вдохнув вольного воздуха, они принялись строить новый мир. Тане решил украсить Землю-мать своими детьми - деревьями. Правда, вначале (ведь все создавалось впервые) он посадил деревья вверх корнями. Поняв свою ошибку, Тане вырвал гигантский каури и прочно вставил его корнями в землю. Потом он с гордостью взглянул на чудесную зеленую крону над гладким прямым стволом. Шелест листьев звучал, как музыка. . . Мир становился прекрасным.
Толщина ствола при основании Тане Махута около 4 м, высота неразветвленной части дерева 13 м. Считается, что возраст этого гиганта - 1200 лет.
Новозеландский ботаник В. Чапмен, который приводит эти цифры, подсчитал, что дерево Тане Махуте дало бы 160 м3 прекрасной древесины (только из колоннообразной части дерева). Надо еще добавить, что условия для роста Тане Махута отнюдь не идеальны: он высится одиночкой в болотистой местности - все остальные экземпляры здесь вымерли (каури обычно не переносит заболачивания).
Тане Махута, однако, не самое старое и не самое крупное дерево в лесу Ваипоуа. Другой гигант - Те Матуа Нгаере (Отец леса) - имеет ствол диаметром 5 м (16 м в обхвате). Описание же гиганта из бухты Меркьюри (на севере залива Изобилия) приводит к мысли, что указания на 7-метровую и даже 10-метровую толщину стволов каури могут быть вовсе не фантастическими. Это дерево при высоте 45 м, т. е. далеко не максимальной, достигло более чем 7-метровой толщины ствола. Мутовка толстых ветвей отходит на высоте 24 м над землей - только отсюда начинается величественная крона. Толщина ствола в этом месте около 3 м. Надо еще добавить, что первые ветви этого экземпляра - толщиной с хорошее дерево - до полуметра и более.
Возраст каури из бухты Меркьюри знатоки флоры Новой Зеландии оценивали по-разному (Т. Кирк - 4000, Т. Чизмен - 1700 лет). Надо думать, что в период бесконтрольного уничтожения новозеландских лесов предприниматели могли специально охотиться за особо крупными экземплярами, каждый из которых мог дать им столько древесины, сколько можно получить с полугектара обычного соснового леса. Поэтому и 10-метровый диаметр ствола можно считать вполне достоверным сообщением.
Сейчас в Новой Зеландии не только бережно охраняют в резервациях остатки агатисовых лесов, но и специально изучают возможности естественного и искусственного их возобновления.
Известно, что леса имеют водоохранное значение. Именно этому обстоятельству обязан своему сохранению лес каури в окрестностях Окленда - самого крупного города Новой Зеландии. Этот лес, расположенный на холмах Ваитекере, был подвергнут специальному изучению. Во-первых, было выяснено, что медленная естественная регенерация леса возможна при условии специальной охраны леса. Но процесс, длящийся несколько столетий, нельзя изучить экспериментально (точнее, можно, но усилиями многих поколений исследователей). Поэтому ботаники исследовали разновозрастные, существующие одновременно сообщества леса каури в Ваитекере. По их мнению, лес каури заселяет пустоши в течение 200-300 лет в такой последовательности. После короткого "травяного" периода следует кустарниковая фаза - фаза заселения местности "чайным деревом" (Leptospermum scoparium) из миртовых. Самый нижний ярус такой кустарниковой заросли оказывается плотно заселенным метровой высоты папоротником - глейхенией скрученной (Gleichenia circinata). Под папоротниковым пологом развивается много сеянцев лесных деревьев, в том числе каури. Затем папоротник отмирает, "чайное дерево" сменяется другим видом того же рода (Leptospermum ericoides), но его начинает перерастать каури. Следующей стадией является зрелая стадия леса с доминирующими вполне взрослыми каури.
