Спорангии и споры
На определенной стадии развития спорофита начинается его спороношение - явление, в биологическом отношении аналогичное цветению. Внутренние физиологические процессы, вызывающие переход спорофита из вегетативной фазы развития в фазу спороношения, еще далеко не разгаданы, но, вероятно, они имеют много общего с теми процессами, которые приводят к цветению. Спороношение начинается с закладки тех начальных (инициальных) клеток или групп клеток, из которых развивается спорангий. Как мы уже знаем, в одних случаях спорангии развиваются на обыкновенных зеленых листьях, в других случаях - на специальных спороносящих частях листа или на особых листьях, резко отличающихся от вегетативных и специализированных для функции спороношения. Обособление фертильных участков и специализированных фертильных листьев (спорофиллов) - явление в эволюционном смысле вторичное.
Интересно, что, в то время как у ужовниковых и мараттиевых (Marattiaceae) каждый развивается из группы начальных клеток и имеет многослойную стенку, почти у всех остальных современных папоротников он развивается из одной-единственной начальной клетки и имеет однослойную стенку (табл. 19, 20). Очевидно, при втором способе развития спорангия требуется меньше строительного материала (меньше клеток), а само развитие происходит быстрее. Поэтому все согласны с тем, что возникновение спорангиев, берущих начало от одной клетки и имеющих однослойную стенку, представляет в эволюционном отношении шаг вперед.
Вспомним еще раз, что листья папоротников возникли в процессе эволюции из ветвей риниофитовых предков. Отсюда вполне понятно, что у древнейших папоротников спорангии имели верхушечное (терминальное) расположение, т. е. сидели на верхушках конечных веточек дихотомически разветвленного листа. Среди ныне живущих папоротников расположение спорангиев, наиболее близкое к верхушечному, наблюдается у гроздовника (Botrychium), относящегося к примитивному семейству ужовниковых.
По мере того как примитивный, похожий еще на ветку лист уплощается, между отдельными веточками образуется как бы перепонка (подобно плавательной перепонке водоплавающих птиц), представляющая собой результат бокового разрастания ткани веточек. Образование перепонки увеличило фотосинтезирующую поверхность листа, что дало огромное преимущество таким растениям. Спорангии остались, в сущности, на своих прежних местах. Но так как верхушки веточек оказались у нового органа растения - листа - концами жилок (ведь каждая веточка жилкования соответствует первоначальному проводящему пучку веточки), спорангии автоматически оказываются на краю пластинки листа. Таким образом верхушечное расположение спорангиев переходит в краевое (маргинальное). Трудно сказать, было ли это на самом деле шагом вперед или только естественным результатом возникновения листовой пластинки. Во всяком случае, краевое расположение спорангиев наблюдается у многих современных папоротников.
Значительным шагом вперед в приспособительной эволюции папоротников был переход спорангиев из краевого расположения в поверхностное (ламинальное). Этот процесс происходил независимо в разных линиях эволюции, причем во всех случаях спорангии перемещались на нижнюю, или "брюшную" (абаксиальную), сторону листовой пластинки. Это вполне понятно, так как на брюшной стороне они больше защищены от всяких неблагоприятных воздействий внешней среды и находятся в лучших условиях питания и рассеивания зрелых спор. Кроме того, если бы спорангии перешли на верхнюю, или "спинную" (адаксиальную), сторону пластинки, они бы заняли некоторую часть наиболее активной фотосинтезирующей поверхности, что неизбежно отразилось бы на эффективности фотосинтеза. Поэтому не удивительно, что во всех без исклю-чения случаях поверхностного расположения спорангиев они занимают нижнюю сторону листовой пластинки. У ныне живущих папоротников наблюдаются все переходные стадии от краевого расположения к поверхностному (а последнее встречается чаще других). Более того, у некоторых из тех папоротников, спорангии которых сидят вдоль края пластинки, это перемещение из краевого положения в поверхностное осуществляется в процессе онтогенеза.
Наряду с перемещением спорангиев на нижнюю сторону пластинки происходит еще и другое, не менее важное изменение в их расположении. Сначала спорангии бывают расположены одиночно, т. е. не образуют групп, как, например, у схизейных. Но уже у некоторых папоротников с краевым расположением, как, например, у диксонии (Dicksonia) и локсомы (Loxsoma) или у всех гименофилловых, спорангии сгруппированы в отдельные спорангиальные кучки, или сорусы (от греч. soros - куча). У папоротников с поверхностным расположением спорангиев это уже становится правилом. Возникновение сорусов связано прежде всего с увеличением числа спорангиев. Так, если у предков диксонии на конце жилки сидело лишь по одному спорангию, то у самой диксонии - целая спорангиальная кучка. Но не менее важно и то, что с образованием сорусов возникает возможность более концентрированного и, следовательно, более эффективного снабжения спорангиев питательными веществами.
В биологическом отношении чрезвычайно важен порядок развития и созревания спорангиев на листе или в каждом отдельном сорусе. У вымерших папоротниковидных и у ныне живущих ужовниковых, мараттиевых, осмундовых, плагиогириевых, схизейных, глейхениевых и матониевых все более или менее близко расположенные на растении спорангии или все спорангии внутри соруса (плагиогириевые, глейхениевые и матониевые) развиваются одновременно (симультанно). Сорусы, в которых развитие и созревание спорангиев происходит одновременно, называют простыми.
