Репродуктивная система: стробилы и спорангии
Так же как у кордаитовых, стробилы у хвойных всегда однополые. Они образуют собрания стробилов, или сложные стробилы, называемые иногда "соцветиями". К сожалению, нет специального термина для обозначения собраний стробилов, так как термин "соцветие" по понятным причинам сюда не подходит. Американский палеоботаник Дж. Уиланд предложил термин метастробил, но он не привился. Собрания стробилов хвойных соответ-ствуют сережковидным собраниям кордаитовых. Но в то время, как у кордаитовых строение этих собраний стробилов довольно ясно и понятно, у хвойных оно сильно затемнено далеко зашедшими процессами редукции и срастания частей. В результате редукции число стробилов в собраниях мегастробилов ("соцветиях") и особенно в собраниях микростробилов нередко сведено до одного. Так, у тисса сохранился лишь один мегастробил (к тому же до крайности редуцированный), а у сосны мы имеем лишь одиночные микростробилы.
Происхождение и эволюция стробилов хвойных представляют собой одну из наиболее увлекательных глав морфологии растений. Этому вопросу посвящена огромная литература на разных языках, и, хотя многое уже прояснилось, осталось также немало спорного.
Относительно легче поддается выяснению вопрос об эволюции микростробилов. Этому особенно помогает то обстоятельство, что, как показала американский ботаник Мэри Уайлд (1944), относительно довольно примитивные, почти кордаитоподобные, собрания микростробилов сохранились у двух видов подокарпа, один из которых произрастает в Новой Зеландии (подокарп колосистый - Podocarpus spicatus), а другой - в Южном Чили (подокарп андийский - P. andinus). Небольшие микростробилы сидят у них в пазухах кроющих чешуй на главной оси "сережки", как у кордаитовых. Ось обычно заканчивается верхушечным микростробилом. У остальных видов подокарпа наблюдается постепенное уменьшение числа боковых микростробилов на оси "сережки" и в конце концов остается лишь один верхушечный микростробил. Но даже такой одиночный микростробил у некоторых видов подокарпа (так же как у торреи - Torreya) несет у своего основания пустые (стерильные) кроющие чешуи - живые свидетели того, что в прошлом число микростробилов было больше. Число таких стерильных чешуй невелико, всего 4-6, но для морфолога их наличие является важным аргументом в пользу гипотезы редукции. Но виды подокарпа не являются единственными, где сохранились относительно примитивные "мужские соцветия" со многими или несколькими микростробилами. Они имеются и у других родов (сциадопитис, австротаксус, головчатотисс и аментотаксус), но у них они более видоизмененного типа, чем у упомянутых выше видов подокарпа. У всех остальных хвойных в результате редукции первоначально сережковидного собрания сохранились лишь одиночные микростробилы. Эти одиночные микростробилы морфологически соответствуют верхушечному микростробилу в "сережках" упомянутых выше видов подокарпа и кордаитовых. Таково, в частности, происхождение одиночного микростробила тисса, как это убедительно показала в 1975 г. М. Уайлд.
Микростробил (часто называемый "мужским колоском") состоит из укороченной оси, на которой расположены сильно редуцированные микроспорофиллы. Расположение микроспорофиллов у большинства хвойных спиральное (очередное), но у кипарисовых микроспорофиллы расположены всегда супротивно или в мутовках по три.
Форма микроспорофиллов хвойных очень различна. Но все их разнообразие можно свести к двум основным типам - дорсивентральному (как у сосны) и радиально-симметричному (как у тисса). Уже давно возник вопрос, какой же из этих двух типов является более примитивным, т. е. исходным в процессе эволюции. Уже знаменитый чешский ботаник Л. Челаковский (1898), а вслед за ним английский ботаник У. Уорсделл (1901), немецкий ботаник Карл Гёбель (1923), Р. Флорин (1948) и ряд других исследователей считали, что радиально-симметричный (периспорангиатный) микроспорофилл типа тисса является более примитивным, а плоский (гипоспорангиатный) микроспорофилл типа сосны возник в результате абортирования адаксиальных спорангиев и дорсивентрализации всего спорофилла. При этом В. Циммерман (1930, 1959), Р. Флорин (1948, 1951) и ряд других ботаников в качестве одного из решающих аргументов использовали теломную теорию и считали, что микроспорофилл большинства хвойных произошел из радиального спорангиального "пучка" с верхушечными спорангиями, а микроспорофилл типа тисса считали промежуточной формой между этой исходной формой (отчасти сходной с микроспорофиллом кордаитовых) и обычным, дорсивентральным микроспорофиллом. Гипотеза эта кажется довольно привлекательной. Однако она все же оказалась не вполне обоснованной.
