Класс беннеттитовые, или беннеттитопсиды (Bennettitopsida)
Порядок беннеттитовые (Bennettitales)
Беннеттитовые - одна из наиболее известных групп голосеменных растений мезозойской эры. Они начали свое существование с триаса, а по некоторым данным - с перми, т. е. с конца палеозойской эры. Расцвет группы приходился на середину мезозоя - юру и ранний мел. В середине мелового периода количество беннеттитовых резко сократилось и к концу мела (около 70 млн. лет назад) они окончательно вымерли. Таким образом, все, что мы знаем о беннеттитовых, основано исключительно на ископаемых остатках (табл. 44).
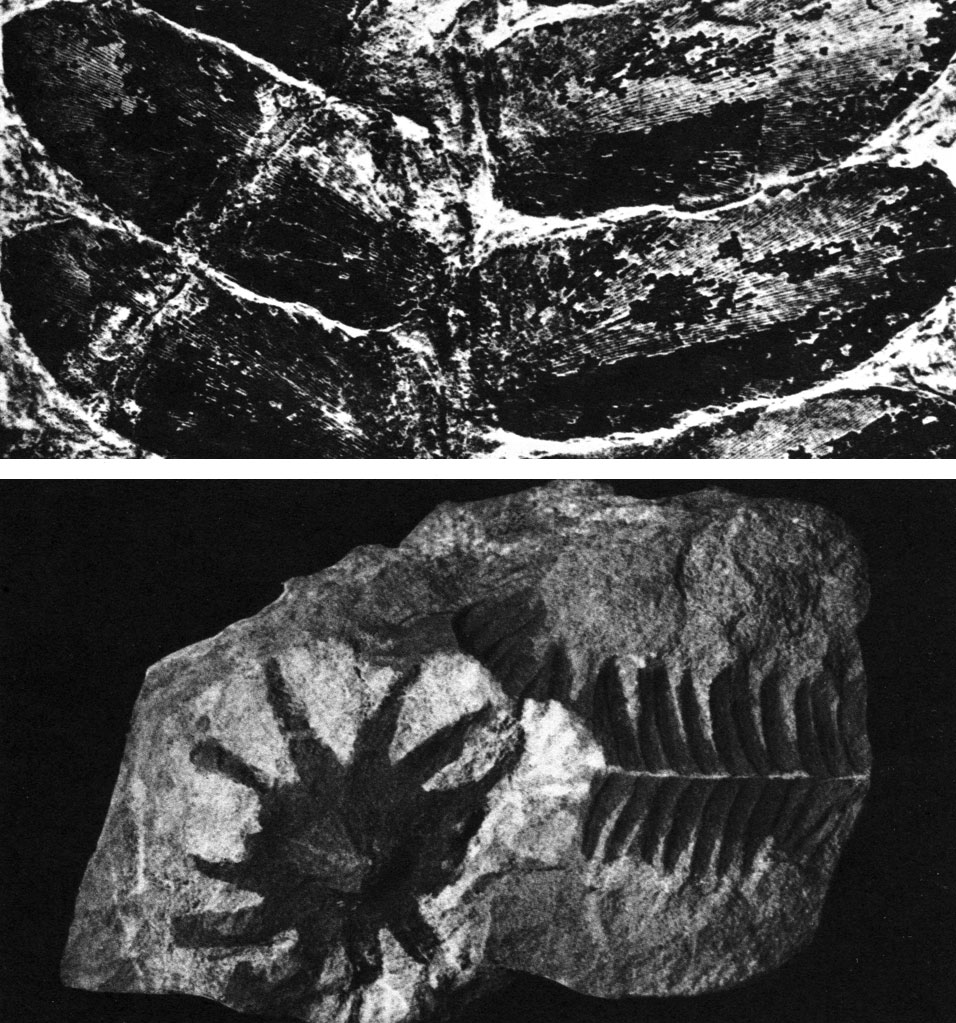
Таблица 44. Беннеттитовые: вверху - участок крупного листа отозамитеса туркестанского (Otozamites turkestanicus) из юрских отложений Казахстана (неск. увел.); внизу - отпечатки микростробила вельтрихии (Weltrichia whitbiensis) и листа птилофиллума (Ptilophyllum acutifolium), рассматриваемые как части одного растения (нат. вел., юра, Ткварчельский угленосный бассейн в Закавказье)
По внешнему виду и характеру вегетативных органов беннеттитовые, как уже отмечалось, были очень похожи на вымершие и современные саговниковые, (рис. 170), и именно к последним еще в начале XX в. отнесли большинство находок беннеттитовых.
