Класс гнетовые, или гнетодсиды (Gnetopsida) (А. Л. Тахтаджян)
"Путешествие в невозможное - я встречаю вельвичию" - так назвал свою статью английский ботаник Гордон Раули, посетивший в 1971 г. пустыни Юго-Западной Африки, где произрастает одно из наиболее замечательных растений на Земле - вельвичия удивительная (Welwitschia mirabilis). Волнение путешественника вполне понятно. Недаром знаменитый английский ботаник Джозеф Xукер, широко известный как один из сподвижников Чарлза Дарвина, назвал это впервые им описанное растение "самым удивительным" из всех известных ему растений (рис. 175). А португальский ботаник Ф. Вельвич, один из первых собравший это названное в его честь растение, говорил, что сначала он даже боялся дотронуться до него ... он опасался, что оно исчезнет.
В своей классической работе, опубликованной в 1863 г., Дж. Хукер дал не только исключительно тщательное и мастерское описание доставленного ему в Англию растения, собранного Вельвичем, но и попытался установить его родственные связи. На основании своих исследований он пришел к выводу, что вельвичия стоит ближе всего к двум другим, также очень своеобразным голосеменным - гнетуму (Gnetum) и эфедре (Ephedra). На первый взгляд этот вывод может показаться очень странным, так как все три рода очень сильно отличаются друг от друга. Если о вельвичии можно сказать, что она "ни на что не похожа", то гнетум напоминает скорее многие тропические двудольные древесные растения с цельными вечнозелеными кожистыми листьями, а эфедра - австралийкие двудольные древесные растения из рода казуарина (Casuarina) с их характерными чешуевидными листьями. Тем не менее последующими исследованиями добыто много интересных данных, доказывающих некоторую общность этих трех родов, хотя в то же время стало очевидным, что Хукер преувеличивал степень их близости. Что же общего между этими тремя "отщепенцами" растительного мира?
Несмотря на очень большое число весьма существенных различий, роды эфедра, вельвичия и гнетум имеют также ряд не менее важных общих признаков. Этими признаками являются: 1) совершенно необычное для современных голосеменных дихазиальное ветвление собраний стробилов; 2) наличие похожего на околоцветник покрова вокруг стробилов, что также совершенно необычно для современных голосеменных; 3) признаки прошлой обоеполости стробилов, особенно хорошо выраженные у вельвичии; 4) длинные микропилярные трубки, образованные вытянутым интегументом; 5) наличие сосудов во вторичной ксилеме, резко отличающее их от всех остальных голосеменных, как современных, так и вымерших.
Кроме того, можно упомянуть такие признаки, как общие черты в строении оболочки пыльцевых зерен, двусемядольные зародыши, супротивные листья, отсутствие смоляных ходов и пр.
Но родство трех родов не является столь близким, чтобы можно было объединять их в одно семейство гнетовых (Gnetaceae), как это делалось прежде. Каждый из них образует вполне самостоятельное семейство: эфедровые (Ephedraceae), вельвичиевые (Welwitschiaceae) и гнетовые (Gnetaceae). Более того, каждое семейство образует отдельный порядок (Ephedrales, Welwitschiales и Gnetales) и даже отдельный подкласс (Ephedridae, Welwitschiidae и Gnetidae).
Гнетопсиды, несомненно, представляют собой очень своеобразную и интересную группу растений, постоянно привлекающую внимание ботаников. Однако отсутствие палеоботанических данных лишает нас возможности выяснить их происхождение с той степенью достоверности, которая достигнута, например, для хвойных.
Семейство эфедровые (Ephedraceae)
В это семейство входит род эфедра, который иногда называют также хвойником или Кузьмичовой травой. Хвойником это растение называют потому, что виды эфедры несколько напоминают некоторые хвойные. Кузьмичовой же травой она была названа по имени народного лекаря Федора Кузьмича Муховникова из Самары, который популяризировал ее медицинское применение.
Род эфедра имеет более 40 видов и довольно широко распространен в областях с сухим климатом, главным образом в Средиземноморье, в Азии, западной части Северной Америки и в Южной Америке.
Виды эфедры представляют собой более или менее ксерофильные и полуксерофильные растения, произрастающие в пустынях, полупустынях, степях и редколесьях, на песчаных и скалистых местообитаниях. В большинстве случаев это низкие, сильно ветвистые кустарники, реже они достигают высоты 6-8 м и являются прямостоячими или вьющимися кустарниками, иногда приобретают древовидный облик.
Один из южноамериканских видов - эфедра трехтычиночная (Ephedra triandra) - представляет собой небольшое дерево. Внешне эфедра напоминает хвощи и своеобразные австралийские покрытосеменные - казуарины.
Проводящая система осевых органов эфедры представляет собой типичную эндархную эвстелу, очень сходную во многих отношениях с проводящей системой хвойных и гинкго. Вся метаксилема и большая часть вторичной ксилемы состоит их трахеид с округлыми окаймленными порами, снабженными типичным торусом. Но у эфедры, в отличие от хвойных и гинкго, во вторичной ксилеме наряду с трахеидами имеются также настоящие сосуды. Однако происхождение сосудов у эфедры, как и вообще у гнетопсид, иное, чем у покрытосеменных. Сосуды у нее возникли в результате расширения округлых окаймленных пор на косых конечных стенках и утери как торусов, так и окаймлений. Это привело к возникновению сосудов с многочисленными мелкими, овальными или округлыми сквозными отверстиями. Такие сквозные отверстия иногда сливаются и образуют перфорацию, часто очень похожую на простую перфорацию сосудов многих покрытосеменных.