Одновременное развитие спорангиев имеет один существенный недостаток. Дело в том, что при ухудшении условий произрастания и физиологическом угнетении растения подвергается риску развитие одновременно формирующихся спорангиев на части растения или даже на всем растении. Выходом из этого положения является последовательное развитие спорангиев, когда их созревание в сорусе наступает в разное время. Это достигается посредством удлинения ложа (рецептакула) соруса и увеличения количества спорангиев при одновременном уменьшении их размеров. Устанавливается так называемая базипетальная (от греч. basis - основание и лат. реtere - устремляться) последовательность, когда развитие начинается с верхней части ложа (в центре соруса) и постепенно переходит к основанию ложа (к периферии соруса). В результате самые молодые спорангии оказываются наиболее близко расположенными к источнику снабжения питательными веществами и наиболее защищенными. Все развитие соруса растягивается во времени, и поэтому процесс спороношения оказывается сравнительно менее уязвимым (если пострадают более ранние спорангии, то будет еще шанс для развития более поздних). Особенно хорошо выражено базипетальное развитие спорангиев у представителей семейства гименофилловых, имеющих длинное цилиндрическое или линейное ложе. Сорусы с базипетальным развитием спорангиев называют градатными (от лат. gradatio - постепенное возвышение, усиление). Такие сорусы характерны для циатейных, гименофилловых, асплениевых, а также для всех разноспоровых папоротников.
В некоторых линиях эволюции папоротников из градатных или прямо из простых сорусов возникают так называемые смешанные сорусы, характеризующиеся неопределенной последовательностью в развитии спорангиев. В таких сорусах молодые и более зрелые спорангии расположены вперемешку, без определенного порядка. Несмотря на то что ложе таких сорусов обычно плоское, спорангии имеют разную высоту, так как у более зрелых спорангиев ножка удлиняется, и поэтому они заметно возвышаются над более молодыми. Преимущество смешанных сорусов заключается прежде всего в том, что развитие спорангиев в сорусе растягивается во времени. Но не менее важно то обстоятельство, что молодые спорангии здесь надежно защищены возвышающимися над ними зрелыми спорангиями, оставшимися ножками уже раскрывшихся спорангиев, а также волосками, которые часто сидят на ложе между спорангиями. Исключительно важно также, что в смешанных сорусах может эффективно и без всяких физических помех действовать наиболее совершенный способ раскрывания спорангиев.
Смешанные сорусы возникают независимо в разных линиях эволюции папоротников, притом они происходят как от простых, так и от градатных сорусов. Происхождение от простых сорусов можно наблюдать в пределах рода диптерис, где у одного из видов (диптерис Лобба - Dipteris lobbiana) развитие спорангиев симультанное, в то время как у более подвинутого вида - диптериса двойчатосложного (D. conjugata), развитие спорангиев происходит в неопределенной последовательности (сорусы смешанные). У близкого к диптерисовым и, вероятно, производного от него семейства хейроплевриевых (Cheiropleuriaceae) сорусы также смешанные. У большого и широко распространенного семейства полиподиевых, имеющего общее происхождение с двумя выше упомянутыми маленькими семействами, также установилась неопределенная последовательность в развитии спорангиев. Таков же порядок развития у другого большого семейства, адиантовых.
В эволюции сорусов произошло еще одно важное событие. Речь идет о защитных приспособлениях спорангиев. Вообще защита молодых спорангиев у папоротников обеспечивается хотя бы тем, что их молодые листья скручены спирально как раковина улитки, причем не только по главной жилке, но и по всем сегментам. Это проявилось уже у некоторых вымерших примитивных родов. Нередко перышки остаются еще закрученными, когда большая часть листа уже расправилась. Кроме этой общей защиты, почти универсальной для папоротников (за исключением ужовниковых), имеются также более специальные формы защиты, обеспечиваемые различными типами покрывальца, или индузия (от лат. indusium - верхняя туника). Термин этот служит для обозначения "покрывалец" самого различного происхождения. Покрывальца имеются далеко не всегда. Простые сорусы с характерным для них одновременным развитием спорангиев нуждаются в покрывальце в меньшей степени, чем два других типа сорусов, у которых развитие спорангиев более растянуто. Поэтому у большинства семейств с простыми сорусами покрывальце совершенно отсутствует. Однако у схизейных отдельные спорангии (сорусов у них нет) защищены загнутыми на нижнюю сторону краями пластинки листа, или же у видов лигодиума (Lygodium) каждый спорангий полностью прикрыт особым кармашковидным выростом поверхностных тканей листа. Имеются и другие формы защиты, в том числе взаимная защита спорангиев в результате их срастания в так называемые синангии (например, у некоторых мараттиевых). Настоящее же покрывальце среди папоротников с простыми сорусами имеется только у матонии (Matonia). Оно очень своеобразное, зонтиковидной формы и возникает из центральной части ложа соруса. По всем данным, оно произошло совершенно независимо от покрывалец градатных и смешанных сорусов, к которым мы теперь перейдем.
Градатные сорусы, характеризующиеся более длительным развитием и более мелкими спорангиями, нуждаются в защите гораздо больше, чем простые сорусы с более крупными и одновременно развивающимися спорангиями. Поэтому в разных линиях эволюции папоротников с градатными сорусами независимо и разными путями возникли покрывальца, эффективно защищающие развивающиеся спорангии. Покрывальца бывают различной формы, чаще всего более или менее чашевидные или двугубые. Приспособительная эволюция покрывалец представляет собой одну из наиболее увлекательных глав сравнительной морфологии папоротников, но в ней еще много неясного. В частности, не всегда понятно исчезновение в процессе эволюции уже имевшегося у предков покрывальца. Что же касается папоротников со смешанными сорусами, то за исключением большинства даваллиевых (Davalliaceae) они лишены покрывальца. Функцию покрывальца часто исполняют у них перемешанные со спорангиями волоски (парафизы).