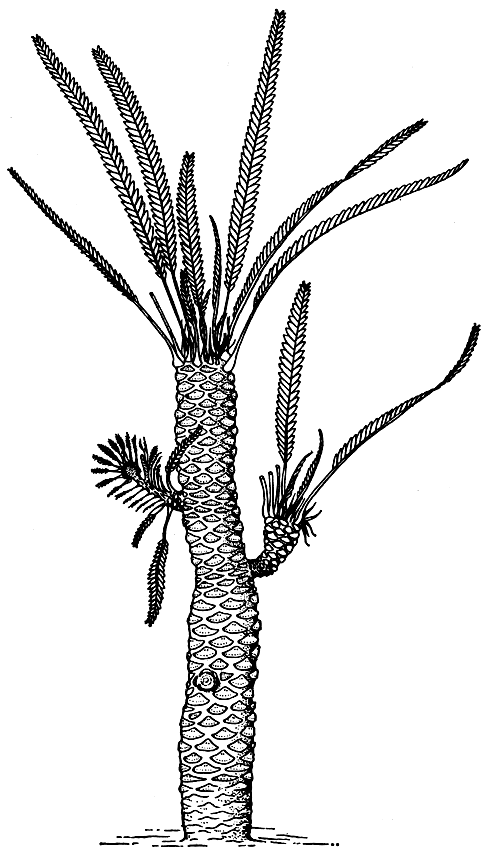
Рис. 170. Вильямсония Сьюорда (Williamsonia sewardiana). Реконструкция целого растения
Стебли (стволы) большинства беннеттитовых, как и у саговниковых, были простыми или слабоветвистыми, а по высоте приземистыми, толстыми и более или менее клубневидными, или же высокими и тонкими. Значительную часть стебля занимала сильно развитая сердцевина, часто со смоляными каналами, окруженная сравнительно узким кольцом . проводящих тканей, а далее, к периферии, хорошо развитой корой. Как мы видим, многие особенности внешнего и внутреннего строения беннеттитовых являются общими с саговниковыми. Сходно и строение проводящих тканей. Первичная ксилема во внутренней их части представляла собой кольцо анастомозирующих; коллатеральных пучков, окруженная снаружи сплошным слоем вторичной ксилемы (эцдархцая эвстела).
Сходство между Двумя рассматриваемыми классами голосеменных распространялось и на листья. У беннеттитовых, как и у большинства современных (а также и вымерших) саговниковых, они имели ксероморфное строение, были жесткими, кожистыми, обычно однажды перистыми, реже цельными, линейными.
Только после того, как в палеоботанике начали широко применять анатомические методы исследования и стала известна структура эпидермы мумифицированных листьев, оказалось, что значительная часть мезозойских растений, внешне столь похожих на саговниковые, существенно от них отличается (рис. 171). Из раздела о саговниковых читатель уже узнал о кардинальных различиях между теми и другими в строении эпидермы и особенно в характере устьичного аппарата (сйндетохейльный5 тип устьиц у беннеттитовых). Важные отличия выявились и в строении репродуктивных органов. Все это дало основание выделить самостоятельную группу голосеменных - беннеттитовые.
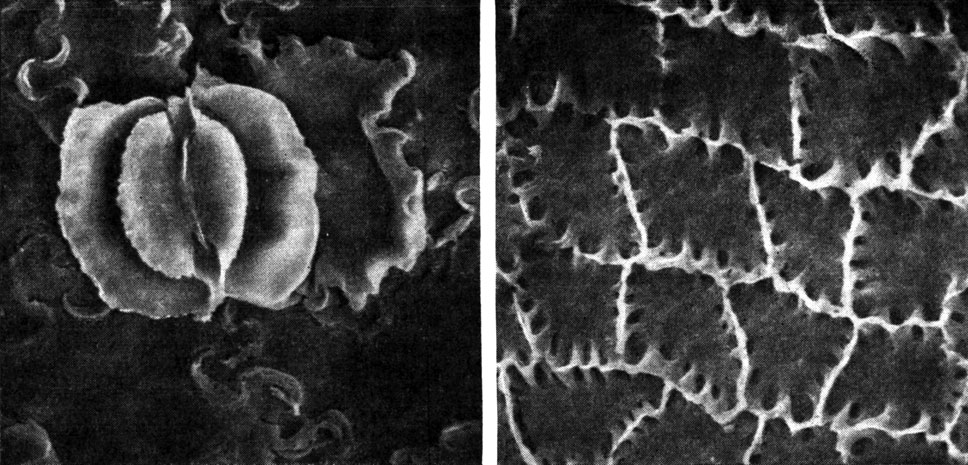
Рис. 171. Нильсониоптерис лентовидный (Nilssoniopteris taeniata) из меловых отложений Приморья, эпидерма листа (сканирующий электронный микроскоп): слева - устьице на нижней эпидерме, окруженное клетками с синусоидально извилистыми стенками - вид изнутри (увел, около 1000); справа - верхняя эпидерма из клеток с неравномерно утолщенными стенками (увел, около 500)
Стробилы беннеттитовых по своему строению были большей частью обоеполыми, реже однополыми. В неизбежно кратком здесь описании размещения мегаспорангиев в стробилах можно дать представление и о мегастробилах, когда стробилы однополы, и о верхней части обоеполых стробилов - они однотипны. Общий план их строения, очевидно, свидетельствует о генетическом единстве всех многообразных беннеттитовых. Как же были устроены мегастробилы?