Молодые ребристые стебли зеленые и несут, следовательно, функцию фотосинтеза. Эпидермальные клетки толстостенные, а устьица расположены в бороздах между ребрами. Ребра образованы группами очень толстостенных клеток. Между ними и эпидермой, с одной стороны, и стелой - с другой, расположена зона очень тонкостенных клеток с обильными хлоропластами и многочисленными межклеточными пространствами.
Листья эфедры большей частью рано опадающие, мелкие, обычно чешуевидные, редко более длинные и нитевидные (длиной до 1 см), супротивные или в мутовках по 3, редко по 4. Листовые следы двойные. Устьиц на листьях очень мало.
Стробилы эфедры однополые и почти всегда двудомные (рис. 174). Редко встречаются однодомные экземпляры. У некоторых видов найдены также аномальные обоеполые собрания стробилов и даже атавистические обоеполые стробилы.
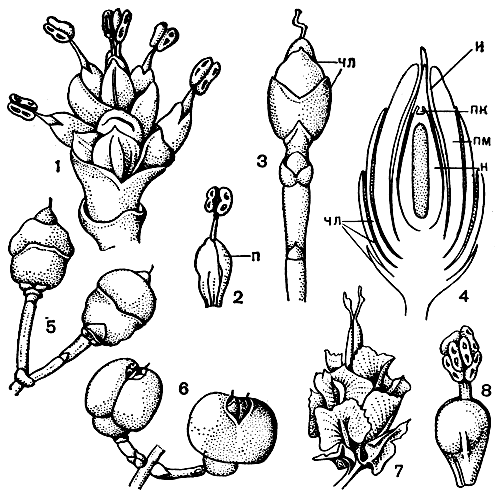
Рис. 174. Стробилы эфедры. Эфедра высочайшая (Ephedra altissima): 1- собрание микростробилов; 2 - отдельный микростробил (п - покров микростробила); 3 - собрание мегастробилов, сведенное к одному мегастробилу (чл - стерильные чешуевидные листья); 4 - продольный разрез верхней части предыдущей фигуры (чл - стерильные чешуевидные листья, н - нуцеллус, пк - пыльцевая камера, и - интегумент, вытянутый наверху в микропилярную трубку, пм - покров мегастробила); 5 - зрелые семена. Эфедра согнутоножковая (Е. campylopoda): 6 - зрелые семена. Эфедра крылатая (Е. alata): 7 - зрелые семена. Эфедра ломкая (Е. fragilis): 8 - отдельный микростробил
Собрания микростробилов сидят супротивно или мутовчато, по 3 или редко по 4, в пазухах обыкновенных листьев на узлах мелких зеленых ветвей и часто дихазиально ветвятся. Каждое такое собрание микростробилов состоит из короткой оси, несущей от 2 до 8 супротивных пар чешуевидных кроющих листьев, из которых одна или две нижние стерильны, а в пазухах остальных сидит по одному очень просто устроенному микростробилу. Весь микростробил состоит из своеобразного покрова, называемого часто "околоцветником", и одного "антерофора". Покров образован двумя тонкими супротивными чешуевидными листьями, расположенными медианно (в заднепереднем плане) и сросшимися основаниями. Что же касается антерофора, то он представляет собой центральную колонку, несущую на верхушке от 2 до 8 микросинангиев. Микросинангии двугнездные или иногда трехгнездные и даже четырехгнездные. Иногда антерофор бывает раздвоен или разделен до основания. В антерофор входят два самостоятельных проводящих пучка. Антерофор эфедры произошел в результате срастания двух микроспорофиллов.
Гнезда микросинангиев открываются овальными дырочками. Пыльцевые зерна эллипсоидальные, с продольными складками, обычно безбороздные, но у эфедры хвощовой (Ephedra equisetina) наблюдается остаточная борозда.
Собрания мегастробилов сидят, как и собрания микростробилов, по 2, 3 или 4 в пазухах листьев, в узлах мелких зеленых веточек. Каждое из них состоит из короткой пазушной оси, нескольких (обычно 4 или больше) пар стерильных чешуевидных листьев и обычно одного, реже 2-3 очень редуцированных мегастробилов. Одиночные мегастробилы кажутся верхушечными, но в действительности они сидят в пазухе кроющей чешуи.
Каждый мегастробил состоит из одного семя зачатка, окруженного особым толстым и мясистым мешочкообразным покровом ("околоцветником"), неправильно называемым часто внешним интегументом, Покров этот соответствует покрову микростробила, но срастание у него более полное. Интегумент семязачатка (называемый часто внутренним интегументом) вытягивается в более или менее длинную микропилярную трубку. На верхушке микропилярной трубки ко времени опыления появляется капелька жидкости.
При прорастании микроспоры отделяется сначала первая проталлиальная клетка. При втором делении образуется ядро второй проталлиальной клетки, но самой клетки не образуется, так как ядро это не отделяется перегородкой от ядра антеридиальной клетки. Далее ядро антеридиальной клетки делится и образует ядро генеративной клетки и трубочковой клетки. Эти два ядра лежат в общей массе цитоплазмы и никогда не отделяются перегородкой. Ядро генеративной клетки делится и образует ядро клетки-ножки и ядро сперматогенной клетки. Последнее дает начало двум спермиям.
В мегаспорангии эфедры из мегаспоры в результате свободного ядерного деления и последующего образования клеточных перегородок развивается массивный женский гаметофит. В каждом из них закладываются обычно два архегония. Архегоний имеет длинную шейку, состоящую из 32 или более клеток. При делении ядра центральной клетки архегония между брюшным канальцевым ядром и ядром яйцеклетки не образуется клеточной перегородки.
