Класс мараттиопсиды (Marattiopsida)
Порядок мараттиевые (Marattiales)
Семейство мараттиевые (Marattiaceae)
В каменноугольном и пермском периодах палеозойской эры мараттиевые занимали большие пространства на земном шаре и местами составляли доминирующий элемент в растительном покрове. Их стройные колонновидные стволы, увенчанные кроной из огромных перистых листьев, возвышались над землей на 10-15 м. Окаменелые остатки стволов этих папоротников, известные под названием псарониус (Psaronius, рис. 99), были найдены в каменно-угольных отложениях всех континентов, вплоть до 70° с. ш., далеко за пределами современного ареала мараттиевых, не выходящего за 30° с. ш. В близком соседстве с остатками псарониуса часто находят и листья мараттиевых, относимые палеоботаниками к сборному роду пекоптерис (Pecopteris), и еще более часто их спороношения, которым даны различные названия: сколекоптерис (Scolecopteris), астеротека (Asterotheca), ацитека (Acitheca) и др. И хотя в последнее время было показано, что аналогичные типы листьев и спороношений существовали и в других группах вымерших растений, например у некоторых голосеменных, явное сходство ископаемых остатков с соответствующими структурами современных мараттиевых оставляет мало сомнений в принадлежности тех и других к одной группе растений. В мезозойских отложениях находят типы мараттиевых, уже почти совсем неотличимые от современных родов.

Рис. 99. Псарониус (Psaronius). Реконструкция
Высокий, конической формы ствол псарониуса на первый взгляд был похож на стволы древесных растений, но по своей структуре был необычен. Собственно стебель у основания был диаметром несколько миллиметров, кверху он постепенно утолщался и достигал толщины у верхушки 20 см. Конечно, такое растение, при высоте свыше 10 м, не могло бы существовать и удерживаться в прямостоячем положении. Проблема эта решалась путем развития плотного покрова из придаточных корней, в который был завернут стебель. Покров из корней достигал особенно большой толщины у основания стебля, а вверх он постепенно уменьшался, по мере того как становился толще стебель. Придаточные корни, интенсивное образование которых наблюдается и у других древовидных папоротников, как и у современных мараттиевых, закладывались в наружном кольце стелы, пронизывали кору и выходили в поверхностную зону стебля, оставаясь в окружении клеток субэпидермальной паренхимы. Слои этих корней, составлявших единое целое со стеблем, образовывали внутреннюю корневую мантию. Часть корней, которая свободно свисала между основаниями отмерших листьев, переплетаясь с ними, составляла наружное влагалище стебля. В такой двойной обкладке ствол был вполне устойчив и мог поддерживать крону мощных листьев, достигавших в длину 3 м.
Не менее интересны ископаемые псарониусы и по структуре своей проводящей системы, сохранившейся в несколько упрощенном виде и у современных мараттиевых. У основания ствол имел амфифлойную сифоностелу, которая выше постепенно превращалась в сложную полициклическую диктиостелу, состоящую из нескольких (до 12) концентрических кругов с большими прорывами. Проводящая система этих папоротников являлась как бы системой вложенных один в другой перфорированных конусов. Прорывы каждого наружного конуса заполнялись тяжами проводящей ткани от внутреннего.
Большая часть мараттиевых вымерла за прошедшие миллионы лет, и до наших дней сохранилось всего 7 родов этих живых ископаемых, обитающих в наше время в тропических областях. И этот свой титул самых древних из ныне существующих папоротников они оправдывают набором редких, не свойственных другим папоротникам признаков.
Вместе с ужовниковыми мараттиевые составляют группу так называемых эвснорангиатных папоротников, ибо только у них спорангии развиваются не из одной, как у лептоспорангиатных папоротников, а из нескольких субэпидермальных клеток. Спорангии мараттиевых имеют массивные многослойные стенки, снабженные устьицами. Для них характерно также отсутствие специализированного кольца; спорангии вскрываются здесь щелью или посредством пор. Нет у них и выраженного покрывальца, а есть только чешуйки или волоски в основании сорусов. У некоторых мараттиевых спорангии свободно лежат сдвоенными рядами на жилках, у других же они, срастаясь, образуют своеобразную, не встречающуюся у лептоспорангиатных папоротников структуру - синангий.