Основой мегастробила была мягкая мясистая ось, являвшаяся продолжением его относительно короткой ножки. Это утолщенное образование, обычно конической или булавовидной формы, можно назвать семяложем, поскольку на нем на коротких ножках (редуцированных мегаспорофиллах) сидели многочисленные семязачатки (семена), окруженные со всех сторон межсемянными чешуями. Верхушки межсемянных чешуй были расширены и смыкались краями, хорошо защищая семязачатки. Снаружи такое образование напоминало шишку, составленную из плотно пригнанных полигональных чешуй, между которыми в углах чуть выдавались небольшие столбики - верхушки микропилярных трубок семязачатков. На одном семяложе беннеттитового стробила могло образоваться до нескольких сотен семян. Семена были довольно мелкими, обычно не более рисового зернышка. К основанию семяложа в спиральном порядке прикреплялись многочисленные, довольно крупные покроволистики, обычно густо покрытые волосками. Как и межсемянные чешуи, они выполняли защитную функцию: смыкаясь над стробилом, они функционировали, как чашечка у цветков покрытосеменных.
Зрелые семена беннеттитовых представляют особый интерес. В то время как у семенных папоротников зародыш, очевидно, развивался уже только после опадения семени с растения, а у саговниковых доразвивался в семени, упавшем на почву, у многих беннеттитовых к моменту созревания семени на растении зародыш был уже вполне развитым. Мало того, у этого зародыша, заполнявшего почти все семя, развивались две крупные мясистые семядоли, служившие местом отложения запасных питательных веществ (как в семенах гороха, фасоли, ивы, одуванчика). Такие семена несвойственны не только остальным голосеменным, как ныне живущим, так и вымершим, но и прими-тивным цветковым, и появление их у беннеттитовых представляло собой большой шаг вперед в процессе эволюции.
Микроспорофиллы у беннеттитовых с обоеполыми стробилами располагались мутовчато между охарактеризованными выше покроволистиками и семяложем с семязачатками. Строение их в этом случае, а также в микростробилах по сравнению с мегаспорофиллами было гораздо более разнообразным.
Беннеттитовые можно подразделить на две группы, которые обычно рассматриваются как семейства: вильямсониевые (Williamsoniaceae) и беннеттитовые (Bennettitaceae).
Вильямсониевые (Williamsoniaceae). С вильямсониевых, по существу, и началось изучение класса беннеттитовых. Особенно много для познания беннеттитовых дала юрская флора Йоркшира в Англии, которая на протяжении более чем столетия тщательно изучалась несколькими поколениями палеоботаников. Однако в начале XIX в. первые исследователи этой флоры, и беннеттитовых в частности, еще не предполагали о существовании новой самостоятельной группы голосеменных и относили ли-стья, напоминающие листья саговниковых, и сопутствующие им другие органы растений к саговниковым.
Самый обширный род семейства - вильямсония (Williamsonia) - назван в честь известных английских исследователей отца и сына Вильямсонов, очень много сделавших для его познания. Первой из изученных вильямсоний была вильямсония гигантская (W. gigas). Это было, по-видимому, неветвящееся прямоствольное растение высотой около 2 м, с кроной крупных перистых листьев на верхушке и с рубцами от опавших листьев на стволе. Среди листьев формировались однополые стробилы, но пока остается неизвестным, были ли вильямсонии одно- или двудомными растениями, т. е. образовывались ли микро- и мегастробилы на одном и том же растении или на разных.