Вернемся снова к спорангию. При созревании он раскрывается, а споры освобождаются и рассеиваются. Нас интересуют, однако, те приспособления, которые способствуют раскрыванию спорангия. Начнем с самых древних, вымерших форм. У некоторых из них спорангий при созревании открывался наверху небольшим отверстием. У них, так же как у современных ужовниковых (обладающих верхушечным щелевидным отверстием), нет еще никаких дополнительных приспособлений, способствующих раскрыванию зрелого спорангия. Но уже у целого ряда древнейших папоротниковидных, например у представителей палеозойского семейства зигоптерисовых (Zygopteridaceae), спорангии снабжены с обеих сторон широкой продольной (вертикальной) полоской из нескольких рядов клеток с утолщенными стенками, служившей для раскрывания спорангиев. Это чисто механическое приспособление называют кольцом (рис. 91). Мы не знаем, как действовало у вымерших форм кольцо, но его механизм у современных папоротников хорошо изучен.
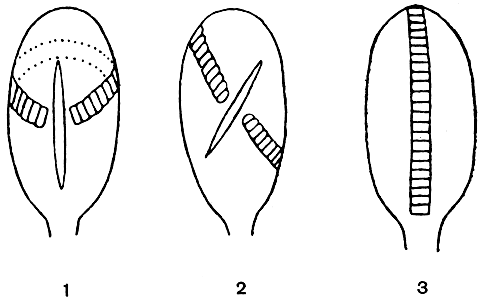
Рис. 91. Основные типы расположения колец у спорангиев папоротников: 1 - поперечное кольцо; 2 - косое кольцо; 3 -продольное кольцо
Познакомимся поэтому со строением и действием наиболее распространенного типа кольца ныне живущих папоротников. Это продольное (вертикальное) образование, состоящее из одного ряда клеток. У большей части клеток кольца три внутренние стенки (две радиальные и внутренняя тангентальная стенка) утолщены, в то время как остальные три (внешние тангентальные стенки) тонкие. Одним своим концом эта часть кольца с утолщенными стенками клеток прочно прикреплена к ножке спорангия. Часть клеток кольца (четверть или треть от общего их количества) имеет относительно тонкие стенки, включая группу из четырех поперечно удлиненных сестринских клеток, образующих так называемый стомий (от греч. stoma - рот). По мере созревания спорангия клетки кольца в результате испарения постепенно теряют воду. Но дело в том, что имеется очень сильное "сцепление" между стенками клеток кольца и водой. Поэтому, когда через тонкие внешние тангентальные стенки вода испаряется, эти стенки благодаря силам сцепления постепенно втягиваются внутрь и становятся все более вогнутыми, в то время как концы радиальных клеток стягиваются друг к другу. По мере того как стенки клеток кольца деформируются, возникает очень сильное механическое напряжение, равное давлению около 300 или более атмосфер. В результате возрастающего напряжения в конце концов наступает момент, когда стенка спорангия разрывается в районе стомия, а само кольцо медленно загибается в обратную сторону, как бы выворачиваясь наизнанку. По мере того как вода продолжает испаряться, возрастающее напряжение деформированных стенок преодолевает силы сцепления и внезапно и одновременно во многих клетках кольца образуется вакуум. Силы сцепления таким образом уже перестают действовать, и кольцо быстро и с силой возвращается к своему исходному положению. В этот момент оно действует подобно катапульте и с силой выбрасывает освободившиеся уже споры. Таким образом, перед нами удивительно остроумный механизм, который, несомненно, в сильнейшей степени способствует широкому распространению папоротников.
Описанного выше типа кольца из однорядных клеток, действующего подобно катапульте, еще не было у вымерших папоротниковидных. Его нет также у ряда современных папоротников. Более того, некоторые современные папоротники вообще не имеют никакого кольца и, как уже было сказано выше, оно совершенно отсутствует у семейства ужовниковых.
У тех родов семейства мараттиевых, спорангии которых еще не срослись в синангии (как у ангиоптериса - Angiopteris), имеется примитивное кольцо, напоминающее кольца некоторых палеозойских папоротниковидных. Однако оно не имеет формы кольца, а представляет собой полоску из несколько более крупных клеток с утолщенными стенками, расположенную на конце продольной линии раскрывания спорангия и более или менее поперечно к ней (поперечное кольцо). Конечно, такое кольцо имеет мало общего с типичным кольцом, о механизме действия которого мы рассказали выше. Оно служит для раскрывания спорангия, но эффект катапульты здесь отсутствует. Столь же примитивно кольцо осмундовых, представляющее собой группу толстостенных клеток, расположенную на одной стороне спорангия близ его верхушки.