Еще в 1896 г. французский ботаник Э. Тибов своей книге о "мужском аппарате" голосеменных, на основании детального изучения анатомии проводящей системы микроспорофиллов саговниковых и хвойных пришел к выводу, что радиальный микроспорофилл тисса является наиболее подвинутым среди хвойных. Позднее американский ботаник P. Tомсон (1939, 1940) привел дополнительные доказательства подвинутости радиально-симметричного микроспорофилла тисса и пришел к выводу, что такой радиальный спорофилл возникает в результате слияния двух дорсивентральных микроспорофиллов. Наконец, в 1975 г. по этому вопросу опубликовала чрезвычайно важную и интересную статью уже упомянутая нами Мэри Уайлд. На основании тщательного исследования внешней морфологии и анатомии микроспорофиллов тиссовых и головчатотиссовых (Cephalotaxaceae) ей удалось убедительно показать, что радиально-симметричный тип микроспорофилла происходит в результате срастания (в процессе эволюции) двух или больше верхушечных плоских микроспорофиллов, причем имеются все переходные формы.
Перейдем теперь к мегаспорангиатным ветвям стробилов, т. е. к тем их фертильным ветвям, на которых расположены семязачатки. Направление их эволюции представляет исключительный интерес как само по себе, так и для систематики и филогении хвойных. Но в то же время здесь очень много сложных вопросов, причем далеко не все они нашли полное разрешение. Можно полагать, что мегаспорангиатные ветви морфологически вполне соответствуют микроспорангиатным ветвям и подобно последним являются продуктом специализации сережковидных собраний стробилов предковых форм. Наиболее примитивный тип организации мегаспорангиатной ветви наблюдается у тех же двух видов подокарпа, у которых M. Уайлд установила примитивный тип собрания микростробилов. У них на главной оси сережковидной ветви сидят мелкие, спирально расположенные кроющие чешуи, в пазухах которых (в верхней части ветви) - по одному очень редуцированному мегастробилу. У остальных видов подокарпа в результате редукции числа стробилов на оси "сережки" остается лишь один верхушечный или почти верхушечный мегастробил.
Уменьшение числа мегастробилов до одного не является единственным направлением эволюции. У подокарповых наблюдается и другое эволюционное направление, которое выражается не столько в уменьшении числа отдельных мегастробилов, сколько в сокращении длины междоузлий главной оси и в образовании таким образом компактного собрания мегастробилов, т. е. того образования, которое мы называем женской шишкой или просто шишкой. Среди подокарповых такую шишку, правда, довольно маленькую, имеет, например, саксеготея (Saxegothaea).
Эти два направления эволюции мегаспорангиатной ветви мы наблюдаем в двух близких семействах - тиссовых и головчатотиссовых. У тисса и псевдотаксуса, или ложного тисса (Pseudotaxus), в результате далеко зашедшей редукции числа мегастробилов мы имеем одиночные односеменные мегастробилы. У аментотаксуса, или сережчатотисса (Amentotaxus), и цефалотаксуса, или головчатотисса (Cephalotaxus), возникли маленькие шишки. Эволюция всех остальных хвойных шла в сторону возрастания компактности собрания мегастробилов, уменьшение же числа частей играло второстепенную роль. В итоге возникли компактные шишки - типа шишек сосны, ели, араукарии и огромного большинства других хвойных.