Крупные листья мараттиевых имеют у своего основания характерные, не свойственные другим папоротникам прилистниковидные придатки, которые сохраняются на стеблях после опадания листьев и служат не только защитными, но и запасающими органами (в них накапливается крахмал). Эти придатки служат также для размножения, так как на них имеются покоящиеся почки, которые при благоприятных условиях дают начало новым растениям. В листьях, стеблях и корнях мараттиевых всегда присутствуют крупные лизигенные слизевые ходы. Интересно, что эта особенность свойственна также древнейшей группе современных голосеменных - саговниковым.
Мараттиевые сохранились в дождевых тропических лесах, часто в местах со сложным горным рельефом, там, где климатические условия не подвергались значительным переменам с древних времен. Ныне они не являются древовидными папоротниками в точном смысле этого слова, хотя и достигают иногда очень значительных размеров. Принадлежащие к этому семейству ангиоптерис (Angiopteris), распространенный в тропиках восточного полушария, макроглоссум (Macroglossum) с двумя видами на Калимантане и Суматре и мараттия (Marattia) с 60 видами в тропиках обоих полушарий относятся к числу самых крупных папоротников на Земле. Огромный их размер обусловлен, однако, не стеблем, а крупными, длиной до 6 м, листьями. Стебли же этих папоротников небольшие, редко достигают в высоту 1 м, имеют клубнеобразный вид и часто наполовину скрыты в почве.
Высокие, благородного облика ангиоптерисы составляют одну из поразительных черт богатой тропической флоры. По влажным лесным склонам, по тенистым ущельям, в болотистых лесах группы этих папоротников встречаются на огромной территории от Мадагаскара до тропической Азии, Полинезии и Северо-Восточной Австралии, на севере достигая Гималаев. Растут они и вдоль дорог, среди бамбуковых рощ, по крутым речным берегам. Огромные, длиной до 4-6 м, обычно дважды перистые листья ангиоптериса с черешками толщиной в человеческую руку спирально расположены на шаровидном стебле, покрытом основаниями старых листьев и многочисленными мясистыми прилистниками. Главный черешок листа и вторичные черешочки утолщены в местах многочисленных сочленений, благодаря чему черешок нередко напоминает бамбуковый стебель. Предполагают, что эти утолщенные сочленения, свойственные и другим мараттиевым, участвуют в важном физиологическом механизме изменения ориентации листьев по отношению к падающим на них солнечным лучам.
В общих чертах все папоротники этого рода обнаруживают между собой столь большое сходство, что иногда их всех относят к одному крупному виду - ангиоптерису вознесенному (Angiopteris evecta, рис. 100). Иногда род разбивают на многочисленные (более 100) самостоятельные виды.