Мегастробилы вильямсоний довольно крупные (диаметром до 8 см и более), но они не отличались каким-либо ярким морфологическим своеобразием и были устроены по "стандартному" для беннеттитовых плану. Микростробилы вильямсоний, известные в палеоботанической литературе под названием вельтрихий (Weltrichia), были похожи на крупную чашу высотой 7-8 см, края которой надрезаны на многочисленные (около 30) клиновидные сегменты. На внутренней поверхности этих сегментов имелись короткие выросты, на которых сидели синангии, состоящие из двух рядов сросшихся микроспорангиев. Микроспорангии заключали большое количество овальных однобороздных микроспор. Интересно, что в сросшейся базальной части микростробила находились секреторные образования, возможно, выполнявшие функцию нектарников и служившие для привлечения насекомых.
С триаса по ранний мел включительно род вильямсония был широко распространен на Земле и представлен многими видами. Удачную реконструкцию вильямсонии Сьюорда (Williamsonia sewardiana), существовавшей в юрский период в Индии, можно видеть на рисунке 170. Это было небольшое слабо ветвящееся древовидное растение со стробилами на верхушках боковых побегов. Среди вильямсоний были и сильно ветвящиеся растения, как, например, вильямсония Лекенби (Williamsonia leckenbyi) из юры Йоркшира. Судя по обилию их листовых остатков, в некоторых районах они могли быть доминирующими в юрских лесах.
Следует упомянуть еще о вильямсониелле (Williamsoniella). Виды этого рода скорее всего были сильно ветвящимися кустарниками. Вильямсониелла корончатая (W. coronata) - одно из наиболее полно изученных растений семейства. На ее тонких вильчато ветвящихся побегах в спиральном порядке сидели цельные линейные листья длиной около 10 см. Ее обоеполые стробилы, в отличие от стробилов вильямсоний, были довольно мелкими, не более цветка лютика, и сидели на относительно длинных ножках. На тонком конусовидном семяложе образовывалось более 300 крошечных семян и около 1200 межсемянных чешуй. Стерильная верхушка семяложа возвышалась над стробилом как корона, с чем и связан видовой эпитет этого растения. К основанию семяложа мутовчато прикреплялись 12-14 не сросшихся между собой микроспорофиллов, по своему строению отличающихся от микроспорофиллов вильямсоний. Они были сочные, мясистые, а по форме напоминали дольки апельсина. Посредине микроспорофилла находились 2 (реже 3) пары пальцевидных выростов, к которым прикреплялись синангии. Снаружи стробил был защищен несколькими рядами покроволистиков, густо покрытых волосками. Во время опыления покроволистики и микроспорофиллы, по-видимому, широко раскрывались, а впоследствии опадали.
Уже из приведенных примеров видно, что вильямсониевые отличались разнообразием и были широко распространены. Находки вильямсониевых известны почти на всех континентах. В основном они населяли области жаркого климата, но существовали и в условиях умеренно теплого климата.
Беннеттитовые (Bennettitaceae). Эти растения были более монолитной группой, чем вильямсониевые, и населяли только области жаркого засушливого климата. В отличие от вильямсониевых виды этого семейства имели толстые и короткие стволы (высотой обычно не более 1 м), как правило, неветвящиеся и с кроной крупных однажды перистых листьев на верхушке (рис. 172). Молодые листья, как и у папоротников, были спирально свернутыми. Стробилы беннеттитовых, расположенные по бокам ствола, были защищены частоколом из оснований черешков отмерших листьев.

Рис. 172. Цикадеоидея (Cycadeoidea). Реконструкция целого растения
Окаменелые стволы цикадеоидей (Cycadeoidea) - самого крупного и широко распространенного рода беннеттитовых (рис. 172) - часто встречаются на территории Северной Америки, известны также из стран Западной Европы, из Монголии, Индии, Японии. Такой окаменелый ствол цикадеоидеи, обнаруженный в Италии, недалеко от Болоньи, в древнем этрусском некрополе, пожалуй, самый старый из известных нам палеонтологических объектов, обративших на себя внимание человека. Этот прекрасно сохранившийся, почти черный ствол своим необычным обликом привлек внимание этрусков более 4000 лет назад и был водружен на могиле как надгробный памятник. Во второй половине XIX в. он был найден вторично, передан в руки ученых и описан под названием цикадеоидея этрусков (Cycadeoidea etrusca).