Пыльцевые зерна переносятся на семязачатки при помощи ветра, хотя для некоторых видов не исключено участие насекомых. Микропилярная трубка заполнена сахаристой жидкостью, которая образуется в результате разрушения тканей верхушки мегаспорангия. Жидкость эта выступает из микропилярной трубки в виде капельки. Пыльцевые зерна попадают на выступающую капельку жидкости и по мере усыхания жидкости в микропилярной
трубке втягиваются внутрь,- доходя до верхушки мегаспорангия. В результате образования воронкообразной пыльцевой камеры пыльцевые зерна попадают прямо на женский гаметофит. Поэтому образуется лишь короткая пыльцевая трубка, которая через шейку архегония достигает яйцеклетки. Таким образом, у эфедровых так же, как у хвойных и цгетковых растений, образуется пыльцевая трубка, а не гаустория. К моменту внедрения пыльцевой трубки в женский гаметофит ядро клетки-ножки и ядро трубочковой клетки исчезают. Один из двух спермиев сливается с яйцеклеткой и дает начало зародышу.
При созревании семени верхние 4 чешуевидных кроющих листа мегастробилов делаются обычно сочными и приобретают красную, оранжевую или желтую окраску, представляя собой, таким образом, зоохорное приспособление. У некоторых видов, однако, эти чешуевидные кроющие листья остаются сухими, иногда деревенеют и становятся более или менее крылатыми. Покров же ("околоцветник") мегастробила становится при семенах деревянистым или реже сочным и окружает семя. Интегумент остается перепончатым.
Уже давно было замечено лекарственное значение некоторых видов эфедры. Широко распространенные в Средней Азии, эфедра хвощовая и некоторые другие виды вошли в официальную медицину как важное сердечное и противоастматическое средство (алкалоид эфедрин). Из "ягод" ("степная малина") иногда готовят варенье, по вкусу напоминающее мед.
Семейство вельвичиевые (Welwitschiaceae)
Единственный представитель семейства - вельвичия удивительная (Welwitschia mirabilis), растет в бесплодных каменистых пустынях Анголы и Юго-Западной тропической Африки, главным образом в прибрежной пустыне Намиб, заходя в глубь материка обычно не далее 100 км. Пустыня Намиб отличается крайней сухостью и знойностью. Целыми месяцами здесь не выпадает ни капли дождя, а в некоторых местах годовое количество осадков не превышает 25 мм. Вельвичия встречается на каменистых равнинах или в сухих руслах временных потоков, там, где стояние грунтовых вод относительно близко. Она является крайне выраженным ксерофитом, приспособленным к условиям резко пустынного климата (карта 11).
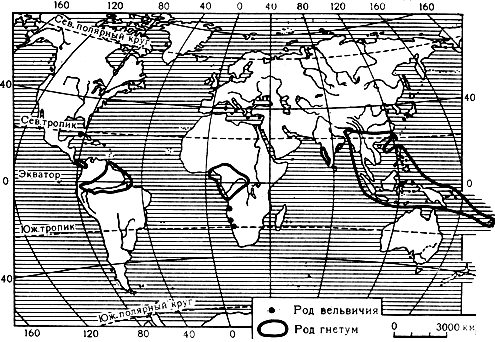
Карта 11. Ареалы родов вёльвичия и гнетум
Внешний вид и строение вельвичии весьма своеобразны (рис. 175). Если эфедру мы могли сравнить по внешнему облику с хвойным или казуариной, то вельвичия не похожа ни на одно из известных нам растений. "Не дерево, не куст, не трава, а нечто совершенно своеобразное", - писал об этом растении в 1922 г. Б. М. Козо-Полянский. Она представляет собой своеобразное дерево-карлик, настоящий "монстр" среди растений. Вельвичия имеет не очень длинный (не более 3 м) главный корень. Ствол у нее похож на обрубок или пень, очень низкий и толстый (диаметром до 1,2 м), почти полностью скрытый в земле - надземная часть редко превышает в высоту 50 см. Большая часть этого ствола представляет собой подсемядольное колено. Книзу ствол вельвичии конусообразно суживается, а наверху он более или менее седловидно-двулопастный и по краям лопастей несет по одному гигантскому листу. На этих двух лопастях заметны ряды концентрических гребней, соответствующих сезонам роста. Два супротивных кожистых листа вельвичии остаются на всю жизнь растения, которая длится столетия, а у некоторых экземпляров 2000 лет и, возможно, даже больше. Листья имеют неограниченный рост при основании (посредством интеркалярной меристемы), растут со скоростью 8-15 см в год и достигают в длину 2 и даже иногда 3 м. Описан гигантский экземпляр с листьями длиной 6,2 м при ширине 1,8 м. Интересно, что живая, фотосинтезирующая его часть достигала в длину 3,7 м, а общая фотосинтезирующая поверхность имела площадь 21 м2. В своей статье "Welwitschia mirabilis - парадокс пустыни Намиб" (1972) американский ботаник Крис Борнман пишет, что он видел вельвичию, у которой лист достигал длины 8,8 м! Листья вельвичии имеют параллельное жилкование. У взрослого растения листья разрываются на длинные ленты, постепенно отмирающие на своих концах. Соседние экземпляры иногда срастаются друг с другом.