Если мы теперь обратимся к схизейным, то найдем у них очень хорошо развитое кольцо, заметно отличающееся от подобного образования у остальных современных папоротников. Оно состоит обычно из одного ряда утолщенных клеток, расположенного непосредственно вокруг верхушки спорангия. Сама верхушка спорангия состоит из одной или нескольких клеток, из чего следует, что кольцо не строго верхушечное. Это, в сущности, поперечное кольцо, расположенное вокруг верхушки. Кольцо способствует продольному раскрыванию спорангия, но опять-таки без катапультирования. Интересно, что у палеозойских зигоптерисовых кольцо спорангиев было вертикальным, а не поперечным. Из этого можно сделать вывод, что схизейные произошли скорее от их общих предков с зигоптерисовыми, чем непосредственно от зигоптерисовых, и что кольцо у них возникло независимо. Папоротники с аппаратом раскрывания спорангиев типа современных схизейных известны уже с карбонового времени. У адиантовых, которые, по современным представлениям, произошли от предков современных схизейных, кольцо приобрело продольное (вертикальное) положение.
Эволюция кольца спорангия других современных семейств папоротников начинается с косого кольца, которое, следовательно, занимает промежуточное положение между поперечным и продольным (вертикальным). Такое косое кольцо характерно для довольно древних семейств глейхениевых и матониевых. У диптериса мы уже наблюдаем более или менее продольное кольцо, а у полиподиевых кольцо уже вполне продольное и раскрывание спорангия поперечное.
Если, наконец, мы перейдем к третьему большому направлению эволюции современных семейств папоротников, то здесь картина окажется сходной. У циатейных кольцо еще более или менее косое (иногда почти продольное). Косое или иногда почти поперечное кольцо наблюдается также в очень близком семействе гименофилловых. У более подвинутых в эволюционном отношении асплениевых и даваллиевых кольцо приобрело продольное положение и спорангии у них раскрываются поперечной щелью.
Таким образом, эволюция кольца во всех трех ветвях родословного древа папоротниковидных заканчивается наиболее прогрессивным продольным (вертикальным) типом. Преимущество продольного кольца, вероятно, состоит в том, что один его конец прочно прикреплен к ножке спорангия, которая служит надежной опорой для движений кольца назад и возвращения в исходное положение. Сходство с продольным кольцом некоторых палеозойских папоротников не очень большое, так как у последних они состояли из нескольких (до 8) рядов клеток и, вероятно, функционировали несколько иначе. Продольное кольцо возникало в процессе эволюции независимо несколько раз и в настоящее время является наиболее распространенным типом.
А теперь попытаемся выяснить, в чем именно заключается биологическое значение перехода от поперечного кольца к косому, а этого последнего к продольному. Исследования английского птеридолога (специалиста по папоротникам) Ф. Бауэра показали, что эволюция кольца спорангия была тесно связана с эволюцией всего соруса.
Раскрывание спорангиев с поперечным кольцом происходит в продольном направлении. Продольное раскрывание мы наблюдаем у осмундовых, схизейных и глейхениевых, т. е. у семейств с относительно еще крупными спорангиями, которые или еще не собраны в сорусы, или образуют примитивные сорусы, как у глейхениевых. На этой стадии эволюции продольное раскрывание достаточно эффективно. Но с образованием градатных и особенно смешанных сорусов дело меняется. При большом числе более скученных спорангиев продольное раскрывание становится неудобным по чисто механическим причинам и поэтому сменяется боковым (сначала косым, а затем поперечным).
Косое раскрывание характерно для градатных сорусов. Удлиненное ложе градатных сорусов облегчает боковое (в данном случае косое) разверзание спорангиев, так как здесь все спорангии раскрываются наружу и тем самым отсутствуют физические препятствия для рассеивания спор. Это хорошо видно у циатейных и особенно у гименофилловых. Косое раскрывание спорангиев является в этом случае достаточно эффективным. Однако ситуация резко меняется с переходом градатных сорусов в смешанные. В смешанных сорусах (где более многочисленные мелкие спорангии тесно скучены, а ложе обычно плоское) непрерывное (полное) косое кольцо переходит в более или менее прерванное в месте прикрепления ножки неполное продольное кольцо и раскрывание тем самым становится поперечным, как это хорошо видно, например, у щитовника (Dryopteris). Так как длинная ножка зрелого спорангия приподнимает его над окружающими более молодыми спорангиями, то продольное кольцо может без всяких физических помех полностью закрутиться на обратную сторону и вновь с си-лой вернуться в исходное положение, разбрасывая таким образом споры. В сорусах промежуточного типа между градатными и смешанными или между простыми и смешанными наблюдается промежуточный тип кольца и промежуточный способ раскрывания спорангия.
В процессе эволюции спорангия и всего соруса и совершенствования механизма его раскрывания число спор в каждом спорангии постепенно уменьшается. Важно подчеркнуть, что речь идет об уменьшении количества спор, производимого не спорофитом в целом, а лишь каждым отдельным спорангием. Число спор, производимых спорофитом, не уменьшается (оно может даже увеличиваться), так как в ходе эволюции равноспоровых папоротников количество спорангиев имеет определенную тенденцию возрастать.
В примитивных, крупных спорангиях количество спор еще довольно велико. Так, у некоторых ужовниковых насчитывают до 15 000 спор (обычно 1500-2000), а у мараттиевых хотя и меньшее количество спор, но тоже достаточно большое - от 1450 до 7500. Более значительное уменьшение количества спор мы наблюдаем у глейхениевых (1024, 512, 256, 128), осмундовых (от 512 до 128), гименофилловых (512, 256, 128, а иногда 64 и даже 32) и схизей-ных (256, чаще 128, иногда всего 64). У всех остальных папоротников лишь редко 128 спор (Cheiropleuria), обычно 64 (чаще всего), 48 или 32 споры, редко 16 и еще реже 8 спор. У разноспоровых папоротников число микроспор 64. Из мегаспор полного развития достигает только одна.