Цикадеоидея получила широкую известность благодаря эффектной реконструкции ее стробила, предложенной в начале XX в. американским ученым Д. Уиландом. Реконструкция, подробно разработанная в нескольких планах - ив стадии "бутона", и в виде распустившегося "цветка", увлекла ботаников очевидным сходством с крупными цветками архаичных покрытосеменных и невольно наталкивала на мысль, не являются ли беннеттитовые примитивными покрытосеменными или хотя бы их предками. Среди изданных в XX в. учебников ботаники и палеоботаники трудно найти такой, где не была бы воспроизведена эта созданная Д. Уиландом реконструкция.
Стробилы цикадеоидей были обоеполыми (рис. 173). Основой стробила, как и у всех беннеттитовых, являлось мясистое семяложе, на котором располагались многочисленные семена и межсемянные чешуи. К основанию семяложа мутовчато прикреплялись микроспорофиллы. Снаружи стробилы были защищены многочисленными густо опушенными покроволистиками. Д. Уиланд представлял себе микроспорофиллы как перистые образования, близкие по строению к фертильным листьям папоротников. Ко времени опыления, по Уиланду, микроспорофиллы широко раскрывались вместе с покроволистиками. Соответственно предполагалось, что цикадеоидеи были ветроопыляемыми растениями. Однако до сих пор раскрытые стробилы не встречены. Учитывая то, что на одном стволе цикадеоидей могло образовываться до 600 стробилов, а их окаменелых стволов известно уже более 1000, трудно ожидать, что раскрывание стробилов имело место. Это подтвердилось новейшими исследованиями американских палеоботаников Т. Делевориаса и др. Они внесли существенные изменения в наши представления о строении стробилов цикадеоидей, в особенности их микроспорофиллов. Оказалось, что микроспорофиллы цикадеоидей были довольно сложно устроенными специализированными образованиями. Каждый из микроспорофиллов был похож на сочную дольку апельсина, в которой удалена центральная часть. "Окно" было заполнено "перекладинами", расположенными попарнорадиально друг над другом. К ним-то и прикреплялись овальные спорангии. Все микроспорофиллы одного стробила срастались в основании. По Делевориасу, устройство стробилов цикадеоидей было таким, что, очевидно, исключало возможность ветроопыления: семяложе с готовыми к восприятию пыльцы семязачатками было окружено массивными, явно не раскрывавшимися широко микроспорофиллами, причем в значительной части сросшимися, затем прикрыто несколькими рядами покроволистиков и, наконец, защищено основаниями черешков листьев, среди которых находился стробил. Против возможности ветроопыления свидетельствуют и размеры пыльцевых зерен цикадеоидей, достигавших 40-90 мкм (у современных ветроопыляемых растений они не более 30 мкм). Предполагается, что основным способом опыления у цикадеоидей было самоопыление. Однако перекрестное опыление у них время от времени могло осуществляться при помощи насекомых. Действительно, стробилы цикадеоидей нередко встречаются со следами повреждения насекомыми (скорее всего это были жуки), которых, очевидно, привлекали сочные ткани семяложа, микроспорофиллов, а возможно, и сами семязачатки.
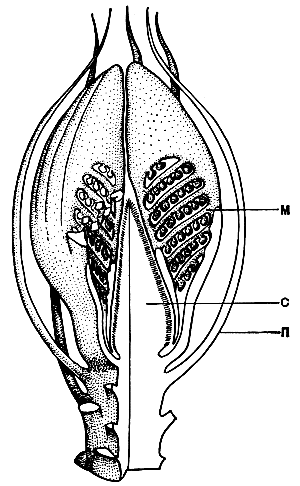
Рис. 173. Реконструкция стробила цикадеоидеи (Gycadeoidea). Части стробила, не покрытые точками, показаны в разрезе : с - семяложе с семенами (семязачатками) и межсемянными чешуями; м - микроспорофиллы со спорангиями; п - покроволистики
Окаменелые стволы цикадеоидей иногда демонстрируют великолепную сохранность. Примером феноменальной сохранности является обнаруженный среди американских образцов стробил с прекрасно сохранившимися семязачатками на начальной стадии мегаспорогенеза. В недрах нуцеллуса среди паренхимных клеток видна даже линейная тетрада мегаспор, что представляет древнейший "документ" о наличии у голосеменных этой фазы мегаспорогенеза.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'