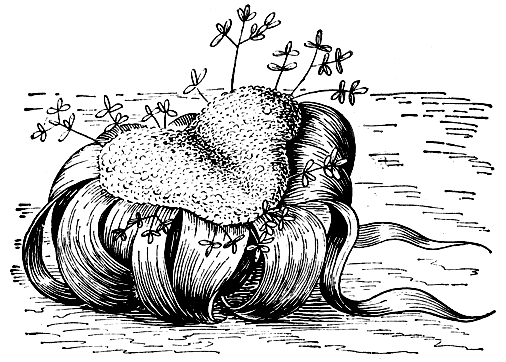
Рис. 175. Вельвичии удивительная (Welwitschia mirabilis)
Как по своей внешней морфологии, так и по анатомическому строению стебля вёльвичия является как бы "взрослым проростком". Надсемядольная часть вельвичии недоразвита, и почти весь клубневидный ствол ее морфологически соответствует эпикотилю, который несет лишь два первых листа. Вегетативная часть спорофита вельвичии представляет собой фиксированную и сильно видоизмененную юношескую стадию развития древовидного предкового типа. Но если вёльвичия - "взрослый проросток", то проросток, очень сильно видоизмененный и всем своим строением прекрасно приспособленный к пустынному климату.
Ствол вельвичии покрыт толстым слоем перидермы, достигающим толщины 2 см. Проводящая система эндархная. Расположение проводящих пучков в стволе неправильное, но в молодых стеблях и особенно в корнях наблюдается концентрическое расположение и ясно видны кольца прироста, которые не являются, однако, годичными. Каждое кольцо имеет свою ксилему и флоэму, как у поликамбиальных саговниковых. В ножках собраний стробилов расположение проводящих пучков более правильное, и на поперечном разрезе у них обычно видны ряды ксилемы толщиной в одну клетку, с тонкостенной паренхимой между ними. Во вторичной древесине трахеиды имеют округлые окаймленные поры, часто с сетчатыми утолщениями между порами. Наряду с трахеидами у вельвичии встречаются настоящие сосуды с простой, но неполной перфорацией на слегка косых конечных стенках и округлыми окаймленными порами на боковых стенках. Флоэма вельвичии состоит из очень своеобразных ситовидных элементов; ядро вместе со своей оболочкой сохраняется даже в зрелых ситовидных клетках, что очень необычно. В различных частях растения имеются слизевые ходы, как у саговниковых. Листовые следы двойные. Листья имеют резко выраженное ксероморфное строение. Устьица синдетохейльные (сложногубые), как у беннеттитовых.
Откуда же черпает вёльвичия необходимую ей влагу в этой практически безводной пустыне? Почти единственным источником влаги является здесь густой туман, который окутывает побережье в течение около 300 дней в году. Восточные ветры часто гонят туман вглубь до 80 км. Конденсированный туман равноценен 50 мм осадков. Конденсируясь на огромных листьях вельвичии, влага поглощается через устьица и поступает в проводящую систему растения. Поэтому неудивительно, что вёльвичия обладает исключительно большим числом устьиц на обеих сторонах листа (22 200 устьиц на 1 см2!). Устьица погруженные, но не столь глубоко, как обычно у ксерофитов. Другими словами, вёльвичия - ксерофит, но ксерофит очень своеобразный, сильно отличающийся от тех ксерофитов, которые знакомы нам, например, по пустыням Средней Азии.
Микро- и мегастробилы вельвичии образуют сложные ветвистые собрания, развивающиеся на разных особях (рис. 176). Многочисленные собрания стробилов возникают непосредственно над основаниями листьев, как бы в их пазухах.
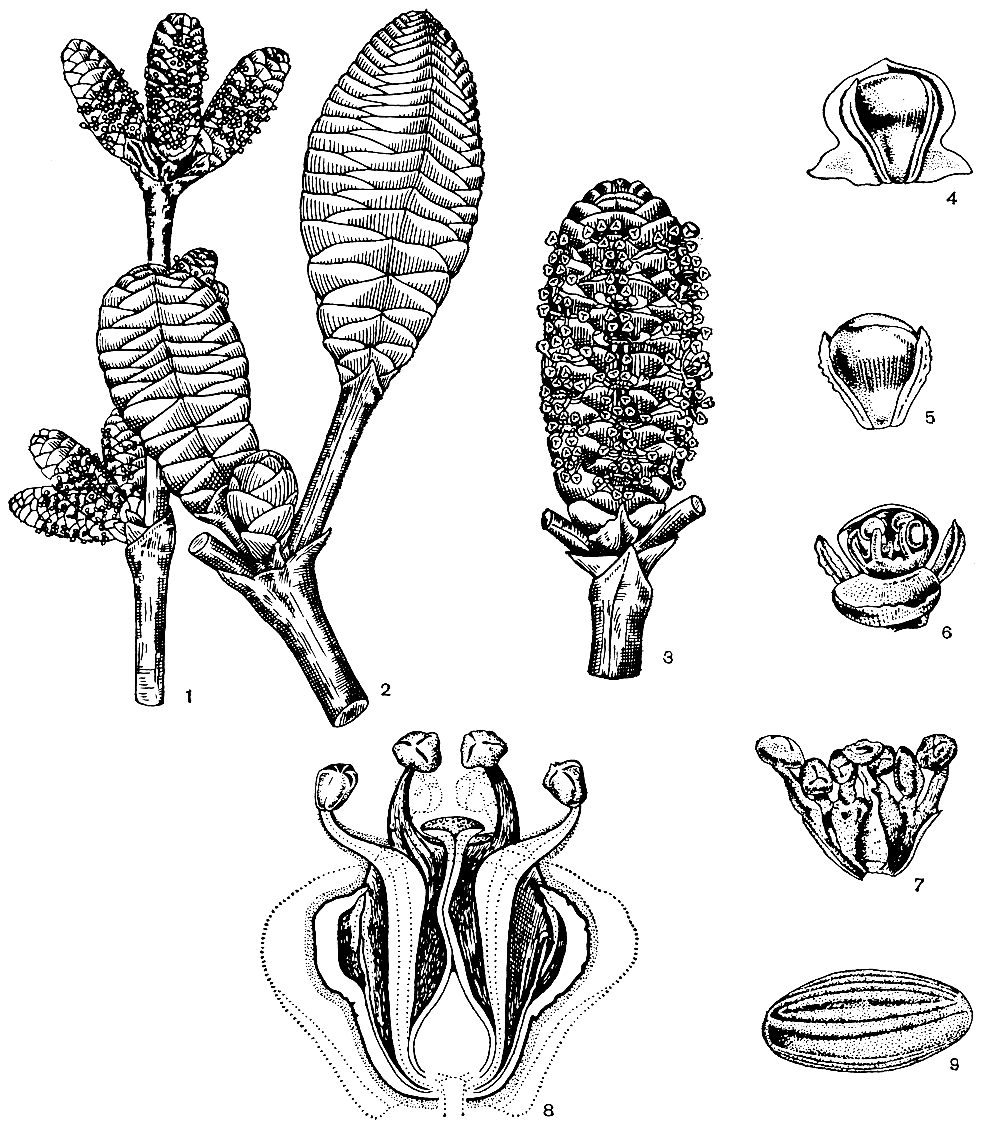
Рис. 176. Вельвичия удивительная (Welwitschia mirabilis). Стробилы: 1 - собрание микростробилов; 2 - часть собрания мегастробилов; 3 - часть собрания микростробилов. Микростробил: 4 в пазухе кроющей чешуи; 5 - без кроющей чешуи; 6 - с раскрытым покровом; 7 - с удаленным покровом (внутри виден рудиментарный семязачаток); 8 - в продольном разрезе. Микроспора - 9