Как мы уже знаем из вводной главы этого тома, в спорах содержится вдвое меньшее число хромосом, чем в клетках спорофита. Мы знаем также, что это уменьшение числа хромосом происходит в процессе деления материнской клетки споры. Из каждой материнской клетки образуются четыре споры, или тетрада спор (рис. 92). На первых стадиях развития эти четыре споры еще тесно связаны между собой своей внутренней стороной, или так называемым проксимальным (от лат. proximus - ближайший) полюсом. Противоположный, обращенный кнаружи полюс называют дистальным (от лат. distalis - дальше отстоящий). Когда тетрада распадается, то каждая отдельная спора на своем проксимальном полюсе несет след своей связи с соседними спорами тетрады - тетрадный рубец, на котором расположена щель. Через эту щель на ранних стадиях развития тетрады осуществляется цитоплазматический обмен между спорами. Затем после расхождения спор щель является единственным местом контакта цитоплазмы споры с внешней средой (ведь спора живет и ей необходимо дыхание) и наконец через щель спора прорастает.
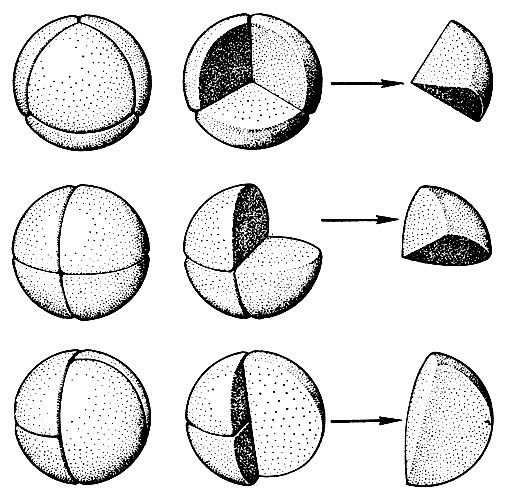
Рис. 92. Три основных типа тетрад и расположение в них трилетных (верхний ряд) и монолетных (средний и нижний ряды) спор
У папоротников имеются два типа тетрадного рубца - трехлучевой и однолучевой. Споры с трехлучевым тетрадным рубцом называют трехрубцовыми или трилетными. Споры с однолучевым тетрадным рубцом называют однорубцовыми или монолетными. Эти два типа спор различаются также по своей геометрической форме. Трилетные споры характеризуются радиальной симметрией и имеют форму более или менее шарообразных тетраэдров, и для простоты их называют тетраэдрическими. Монолетные споры имеют только две плоскости симметрии. Они более или менее билатеральные - бобовидные или эллипсоидальные. От чего же зависят форма споры и тип ее тетрадного рубца? Как показали специальные исследования (Ф. Бауэр, Н. Маренго, А. Н. Сладков), форма споры и тип ее тетрадного рубца определяются на стадии тетрады. Деление материнской клетки споры происходит разными способами. В случае образования трилетных спор материнская клетка дает начало четырем тетраэдрически симметричным спорам, соприкасающимся одна с другой гранями пирамидальной проксимальной стороны. Каждая такая спора соприкасается с тремя другими спорами, и на ее проксимальном полюсе образуются три радиально расходящихся рубца, составляющие вместе один трехлучевой рубец.
При образовании монолетных спор материнская клетка делится на четыре удлиненные билатерально-симметричные "четверти сферы", которые на стадии тетрады соприкасаются гранями клиновидной проксимальной стороны. Такие билатеральные споры могут образоваться в тетрадах двух типов - изобилатеральных и крестообразных. В изобилатеральных тетрадах длинные оси всех четырех спор расположены параллельно по отношению друг к другу и таким образом каждая спора соприкасается лишь с двумя соседними спорами. В крестообразных тетрадах споры образуют две пары, причем длинная ось одной пары образует прямой угол с длинной осью другой пары. В таких тетрадах каждая спора соприкасается со всеми остальными, причем одной половиной проксимальной стороны с одной спорой, а другой половиной - с двумя. В результате у споры образуется простой однолучевой рубец.
Билатеральные монолетные споры произошли из тетраэдрических трилетных спор, причем это превращение трилетных спор в монолетные произошло независимо в разных линиях эволюции. Монолетные споры появляются уже у некоторых относительно примитивных семейств. Они встречаются у мараттиевых (мараттия - Marattia, данея - Danaea), схизейных (схизея - Schizaea), некоторых глейхениевых и циатейных, у адиантовых (около половины представителей подсемейства виттариевых - Vittarioideae), а также характерны для древнего рода диптерис. Такие же высокоподвинутые семейства, как многоножковые и асплениевые, характеризуются уже монолетными спорами.
Если монолетные споры произошли из трилетных, то это, очевидно, связано с тем, что в процессе эволюции исходный тетраэдрический тип тетрады дал начало двум другим типам - изобилатеральному и крестообразному. В обоих случаях возникает монолетная спора, но, как показал А. Н. Сладков (1962, 1970) на основе моделирования спор и тетрад, это происходит двумя разными способами. В случае превращения тетраэдрических тетрад в изобилатеральные монолетная билатеральная спора возникает путем редукции и исчезновения двух лучей первоначального трехлучевого тетрадного рубца. Когда тетраэдрическая тетрада превращается в крестообразную, монолетная спора образуется за счет редукции и исчезновения одного луча трехлучевой тетрады трилетных спор и увеличения угла между оставшимися лучами до 180°.