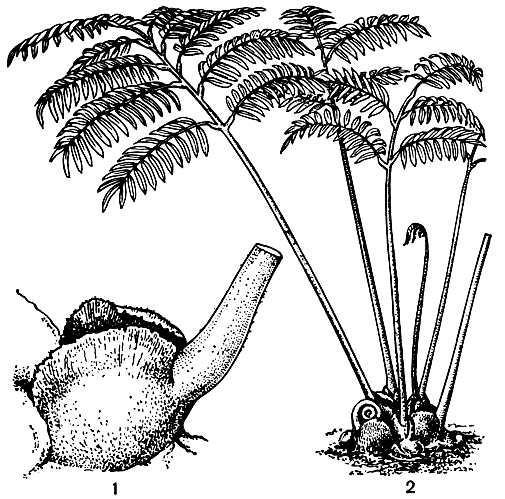
Рис. 100. Ангиоптерис вознесенный (Angiopteris evecta): 1 - основание листа с прилистниками; 2 - общий вид
Листьям всех видов ангиоптериса свойственно свободное жилкование и наличие так называемых ложных или возвратных жилок. Эти возвратные жилки располагаются в виде полосок между двумя зубцами, они не содержат проводящей ткани и соответствуют но структуре краю листового сегмента. Предполагают поэтому, что ангиоптерис имел когда-то более рассеченные листья, и ложные жилки очерчивают границы бывших мелких сегментов. На нижней стороне листьев на боковых жилках ближе к краю листа лежат эллиптической или удлиненной формы сорусы, включающие до 20 свободных спорангиев, расположенных в 2 ряда (табл. 19). В отличие от большинства других мараттиевых на верхушке спорангиев ангиоптериса присутствует полоска клеток с более толстыми стенками, ориентированная поперек к продольной линии растрескивания спорангия. Эта полоска клеток еще не действует как специализированное кольцо других папоротников, но в соединении со всеми поверхностными клетками она способствует более широкому открыванию спорангиев. У ангиоптериса преобладают тетраэдрические споры, но изредка встречаются и билатеральные. В каждом спорангии насчитывается до 1450 спор.
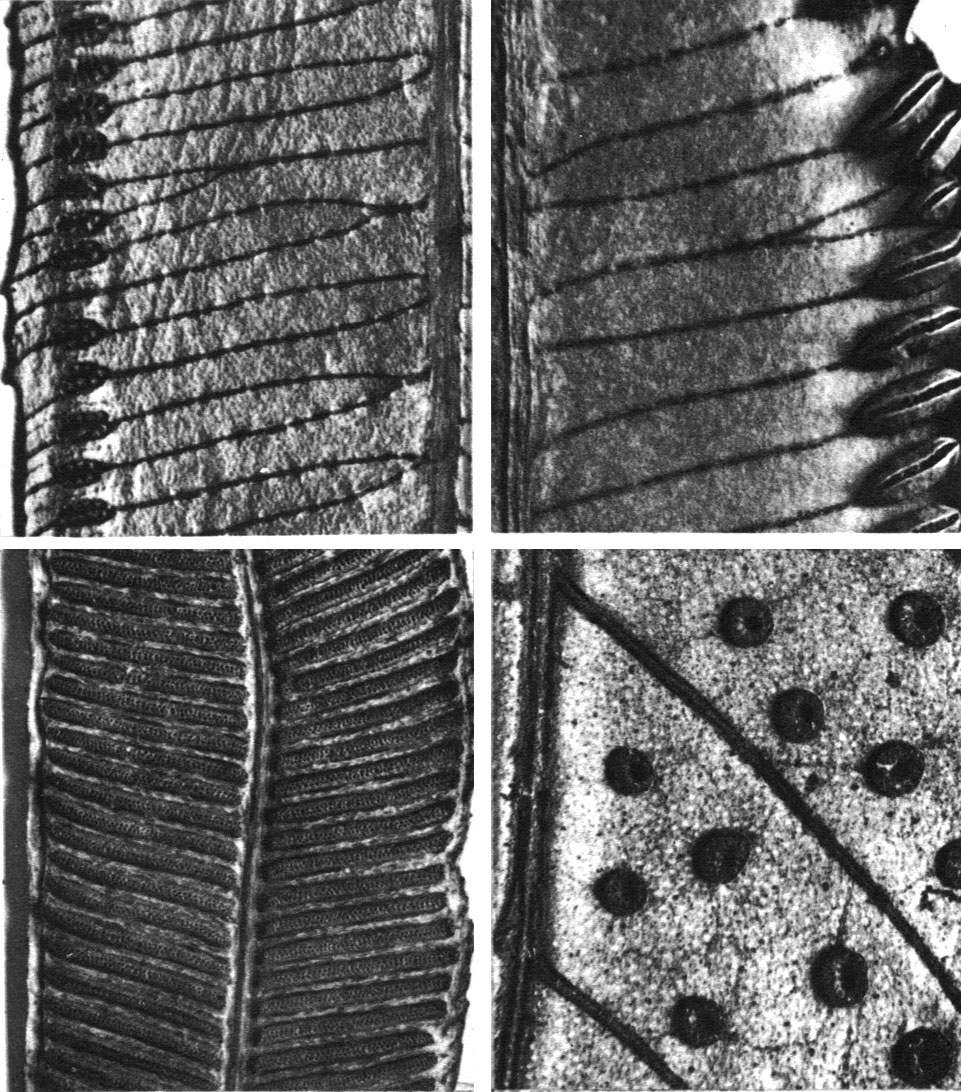
Таблица 19. Типы спороношений у папоротников семейства мараттиевых: вверху слева - ангиоптерис вознесенный (Angiopteris evecta); вверху справа - мараттия ясеневая (Marattia fraxinea); внизу слева - данея эллиптическая (Danea elliptica); внизу справа - кристенсения каштанолистная (Christensenia aesculifolia)
В зарослях ангиоптериса, там, где наряду со взрослыми растениями часто встречаются и молодые спорофиты, внимательный наблюдатель всегда может отыскать и своеобразные гаметофиты этих папоротников. Внешне темно-зеленые пластиночки гаметофитов диаметром 1-3 см похожи на талломные печеночники. У ангиоптериса они имеют сердцевидную или округлую форму и относительно толсты, особенно в центральной части, которая всегда содержит много слоев клеток, образуя как бы подушечку. С