Собрания микростробилов ветвятся дихазиально. Каждая отдельная веточка похожа на шишку, состоящую из крест-накрест расположенных супротивных кроющих чешуй. В пазухе каждой кроющей чешуи, за исключением лишь самых нижних, сидит по одному микростробилу.
Микростробил состоит из двух пар супротивных покроволистиков, расположенных циклически и сросшихся основаниями микроспорофиллов и находящегося в центре рудиментарного семязачатка. Два внешних боковых покроволистика мелкие и с острым килем, а два внутренних (медианных) - более крупные, расположенные параллельно большой кроющей чешуе. Два боковых микроспорофилла, находящиеся против боковых покроволистиков, обычно более крупные, чем остальные четыре, расположенные по два против каждого из двух медианных покроволистиков. Каждый микроспорофилл снабжается одним проводящим пучком. Основания микроспорофиллов срастаются в короткую трубку. На верхушке микроспорофиллы несут по одному трехгнездному микросинангию, каждое гнездо которого открывается одной радиальной трещиной. Самой замечательной особенностью микростробилов вельвичии является рудиментарный семязачаток, свидетельствующий об обоеполости стробилов предков вельвичии. Некоторые авторы предполагают, что этот рудиментарный семязачаток исполняет роль нектарника (пример смены функции в процессе эволюции).
В микроспорангиях развиваются многочисленные эллипсоидальные микроспоры. Пыльцевые зерна эллипсоидальные, однобороздные, с продольно-мелкоскладчатой поверхностью. Как указывают московские палинологи В. В. Бернард и Н. Р. Мейер (1972), в строении оболочки пыльцевых зерен эфедры и вельвичии имеются такие важные общие признаки, как эллипсоидальная форма зерен, однослойная складчатая эктэкзина, имеющая много мелких отверстий, расположенных между гребнями, плотная эндэкзина и пр.
Дихазиально ветвистые собрания мегастробилов гораздо крупнее, чем собрания микростробилов. Отдельные шишкообразные веточки таких собраний состоят из расположенных крест-накрест супротивных пар кроющих чешуй, сперва зеленых, позже ярко-красных. В отличие от микростробила мегастробил состоит лишь из одной пары покроволистиков, окружающих единственный семязачаток; медианные покроволистики у них отсутствуют и сохранились лишь боковые из них, но они крупнее, чем в микростробилах, и дают большие крыловидные выросты. Интегумент семязачатка вытягивается в очень длинную микропилярную трубку, которая, однако, не расширяется на верхушке грибообразно, как у рудиментарных семязачатков в микростробилах.
При прорастании микроспоры сначала образуются ядра проталлиальной и антеридиальной клеток, которые не разделяются клеточной перегородкой. Антеридиальное ядро делится и дает начало большому ядру трубочной клетки и генеративному ядру. В таком трехъядерном состоянии пыльцевые зерна переносятся ветром на семязачаток. Предполагалось, что вельвичию опыляют также насекомые. Находки на некоторых растениях перепончатокрылых, казалось, подтверждали это. Но эти насекомые оказались просто паразитами, которые питаются молодыми семязачатками и сочной флоэмой мегастробила. Более того, эти насекомые с рудиментарными крыльями вообще не могут летать.