В чем заключается биологическое преимущество монолетных спор по сравнению с трилетными? К сожалению, на этот вопрос можно ответить только догадками. В случае монолетных спор деление материнской клетки происходит более простым способом и достигается путем образования меньшего числа перегородок. Возможно, это обстоятельство играло определенную роль в переходе тетраэдрических спор в билатеральные. Но было, вероятно, не менее важным изменение формы споры и укрепление ее рубца. Не лишено вероятности, что рассеивание и прорастание монолетных спор происходит легче, чем трилетных. Однако для выяснения этого вопроса нужны специальные исследования.
Переход трилетного типа в монолетный происходит без большого труда. Это видно из того, что некоторые папоротники, нормально производящие трилетные споры, могут наряду с последними производить также некоторое количество монолетных спор, что наблюдалось, например, у ужовника (Ophioglossum), гроздовника, ангиоптериса и птериса (Pteris). Но нередко наблюдается и обратное - наличие некоторого количества трилетных спор у папоротников, нормально образующих только монолетные споры. Это можно наблюдать у мараттии и схизеи вильчатой (Schizaea dichotoma) или у щитовника пильчатого (Dryopteris serrata). Более того, в ряде случаев наблюдаются промежуточные формы между трилетными и монолетными спорами. Так, например, у диптериса китайского (Dipteris chinensis) и птериса критского (Pteris cretica) можно видеть сильное удлинение одного из трех лучей тетрадного рубца при одновременном укорочении, а затем и исчезновении двух других лучей. При наблюдении таких промежуточных спор возникает мысль, что их образование обязано промежуточным типам тетрад. И действительно, по исследованиям А. Н. Сладкова (1962), моделирование промежуточных форм спор и составление из таких моделей тетрад показали, что тетрады также получаются промежуточные, причем двух типов: переходные от тетраэдрических к изобилатеральным и переходные от тетраэдрических к крестообразным.
Из двух основных слоев оболочки спор - интины (эндоспория) и экзины (экзоспория) - наибольший интерес представляет последний. Экзина интересна тем, что, как и у подавляющего большинства других высших растений, она отличается большим разнообразием строения. У разных групп папоротников она различна по своей толщине и, что гораздо важнее, по своей структуре. При исследовании спор папоротников с помощью сканирующего электронного микроскопа споры большинства папоротников обнаруживают поразительное разнообразие экзины. На таблицах 22, 24, 27 и 28 показаны некоторые из наиболее интересных спор. Строение экзины, особенно когда ее изучают с помощью электронного микроскопа, все шире используют для решения вопросов систематики и эволюции папоротников, как ныне живущих, так и вымерших.