нижней стороны на этой подушечке и близ нее развиваются массивные погруженные антеридии, а позднее также архегонии (последние небольшими группами появляются и на верхней стороне). Живут гаметофиты ангиоптериса несколько лет. Старые гаметофиты могут дихотомически ветвиться, они продолжают свой рост иногда и после оплодотворения и образования спорофита.
Характерным признаком гаметофита ангиоптериса, как и других мараттиевых, является присутствие в тканях его центральной утолщенной части эндофитного гриба стигеоспориума мараттиевого (Stigeosporium marattiacearum). Инфицированы грибом обычно и первые корни мараттиевых. Но микориза является здесь факультативной, так как папоротник может нормально развиваться и без взаимодействия с грибом, и это сожительство не является для них жизненно необходимым.
Папоротники рода мараттия по внешнему облику похожи на ангиоптерис, но менее крупные, обычно высотой 1-2 м, иногда встречаются экземпляры высотой более 3 м, со стеблем диаметром до 20 см.
Как и у ангиоптериса, спорангии расположены вдоль свободных жилок ближе к краям сегментов многократно перистых листьев, но, в отличие от ангиоптериса, они здесь срослись в линейные синангии (табл. 19). Каждые два синангия, лежащие по обеим сторонам жилки, в свою очередь, срослись друг с другом. По созревании весь сложный синангий раскрывается над жилкой двумя створками, а каждое гнездо синангия растрескивается внутрь брюшной щелью. Казалось бы, столь принципиальное различие в строении спороношений ангиоптериса и мараттии не позволяет считать эти роды близкородственными. Однако изучение развития спорангиев в онтогенезе показало, что свободные спорангии ангиоптериса не имеют четко выраженной морфологической определенности при заложении, они закладываются как непрерывный линейный ряд, подобно синангию мараттии. Специалисты считают поэтому, что роды мараттия и ангиоптерис очень близки, а свободное положение спорангиев у мараттиевых происходит от синангиатного. Интересно отметить, что оба типа спороношений являются очень древними, так как они встречаются уже у мараттиевых каменноугольного периода.