Каждая шишка, например, всем читателям хорошо известная шишка сосны, состоит из центральной оси, на которой сидят кроющие чешуи, в пазухах каждой из которой находится так называемая "семенная чешуя". Их называют "семенными", потому что на своей верхней стороне они несут семязачатки. Но хотя функция семенной чешуи вполне понятна, вопрос о ее происхождении долгое время служил предметом больших разногласий и породил огромную литературу. Мало какому вопросу морфологии растений уделялось так много внимания и посвящено так много работ, как проблеме семенной чешуи хвойных. Что же такое семенная чешуя?
Прежде всего совершенно ясно, что семенная чешуя - это не листовой орган, не спорофилл и ничего общего не имеет с плодолистиком. Это было ясно уже немецкому ботанику Маттиасу Шлейдену (1839), хорошо известному своими работами о клеточном строении растений. Он протестовал против идеи знаменитого английского ботаника Роберта Брауна (1827), толковавшего семенную чешую как открытый "плодолистик", и справедливо указывал на то, что листовой орган никогда не возникает в пазухе другого листа (в данном случае в пазухе кроющей чешуи). Но если семенная чешуя не есть листовой орган, то, следовательно, морфологически он представляет собой побег, пусть даже очень видоизмененный и специализированный. Когда немецкий ботаник и натурфилософ Александр Браун исследовал уродливые (аномальные) шишки лиственницы, где кроющие чешуи были превращены в вегетативный лист, а семенная чешуя - в короткий побег с двумя листьями, а также аналогичные аномалии у других хвойных, он сделал в 1842 г. вывод, что семенная чешуя соответствует двум первым сросшимся листьям редуцированного побега. Таким образом, А. Браун выдвинул идею "побеговой" природы семенной чешуи и тем самым сильно приблизился к решению этой трудной морфологической проблемы. Он был первый, кто считал шишку хвойных целым "соцветием", а не простым "цветком" (мы бы сказали стробилом, а не "цветком", но здесь нет очень большой ошибки).
Выдвинутая А. Брауном "теория пазушного брахибласта", как ее назвали впоследствии, была развита рядом последующих исследователей, из которых нужно в первую очередь назвать Ладислава Челаковского, опубликовавшего целый ряд интереснейших работ (1879-1900). Его обширные исследования и остроумные построения помогли прочно обосновать основную идею А. Брауна и способствовали широкому распространению "теории пазушного брахибласта" среди ботаников. Главным аргументом в пользу этой теории для Л. Челаковского служили случаи замещения семенной чешуи коротким побегом в аномальных шишках лиственницы, а также сосны, ели и др. Путем исследования большого материала ему удалось обнаружить постепенные переходы между первыми двумя листьями пазушного побега аномальных шишек и нормальной семенной чешуей. Кроме того, Челаковский показал, что у сосны третий передний лист пазушного побега аномальных шишек, поворачиваясь на 180 принимает участие в образовании семенной чешуи вместе с первой парой листьев.
Но о чем говорит строение обычной, нормальной семенной чешуи? Не несет ли она в своем внутреннем, анатомическом строении какие-либо признаки, указывающие на "брахибластную" ее природу? На этот вопрос попытался ответить известный американский анатом Артур Имс (1913). Детальное исследование проводящей системы семенной чешуи привело его к выводу, что она представляет собой не что иное, как укороченный побег (брахибласт), расположенный в пазухе кроющей чешуи. Из оси шишки в каждую кроющую чешую отходят проводящие пучки, в которых взаимное расположение ксилемы и флоэмы такое же, как в листе. В тех же проводящих пучках, которые снабжают семенную чешую, расположение ксилемы и флоэмы обратное. В процессе эволюции хвойных кроющая и семенная чешуя полностью или почти полностью сливаются в одну простую чешую, которую можно очень хорошо видеть, например, у араукариевых. Но двойная природа таких "простых" чешуй доказывается присутствием двух серий проводящих пучков - нижней, с нормально ориентированной ксилемой и флоэмой, и верхней, с обратным их расположением, а также наличием у чешуи двух свободных верхушек. К сходным выводам и также на основании анатомических исследований пришел другой американский ботаник - Э. Синнот (1913). Семенную чешую он сопоставлял с язычком араукариевых и так называемым эпиматием подокарповых и привел ряд убедительных доводов в пользу их гомологичности. В дальнейшем появляется целая серия работ, в которых "теория брахибласта" обосновывается и детализируется на основании привлечения все более широкого фактического материала.