Таблица 24. Фегоптерис связывающий (Phegopteris connectilis): 1 - часть листа с сорусами; 2, 3 - спорангий в разных положениях (увел, около 300); 4 - спора (увел, около 2000); 5 - деталь поверхности споры (увел, около 4500); (2-5 - под сканирующим электронным микроскопом)
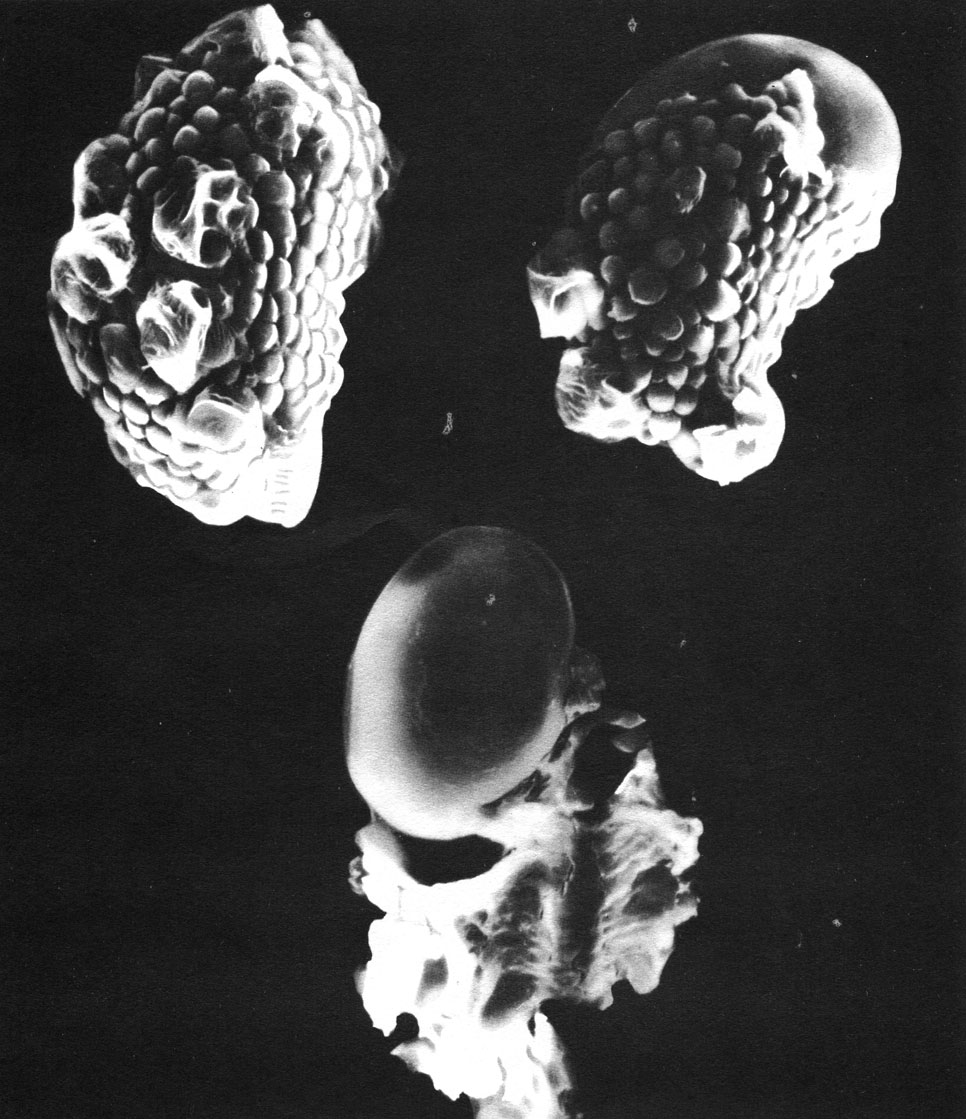
Таблица 28. Споры пиррозии прирастающей (Pyrrosia adnascens) под сканирующим электронным микроскопом (увел, около 1300): вверху слева - спора, поверхность которой образована периной (периспорием); вверху справа - перина частично сползла и на поверхности оказалась эктэкзина (экзоспорий); внизу - спора со сброшенной периной
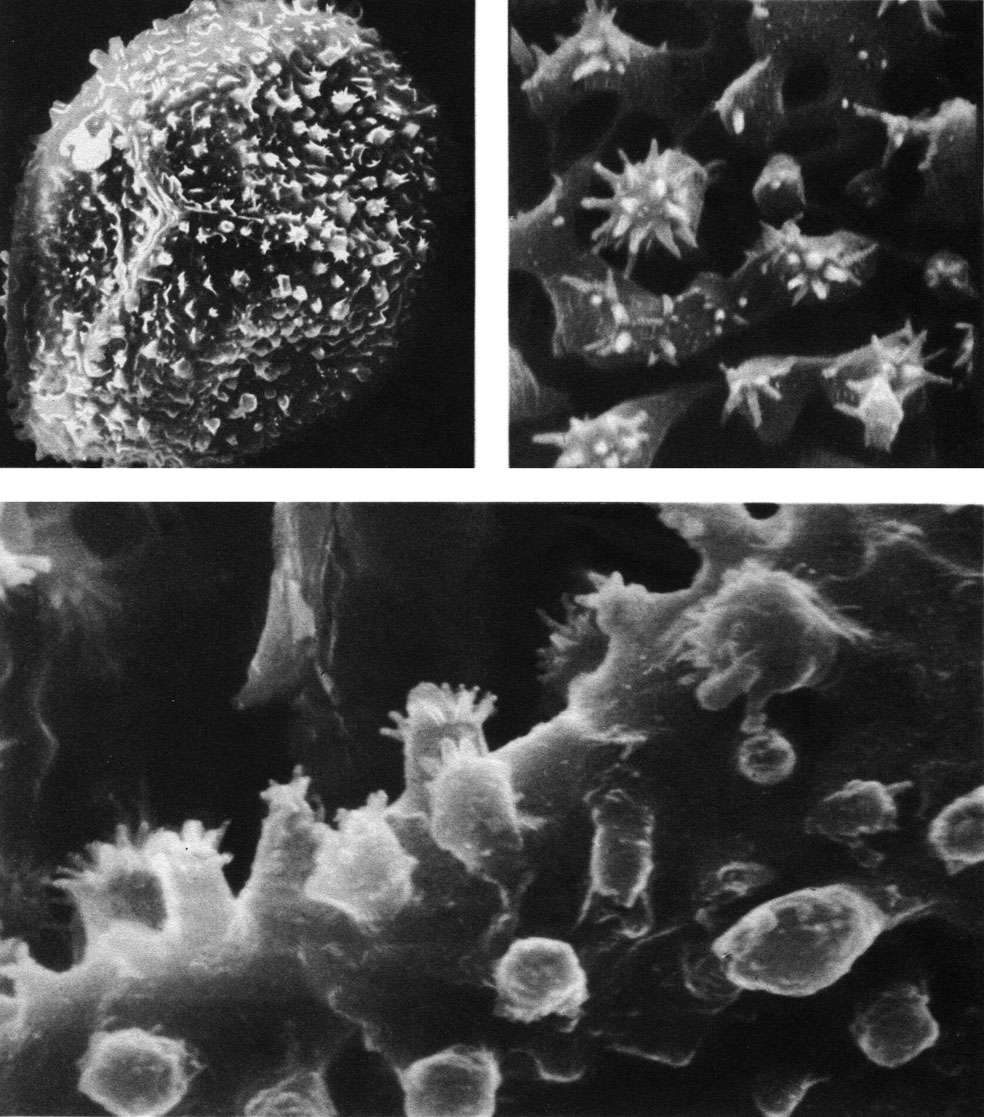
Таблица 27. Спора осмунды коричной (Osmunda cinnamomea) под сканирующем электронным микроскопом: вверху слева - увел. 4000, вверху справа - увел. около 25 000; внизу - увел. около 25 000
Споры большинства папоротников (особенно асплениевых) имеют также дополнительную наружную оболочку, откладываемую на поверхности уже сформировавшейся экзины так называемым плазмодием. Плазмодий (или периплазмодий) образуется в результате разрушения клеток тапетума (выстилающего слоя спорангия). У более примитивных папоротников с многослойными стенками спорангия тапетум образуется из клеток самого внутреннего слоя стенки, а у папоротников с однослойной стенкой он образуется из самой спорогенной ткани. Плазмодий служит питательной средой для развивающихся спор. В него споры погружены во время созревания. Так образуется дополнительный слой оболочки споры - перина (периспорий). Перина обычно резко отличается от экзины, бывает неплотно с ней соединена и более или менее легко разрушается и отпадает. В ископаемом состоянии она не сохраняется. Перина может быть гладкой, сетчатой, шиповатой и т. д. и в некоторых случаях достигает значительной сложности. Поэтому она также может иметь таксономическое значение. Наличие такого рода перины рассматривается как признак специализации. Как показали электронномикроскопические исследования французского палинолога Б. Люгардона (1974), перина имеется не только у асплениевых. Оказалось, что и у остальных папоротников имеется перина, но она образует столь тонкий слой, что не может быть обнаружена с помощью светового микроскопа.