И ангиоптерис и мараттия не имеют достаточно развитой механической ткани в листьях. Поэтому при подсыхании корней мясистые листья этих растений, быстро теряя тургор, становятся вялыми и поникают, складываясь и провисая в местах многочисленных утолщенных сочленений, которые также теряют воду. В таком виде они могут существовать довольно долго без заметного ущерба для себя, восстанавливая тургор при возобновлении водоснабжения. Это явление можно наблюдать в оранжереях, где виды ангиоптериса и мараттии охотно выращивают как декоративные растения.
Иногда мараттии культивируют и как съедобные. Их сочные листья использует в пищу в жареном, вареном или печеном виде аборигенное население Новой Зеландии, Антильских островов, Мексики и Бразилии. С этой же целью используют также стебли и прилистники, а слизь из прилистников иногда применяют в этих странах как лекарство.
В муссонных лесах Юго-Западного Китая (Юньнань), Тайваня и Северного Вьетнама (Тонкин) наряду с ангиоптерисом встречается еще один представитель мараттиевых - архангиоптерис (Archangiopteris), включающий около 10 видов, которые, хотя и родственны ангиоптерису по расположению и строению свободных спорангиев, внешним обликом резко от него отличаются (рис. 101). Они гораздо меньшего размера, с горизонтальным ползучим или косым дорсивентральным корневищем. На верхней стороне корневища тесно сидят перистые (однажды перистые) листья с длинными черешками, покрытыми решетчатыми, часто щитовидно прикрепленными чешуйками. Черешки листа снабжены обычно одним утолщенным сочленением.

Рис. 101. Архангиоитерис хэкоуский (Archangiopteris hokouensis). Видны утолщения на черешках листьев
Такой же тип корневища свойствен и роду кристенсения (Christensenia), представленному единственным видом кристенсенией каштанолистной (С. aesculifolia) в лесах Индии, Индонезии, Филиппин. Его мясистое корневище несет два ряда сближенных листьев необычной для папоротников пальчатой формы. Листья, напоминающие по виду листья конского каштана (Aesculus hippocastanum), состоят из 3-5 листочков, присоединенных короткими черешочками к главному длинному (до 70 см) черешку, превосходящему в два раза по длине пластинку листа. От других мараттиевых листья кристенсении отличаются еще сетчатым жилкованием и особыми округлыми синангиями, разбросанными на анастомозах жилок по всей нижней поверхности листа (табл. 19). Каждый синангий состоит из 10-15 сросшихся спорангиев (гнезд синангия), расположенных кольцом вокруг углубления в центре. Каждый спорангий открывается верхушечной щелью. Такой набор несвойственных другим мараттиевым признаков говорит о том, что кристенсения - остаток своеобразной самостоятельной эволюционной ветви этой группы папоротников.
Название кристенсения дано в честь известного птеридолога Карла Кристенсена, автора фундаментального труда "Index filicum" ("Указатель папоротников").
Род данея (Danaea) включает 30 видов, встречающихся, в отличие от других мараттиевых, исключительно в тропиках западного полушария. Виды этого рода, отличающиеся экзотической красотой, растут в теплых, постоянно влажных и обычно тенистых местах. Листья даней перистые, иногда цельные (у даней простолистной - D. simplicifolia). Фертильные листья всегда на более длинных черешках и с более узкими сегментами. Синангии у рода данея удлиненные (до 3 см), обычно погруженные в бороздку.
Срастание спорангиев у даней столь полное, что отдельные спорангии, а их насчитывается в каждом синангии до 100, слаборазличимы. Вскрывание гнезд синангия даней происходит путем образования верхушечной поры.
Как видно из приведенного описания, современные роды мараттиевых представляют собой неоднородную группу растений, различающихся между собой как характером спороношений, так и строением вегетативных органов. Происходит это потому, что они являются всего лишь фрагментарными остатками некогда большой и многообразной группы папоротников. Многие птеридологи разделяют поэтому современные мараттиевые на несколько самостоятельных подсемейств или даже семейств. Известный американский морфолог Д. Бирхорст выделяет два семейства - собственно мараттиевые (Marattiaceae) и данеевые (Danaeaceae).
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'