Свое окончательное подтверждение теория брахибласта получила со стороны палеоботаники. В замечательных исследованиях Р. Флорина (1939, 1951, 1954) мы находим интереснейшие данные о строении шишки палеозойских хвойных, проливающие яркий свет на вековую проблему семенной чешуи. Наиболее примитивными и поэтому для решения данной проблемы наиболее интересными оказались шишки палеозойской лебахии сосковидной (Lebachia piniformis, рис. 185) из вымершего семейства вальхиевые (Walchiaceae). Шишки лебахии несут многочисленные вильчатые кроющие чешуи, в пазухах которых сидит по одному редуцированному и более или менее радиально-симметричному мегастробилу. Каждый такой мегастробил состоит из короткой оси, на которой расположено несколько стерильных чешуй и один редуцированный чешуевидный мегаспорофилл. Мегастробил лебахии значительно более редуцирован и упрощен, чем мегастробилы кордаитовых, но по сравнению с мегастробилами позднейших хвойных он чрезвычайно примитивен. Он занимает как бы промежуточное положение между мегастробилами кордаитовых и хвойных и фактически представляет собой ту исходную структуру, из которой в результате дальнейшей специализации образовалась семенная чешуя. В отличие от кордаитовых мегастробил лебахии обнаруживает некоторую, хотя еще слабо выраженную тенденцию к дорсивентрализации. Эта тенденция более ясно выражена у близкого к лебахии пермо-карбонового рода эрнестиодендрон (Ernestiodendron, рис. 186). Мегастробилы у этого рода уже более или менее уплощенные и с более редуцированной осью, но в то же время мегаспорофиллов здесь больше (от 3 до 7). У всех же более молодых палеозойских хвойных мегастробилы явственно дорсивентральные, более приближающиеся к семенной чешуе современных хвойных. Так, у верхнепермского рода псевдоволmции (Pseudovoltzia) имеется примитивная пятилопастная семенная чешуя, которая занимает как бы промежуточное положение между мегастробилами лебахии и эрнестиодендрона и более упрощенными и редуцированными семенными чешуями позднейших хвойных. Еще большая редукция мегастробила наблюдается у верхнепермского рода ульмания (Ullmannia).
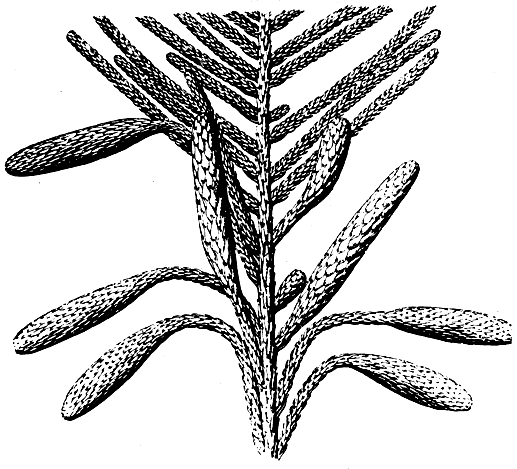
Рис. 185. Лебахия сосновидная (Lebachia piriformis); боковая ветвь с прямостоячими собраниями мегастробилов и висячими микростробилами
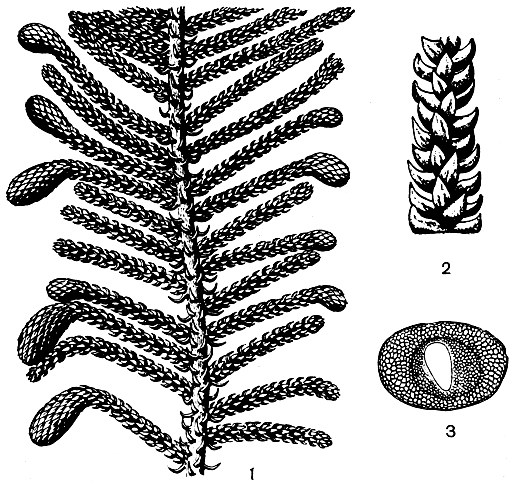
Рис. 186. Эрнестиодендрон папоротниковидный (Ernestiodendron filiciforme): 1 - ветвь с собраниями микростробилов на концах боковых побегов; 2 - часть облиственной веточки; 3 - пыльцевое зерно
Таким образом, уже в пермском периоде существовали хвойные с довольно специализированным и более или менее чешуевидным мегастробилом. Дальнейшие изменения происходят у мезозойских хвойных. В частности, у триасового рода вольция (Voltzia) и верхнетриасового и нижнеюрского рода схизолепис (Schizolepis) мы наблюдаем новое явление в эволюции шишки, а именно - срастание мегаспорофиллов со стерильными чешуями мегастробила в единый "пазушный комплекс". Исследование мегастробилов целого ряда других мезозойских хвойных показывает различные пути дальнейшего видоизменения и специализации мегастробилов.