Любопытной особенностью спор некоторых папоротников является наличие в них хлоропластов. У большинства папоротников споры содержат только пропластиды и поэтому они желтые или коричневые (редко почти черные), однако споры осмундовых, гименофилловых и граммитисовых и некоторых асплениевых (оноклея и страусник) содержат значительное число хлоропластов и имеют поэтому зеленый цвет. Биологическое значение хлоропластов в спорах не вполне ясно. Но приуроченность спор с хлоропластами к характерным эпифитам тропических лесов (гименофилловые, граммитисовые) позволяет предполагать, что наличие хлорофилла способствует быстрому прорастанию спор, что, несомненно, облегчает расселение. У обитателей умеренных зон (оноклея, страусник, осмундовые) зеленые споры продуцируются только весной, когда выпадает наибольшее количество дождей. В обоих случаях связь с избыточной влажностью очевидна. Зеленые споры прорастают сразу, без периода покоя.
Наше знакомство со спорами папоротников было бы неполным, если бы мы не рассмотрели вопрос о происхождении разноспоровости. Разноспоровость, т. е. дифференциация спор на микро- и мегаспоры (иногда их называют мужскими и женскими спорами), возникала в процессе эволюции, несколько раз. Разноспоровые группы известны как среди вымерших, так и среди ныне живущих папоротников. У одного из современных папоротников, а именно у монотипного северо-восточноавстралийского рода платизома (Platyzoma), составляющего отдельное семейство платизомовых (Platyzomataceae), уже давно была обнаружена зачинающаяся разноспоровость, что полностью подтверждается новейшими исследованиями. У этого папоротника одни спорангии крупнее, а другие мельче. В первых содержится до 16 относитель-но крупных спор, а во вторых - до 32 мелких спор. Мелкие споры при прорастании дают начало гаметофиту, на котором развиваются только антеридии. Из крупных спор развиваются гаметофиты, на которых в молодости образуются антеридии, а позже только архегонии. Таким образом, если мелкие споры можно назвать микроспорами, то более крупные споры не являются еще настоящими мегаспорами. Разноспоровость у платизомы еще не полная, не вполне установившаяся. Но среди современных папоротников есть три семейства, у которых разноспоровость достигла полного развития. Это - марсилеевые (Marsileaceae), сальвиниевые (Salviniaceae) и азолловые (Azollaceae).
Биологическое преимущество разноспоровости перед равноспоровостью заключается в том, что гаметофит развивается внутри споры (становится эндоспорическим) и развивается за счет тех питательных веществ, которые содержатся в споре, особенно обильно - в мегаспоре. Благодаря быстрому развитию сильно редуцированного паразитического гаметофита процесс оплодотворения происходит рано, гораздо раньше, чем у равноспоровых папоротников. Обильная пища, накопленная в мегаспоре, создает также максимально благоприятные условия для развивающегося зародыша.
Биологические преимущества, достигнутые в результате разноспоровости, вероятно, важны в любых условиях среды, но особенно важны в крайних или меняющихся условиях существования. Марсилеевым, сальвиниевым и азолловым биологические преимущества разноспоровости дают возможность существовать в условиях высыхающих время от времени болот и прудов. Чередующиеся дожди и образование временных водоемов дает им возможность быстро завершить весь репродуктивный цикл.
Таковы основные морфологические и биологические особенности спор. В последние десятилетия всестороннему изучению спор папоротников, как и спор вообще, уделяется все большее внимание. Ведь производство спор есть завершающий этап в развитии спорофита. Все структурные и функциональные изменения спорофита, происходящие в ходе эволюции, связаны с необходимостью размножения посредством спор. "Победителями в борьбе за существование" оказываются те, у кого достигается наибольшая эффективность в производстве, рассеивании и прорастании спор.
В заключение этого раздела нужно сказать несколько слов о так называемой апоспории - чрезвычайно интересном в биологическом отношении явлении, когда гаметофит образуется не из споры, а из вегетативной клетки или клеток спорофита. Вполне понятно, что такой гаметофит будет иметь одинаковое с клетками спорофита число хромосом. В природе апоспория встречается не часто, но ее обычно бывает нетрудно вызвать искусственно. Легче всего это удается, если отрезанные листья молодого спорофита поместить на агаровую среду. Через некоторое время клетки листа, соприкасающиеся с агаром, дадут начало нитям, которые в дальнейшем превращаются в типичный (но диплоидный) гаметофит.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'