Пыльцевые зерна попадают на капельку сахаристой жидкости, выступающей из микропилярной трубки. По мере усыхания этой жидкости, находясь еще в трехъядерной стадии, они спускаются вниз по длинной трубке и достигают нуцеллуса. Пыльцевая трубка может начать развиваться даже еще в микропиле. Генеративное ядро вскоре после проникновения пыльцевой трубки в нуцеллярный конус делится, образуя два ядра спермиев, которые лежат в общей массе цитоплазмы. Таким образом, развитие мужского гаметофита максимально приближается к покрытосеменным и отличается лишь развитием ядра проталлиальной клетки. Оба ядра спермиев являются потенциально функционирующими, но одно из них крупнее и расположено впереди. Часто, однако, одно из ядер разрушается и исчезает. Ядро трубки к этому времени исчезает.
В мегаспорангии в результате свободного ядерного деления возникает крайне своеобразный женский гаметофит. Самой замечательной особенностью его является полное отсутствие архегониев. Отсутствует также большая центральная вакуоль, и многочисленные (около 1024) свободные сначала ядра равномерно распределены по всей массе гаметофита. Далее женский гаметофит начинает ясно дифференцироваться на верхнюю фертильную и нижнюю большей частью стерильную часть. Фертильная часть охватывает приблизительно микропилярную четверть гаметофита. В обеих частях гаметофита образуются клеточные перегородки, отделяющие несколько, часто до дюжины и даже больше, ядер в одно отделение в нижней части, в то время как в верхней части в каждом отделении имеются обычно 2 или 3 ядра, реже больше. Клетки в верхней части крупнее, чем в нижней. Ядра в каждой клетке постепенно сливаются в одно, причем этот процесс завершается раньше в микропилярной части. Ядра в клетках микропилярной части увеличиваются постепенно в размерах, и цитоплазма вакуолизируется. По мере увеличения вакуоли, цитоплазма с содержащимися в ней ядрами перемещается к одной из стенок клетки. Здесь часть стенки начинает выпячиваться в форме трубки, в которую переходят цитоплазма и ядро. Эти так называемые проталлиальные трубки удлиняются, растут вверх, переходят через оболочку мегаспоры и врастают в стенку мегаспорангия. Вся масса цитоплазмы вместе с ядрами передвигается к кончику трубки. Редко все ядра сливаются вместе. Многие авторы считают проталлиальные трубки многоядерными яйцеклетками.
Обычно на полпути проталлиальные трубки встречаются с пыльцевыми трубками. Эта встреча происходит близ центра нуцеллярного конуса. Проталлиальная и пыльцевая трубки сталкиваются своими концами, стенки их в местах соприкосновения разрушаются, одно из передних ядер проталлиальной трубки входит в цитоплазму пыльцевой трубки и сливается с одним из спермиев. Другими словами, происходит нечто противоположное тому, что наблюдается у остальных семенных растений, где спермий входит в яйцеклетку, а не наоборот, как у вельвичии. После оплодотворения ядра зигота на своем пути в женский гаметофит переходит из пыльцевой трубки в проталлиальную.
При семенах покров стробила становится крылатым (анемохорное приспособление). Проросток вельвичии имеет две семядоли и два листа. Семядоли сохраняются два или три года, а затем опадают. В пазухе каждой семядоли появляется почка, которая уплощается и расширяется, пока обе почки не встретятся, образуя сплошной покров, под которым скрывается остановившаяся в росте верхушка стебля.
Вельвичия иногда культивируется в оранжереях, что, однако, требует со стороны садоводов большого искусства и умения.
Семейство гнетовые (Gnetaceae)
В семейство входит один род гнетум (Gnetum), заключающий около 30 видов, распространенных во влажных тропических странах, главным образом в Азии и в Малезии, а также в северной части Южной Америки и тропической Западной Африке; два вида произрастают в Африке (Камерун и Ангола). Нет ни одного общего вида между западным и восточным полушариями (карта 11).
Если эфедра и вельвичия являются типичными ксерофитами, то виды гнетума, наоборот, обитают во влажных тропических лесах. В большинстве случаев это лианы, вьющиеся, лазящие или цепляющиеся, достигающие часто вершин высочайших деревьев. Лишь два вида - гнетум гнемон (Gnetum gnemon, рис. 177), распространенный от Ассама до Фиджи, и гнетум ребристый (G. costatum), встречающийся на Соломоновых островах и в Новой Гвинее, - являются прямостоячими деревьями (хотя гнетум гнемон иногда становится лианой, как это неоднократно замечалось в Новой Гвинее). Некоторые виды являются кустарниками.
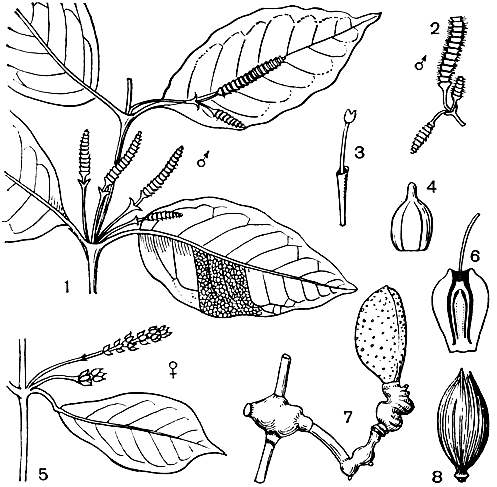
Рис. 177. Гнетум гнемоновидный (Gnetum gnemonoides): 1 - часть ветви с собраниями микростробилов; 2 - собрание микростробилов; 3 - микростробил; 4 - стерильный мегастробил; 5 - собрание мегастробилов; 6 - мегастробил в продольном разрезе; 7 - ветвь с семенем; 8 - семя без внешнего слоя
Для рода гнетум характерны большие, широкие, цельные, кожистые, перистонервные листья с типичным сетчатым жилкованием, очень похожие на листья многих тропических двудольных растений (рис. 178). Листья все супротивные и перекрестнопарные, сидящие на коротких черешках. По краям листьев гнетума гнемона иногда образуются выводковые почки, как у бриофиллума (Bryophyllum). Стебли членистые и часто вздутые у узлов.