Исследования Р. Флорина окончательно подтвердили "теорию пазушного брахибласта", которая первоначально носила скорее характер рабочей гипотезы, но теперь уже нет сомнений в том, что семенная чешуя есть брахибласт, гомологичный отдельному элементарному мегастробилу в мегаспорангиатной "сережке" кордаитовых. При этом у примитивных хвойных, так же как у кордаитовых, семязачатки были расположены на верхушках ножковидных мегаспорофиллов. Но у подавляющего большинства хвойных в результате возрастающей дорсивентрализации мегастробила и постепенного слияния всех его частей в одну чешуевидную структуру верхушечное расположение семязачатков переходит в поверхност-ное (ламинальное) - они оказываются на верхней (адаксиальной) стороне семенной чешуи. При занятии семязачатком поверхностного положения на семенной чешуе появляются тенденции к слиянию этой последней с кроющей чешуей. В восходящем ряду форм, ведущем от сосновых к таксодиевым и кипарисовым, происходит постепенное срастание семенной чешуи с кроющей. Конечным результатом этого эволюционного процесса является доминирование семенной чешуи и почти полное исчезновение кроющей чешуи. В некоторых случаях, как у родов атротаксис (Athrotaxis) и куннингамия (Cunninghamia) из семейства таксодиевых, доминирует кроющая, а не семенная чешуя, причем семенная чешуя редуцирована настолько, что ее проводящая система представлена немногими мелкими проводящими пучками, а иногда только одним пучком.
Исключительно интересны также те направления в эволюции элементов женской шишки, которые наблюдаются в семействах араукариевых и сосновых, но здесь мы можем коснуться этого вопроса только в нескольких словах. В семействе араукариевых наблюдаются все переходы от форм с чешуями шишки, имеющими явственно двойную природу, к формам с сильно упрощенными чешуями. Среди араукарий особенно выделяется явственно двойной природой своих шишечных чешуй араукария Бидвилла (Araucaria bidwillii). У рода агатис (Agathis) также встречаются виды, у которых чешуя имеет двойную серию проводящих пучков. В семействе подокарповых имеются постепенные переходы от видов подокарпа, имеющих свободные, не сросшиеся между собой кроющие и семенные чешуи, к простой, слитной чешуе родов саксеготея и микрокахрис.
Таким образом, в различных линиях эволюции хвойных происходит параллельный процесс постепенного срастания кроющей и семенной чешуи (точнее, чешуевидного мегастробила), что в конце концов приводит к образованию "простой", слитной "чешуи", которую некоторые ботаники предпочитают называть "фертильным комплексом".
Такова, в немногих словах, разгадка более чем вековой загадки семенной чешуи, достигнутая в результате кропотливых исследований нескольких поколений ботаников.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'