Было установлено, что каури не возобновляется в зрелом дождевом лесу. При заболачивании леса погибают и взрослые экземпляры, и сеянцы. Но семена не прорастали и сеянцы гибли и в других, казалось бы, благоприятных условиях. Пытались найти этому внешние причины. И действительно, заметили поедание листьев сеянцев насекомыми, различные грибные поражения ("вымокание" сеянцев вскоре после прорастания, повреждение корневой шейки сеянцев). Но оказалось также, что большое количество упавших на землю семян поедают животные. Особенно большой ущерб наносит насекомое из прямокрылых - вета (Hemideina thoracica), с помощью своих сильных мандибул разгрызающая семена и поедающая маслянистое содержимое (кстати, семена некоторых австралийских агатисов употребляют в пищу и люди). В истреблении семян повинен также местный попугай (Nestor occidentalis). Его вина даже больше, чем веты, ибо он извлекает клювом семена из шишек, которые после этого опадают недозрелыми. Шишки опадают в незрелом состоянии и вследствие поражения песталотией погребальной (Pestalotia funerea) из несовершенных грибов.
Обнаружены и более существенные причины слабого естественного возобновления каури. Эксперименты и наблюдения в лесу и в лаборатории показали, что его семена быстро теряют жизнеспособность при высокой (выше 65%) относительной влажности воздуха. В то же время для прорастания семян почва должна быть достаточно влажной. Шишки каури созревают поздним летом, и опад семян происходит с конца февраля до середины апреля. Относительная влажность воздуха в эту пору превышает 65%, и большое количество семян теряет способность к прорастанию. А в поверхностных слоях почвы в это время, как это ни парадоксально, влаги для прорастания семян каури недостаточно.
По-видимому, каури все же является вымирающим растением, и человек лишь подтолкнул его к гибели. Правда, новозеландцы вовремя спохватились и объявили каури охраняемым растением. Конечно, о заготовке древесины в больших количествах сейчас нет и речи. Более того, ограничен также сбор смолы каури, прежде столь популярной и у маори, и во всем мире. Вывоз смолы каури, еще в 20-х годах XX в. достигавший 15 000 т в год, ныне резко упал. Достаточно сказать, что в лесу Ваипоуа собирать естественно вытекающую смолу имеет право только один человек. О повреждении стволов, "подсочке", и говорить нечего - она была запрещена намного раньше.
Смолу каури называют еще каури-копалом. Слово "копал" (исп. copalli), взятое из языка одного индейского мексиканского племени, означает вообще любую твердеющую на воздухе смолу растительного происхождения. Во всем мире производят несколько десятков разновидностей копала (большей частью из камедей различных цветковых растений, обычно тропических). Но и виды агатиса внесли значительную лепту в общий копаловый рынок. Правда, некоторые названия копалов, например даммара-копал, являются источником путаницы. Дело в том, что "Dammara" - это синоним названия как рода хвойных - агатиса (Agathis), так и рода цветковых - протиума (Protium) из семейства бурсеровых (Burseraceae). Эта путаница с названиями растений произошла потому, что малайское слово "даммар" означает "смола". Поэтому и получается, что даммара-копал может быть производным как агатиса белого, так и растений из рода протиум, из которых действительно получают копал. Многие затвердевшие смолы - копалы - получают свои названия по месту добычи. Таков манила-копал, добываемый на Филиппинах из разных растений, в том числе и из агатиса. Каури-копал является исключением, так как во всех случаях представляет собой застывшую смолу именно каури (агатиса южного). Конечно, подсочка ствола каури - самый простой способ добычи копала, но деревья погибали после многократных поражений ствола. И боязнь утратить свою национальную гордость - каури - привела к запрету подсочки. После этого главным способом добычи копала оставался очень опасный - сбор естественных натеков, главным образом из развилок ветвей. А как мы помним, первая развилка у каури находится на большой высоте. Учитывая толщину ствола, который не обхватишь руками, можно понять трудность и опасность такой работы. Поэтому сборщики копала обратились к поискам самой драгоценной его разновидности - ископаемой. Различают копалы полуископаемый (четвертичного периода) и собственно ископаемый (более древний, чаще всего третичного периода). Полуископаемый копал добывают из почв обычно на месте сведенного леса каури, но также и в существующих еще лесах. Смола стекает по стволам живых каури или вытекает из поваленных деревьев, проникает в почву и оказывается погребенной. Копалы, пролежавшие в земле долгие годы, иногда многие тысячелетия и даже миллионы лет, приобретают вид янтаря и часто служат для его имитации. Последнему способствует то, что в смоле каури, так же как в янтаре, нередки включения: листья, насекомые. Цвет каури-копала охватывает цвета всех разновидностей балтийского янтаря: от бледно-лимонно-желтого до красновато-коричневого и даже черного.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'