Рис. 178. Лист гнетума гнемон (Gnetum gnemon)
Проводящая система стебля представляет собой эндархную эвстелу, с хорошо развитыми пучками, широкими лучами, толстой корой и многочисленными слезевыми ходами. Протоксилема состоит из трахеид со спиральными утолщениями; вторичная ксилема - из трахеид с многорядными округлыми окаймленными порами и из более широких (в 4-5 раз) сосудов с боковой поровостью, как у трахеид. Простая перфорация у сосудов образуется в результате расширения или слияния окаймленных пор и исчезновения замыкающих пленок. Они напоминают сосуды очень подвинутых покрытосеменных, но образовались совершенно иным путем. У некоторых видов встречаются клетки, аналогичные клеткам-спутницам покрытосеменных, но возникающие также иным путем. Они возникают не из одной и той же материнской клетки, как у покрытосеменных, но образуются независимо из камбия. Таким образом, хотя функционально они и аналогичны клеткам-спутницам покрытосеменных, но по происхождению резко отличны. Строение стебля обычного монокамбиального типа со сплошным цилиндром древесины и постоянно действующим камбием. У лиановых форм стебли имеют поликамбиальное строение.
Как и двум другим родам, гнетуму присуща двудомность. Собрания стробилов у гнетума или простые в виде колосков, или у их основания возникают вторичные боковые веточки, которые могут, в свою очередь, ветвиться (рис. 177). Тонкая ось каждого колоска несет попарно чашеобразно сросшиеся супротивные кроющие чешуи, в пазухах которых вперемежку с многочисленными волосками сидят стробилы.
В собраниях микростробилов междоузлия у большинства видов короткие, и кроющие чешуи сближены. Вокруг оси в пазухах чашеобразно сросшихся кроющих чешуй кольцеобразно располагаются несколько рядов микростробилов. Эта многорядная мутовка возникла, вероятно, в результате сильного укорочения и видоизменения двух пазушных веточек.
Микростробил состоит из трубчатого и наверху двулопастного покрова, окружающего один-единственный микроспорофилл. Покров состоит из двух сросшихся покроволистиков, как в микростробиле. Микроспорофилл гнетума, в отличие от микроспорофилла эфедры, всегда цельный и несет на верхушке обычно 2 боковых микроспорангия, которые иногда более или менее срастаются между собой.
Пыльцевые зерна гнетума шарообразные, безбороздные или однобороздные, мелкошиповатые, без мелких отверстий на поверхности (табл. 45). Внешне они сильно отличаются от пыльцевых зерен эфедры и вельвичии, но, как показал японский палинолог Й. Уэно (1960), ультраструктура пыльцевых зерен всех трех родов во многом сходна.
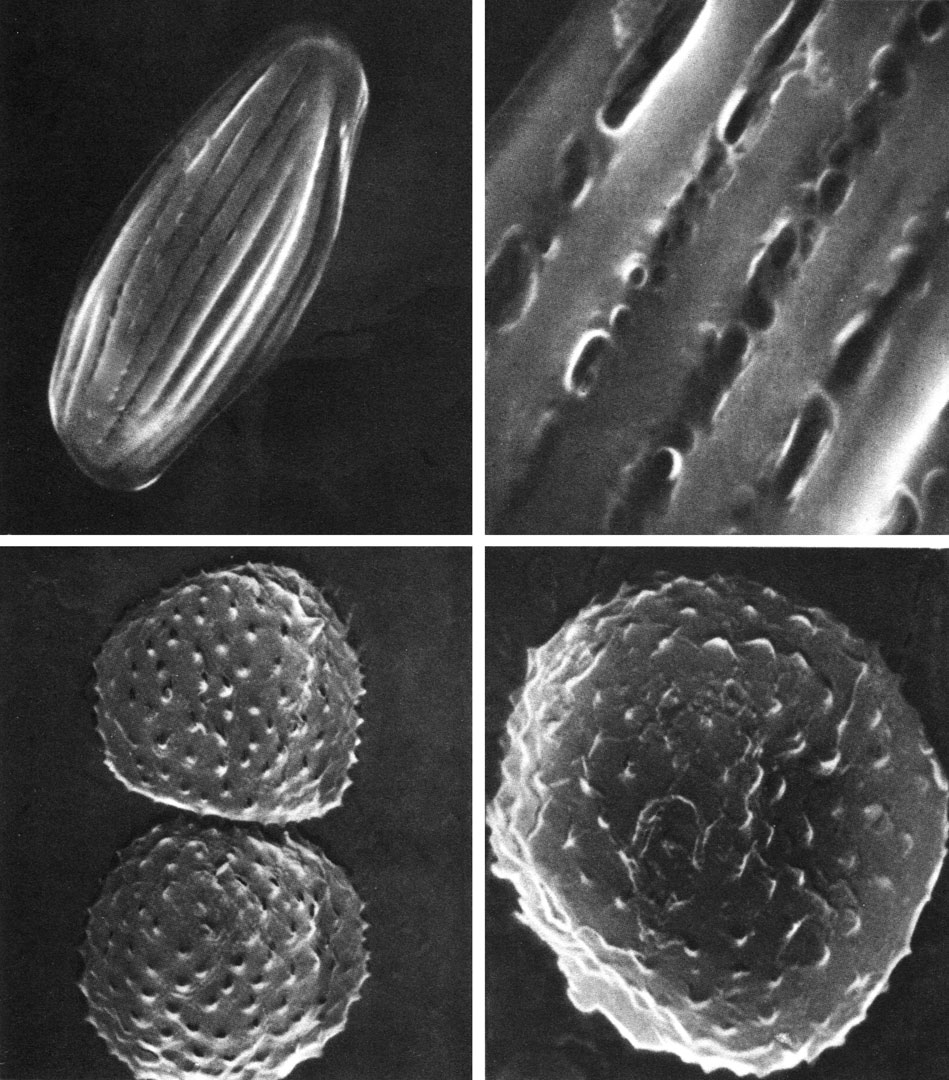
Таблица 45. Пыльцевые зерна растений из класса гнетовых под сканирующим электронным микроскопом: вверху - вельвичия удивительная (Welwitschia mirabilis): слева - увел, около 1500, справа - участок экзины, увел, около 10 000; внизу - гнетум гнемон (Gnetum gnemon); слева - увел, около 5000, справа - увел, около 8000
Собрание мегастробилов также состоит из главной оси и сросшихся основаниями супротивных листьев, но междоузлия у них длиннее, чем в собраниях микростробилов, мутовки мегастробилов в пазухах чешуй однорядные и число мегастробилов в каждой мутовке значительно меньше (3-8). Каждый мегастробил состоит из одного семязачатка и двух покровов, внутренний из которых часто считается внешним интегументом. Наружный покров очень толстый, снабженный многими проводящими пучками. Он состоит из двух сросшихся покроволистиков. Внутренний покров также снабжен проводящими пучками. Так же как у эфедры и вельвичии, интегумент семязачатка вытягивается наверху в характерную микропилярную трубку. В зрелом семени он превращается в тонкий бумажистый слой. Внутренний покров мегастробила образует твердый, каменистый слой, а внешний покров становится мясистым и ярко окрашенным. Зрелое семя имеет поэтому трехслойный покров, с каменистым слоем посредине, напоминая внешние семена саговниковых. В мегаспорангии обычно две или три материнские клетки мегаспор образуют по 4 мегаспоры. Из них некоторые прорастают.
При прорастании микроспоры содержимое ее делится на маленькую проталлиальную клетку и большую антеридиальную клетку. Ядро антеридиальной клетки делится на 2 ядра, одно из которых представляет собой ядро трубки, а второе - ядро генеративной клетки. Вокруг ядра генеративной клетки имеется слой цитоплазмы, и мы можем поэтому говорить о генеративной клетке. Ядро трубки первым входит в пыльцевую трубку, а за ним следует генеративная клетка. Проталлиальная клетка постепенно исчезает. Таким образом, зрелое пыльцевое зерно состоит из проталлиальной клетки, генеративной клетки и ядра трубки. При прорастании пыльцевого зерна генеративная клетка входит вслед за ядром трубки в пыльцевую трубку и делится при этом на две неравные мужские клетки (или ядра), меньшая из которых, по-видимому, дегенерирует. Микроспоры переносятся, вероятно, насекомыми на капельку жидкости, выделяемой микропилярной трубкой. Здесь они втягиваются высыхающей жидкостью до мегаспорангия, причем пыльцевая трубка образуется часто еще в микропилярной трубке.
Прорастание мегаспоры начинается, как и у всех остальных голосеменных, с последовательного ряда свободных ядерных делений, но, однако у гнетума свободноядерное состояние сохраняется в микропилярной части гаметофита до момента оплодотворения. Лишь в нижней части гаметофита гнетума образуется клеточная ткань, которая развивается из свободноядерного состояния, как у вельвичии, с неправильным образованием перегородок и несколькими вначале ядрами в каждой клетке. Всего у гнетума образуется 256 или реже 512 ядер.
Дальнейшие изменения начинаются после соприкосновения пыльцевой трубки с женским гаметофитом. В это время одно, два или три ядра в верхней части женского гаметофита обособляются от остальных ядер, вокруг них формируется слой цитоплазмы, и они становятся яйцеклетками. Пыльцевые трубки, прорастая через нуцёллус, достигают гаметофита, и оба спермия, ядро трубки и некоторое количество цитоплазмы изливаются в женский гаметофит.
Один из спермиев сливается с яйцеклеткой. После оплодотворения верхняя часть женского гаметофита также приобретает клеточное строение.
В развитии зародыша, так же как у вельвичии, нет свободной ядерной стадии. Развитый зародыш имеет сильно развитую ножку (гаусторию), как у вельвичии. Зародыш имеет длинный гипокотиль и две семядоли.
Ярко-розовые семена гнетума распространяются птицами, но некоторые виды (например, гнетум гнемоновидный - G. gnemonoides с крупными семенами с пробковым слоем) распространяются, вероятно, водой.
В некоторых тропических странах виды гнетума имеют разнообразное применение. Так, внутренняя кора ряда видов (гнетум гнемон, гнетум широколистный - G. latifolium и др.) очень ценится в Юго-Восточной Азии из-за прочного волокна, из которого изготовляют крученые нитки, веревки и снасти. Из обработанных волокон получают бумагу. Гнетум культивируется в Юго-Восточной Азии как "плодовое" дерево, молодые листья, стробилы и зародыши которого употребляют в пищу. Гнетум ула (G. ula) является источником съедобного масла, которое применяется также для массажа (предполагается антиревматическое действие).
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'