Корень, стебель и лист
Как и у большинства высших растений, спорофит папоротников снабжен корнями, отсутствующими только у части гименофилловых (Hymenophyllaceae) и у рода сальвиния (Salvinia), что представляет собой результат редукции. Корни папоротников придаточные. Это значит, что первичный корень (корень зародыша) не получает дальнейшего развития и вскоре отмирает и вместо него развиваются корни из стебля, а в некоторых случаях также из оснований листьев. Корни обычно волокнистые, но у ужовниковых (Ophioglossaceae) и некоторых других примитивных групп имеют тенденцию к мясистости. Ветвление корней, как правило, моноподиальное.
Стебли папоротников никогда не бывают так сильно развиты и не достигают таких размеров, как у хвойных или у древесных двудольных. У папоротников, как правило, листва по массе и размерам преобладает над стеблем. Тем не менее стебли папоротников довольно разнообразны как по внешней своей форме, так и особенно по внутреннему, анатомическому строению. Прямостоячий стебель древовидных папоротников, несущий на верхушке крону листьев, называют стволом. Высокие стволы обычно снабжены у основания многочисленными воздушными корнями, придающими им устойчивость. Когда стебель ползучий или вьющийся, его называют корневищем. Корневище может быть довольно длинным (у вьющихся форм) или, наоборот, очень коротким и клубневидным. Оно бывает или радиаль-ным, и тогда листья и корни покрывают его равномерно со всех сторон, или дорсивентральным (спинно-брюшным), и тогда листья сидят на его верхней (спинной) стороне (часто в двух или иногда более чередующихся рядах), а корни только или главным образом на нижней (брюшной) стороне. Внутреннее строение соответствует внешней морфологии, т. е. бывает радиальным или дорсивентральным.
Стебли папоротников нередко ветвятся. Иногда ветвление бывает дихотомическим, но чаще ветви возникают в местах отхождения листьев, обычно перед листовыми основаниями.
Для классификации папоротников имеют значение не только форма и размеры стебля, но и строение тех волосков или чешуй, которыми они бывают почти всегда покрыты в молодости и реже также во взрослом состоянии (аналогичные волоски или чешуйки покрывают также листья, особенно еще не развернувшиеся). Волоски состоят из одной клетки или же одного ряда клеток и у разных папоротников имеют разную длину и толщину. В некоторых случаях встречаются также щетинки, которые отличаются от волосков тем, что у основания они толщиной более чем в одну клетку. Верхушечная клетка волоска нередко превращается в железку, как, например, у некоторых видов хейлантеса (Cheilanthes). Часто верхушечная клетка волоска выделяет слизь, защищающую молодые части, как у осмундовых (Osmundaceae), или у блехнума (Blechnum). Простые волоски особенно характерны для таких относительно примитивных групп, как ужовниковые, осмундовые, матониевые (Matoniaceae), некоторые циатейные (Cyatheaceae). У некоторых папоротников базальная клетка волоска подвергается продольному делению, в результате чего образуется жесткая щетинка, как у диптериса (Dipteris). Волоски могут также ветвиться. В тех случаях, когда ветви остаются свободными, могут образовываться различного рода звездчатые волоски. Когда эти ветви срастаются, образуются чешуи, представляющие собой пластинки толщиной в одну клетку. Детали строения чешуй имеют большое таксономическое значение, и поэтому в работах по систематике папоротников микроскопическому строению чешуй уделяется большое внимание. Из наиболее интересных типов чешуй отметим щитовидные (пельтатные) и решетчатые (клатратные). Щитовидные чешуи прикреплены одной из точек своей поверхности, а не краем пластинки, как обычно. Решетчатые чешуи характеризуются тем, что боковые стенки клеток утолщены, образуя явственный решетчатый узор.
Стебли древнейших девонских папоротниковидных мало отличались от стеблей риниофитов, и их проводящая система представляла собой очень примитивную протостелу (рис. 13). Протостела имеется и у некоторых современных папоротников, например у схизейных (Schizaeaceae) и гименофилловых и у многих глейхениевых (Gleicheiniaceae). Но у большинства современных папоротников проводящая система представляет собой различные формы сифоностелы, которая достигла у них большого развития. В зависимости от характера взаимного расположения флоэмы и ксилемы различают два типа сифоностелы: эктофлойную сифоностелу, у которой флоэма окружает ксилему только снаружи, и амфифлойную сифоностелу, у которой флоэма окружает ксилему с обеих сторон, т. е. как снаружи, так и изнутри.
Сифоностела может представлять собой относительно сплошной цилиндр проводящей ткани, но чаще проводящая система представляет сеть проводящих пучков. В последнем случае на поперечном срезе она имеет вид кольца из отдельных пучков. Промежутки между отдельными пучками не являются пустыми, но заполнены паренхимной тканью. Паренхимные участки, расположенные в стеле над местами отхождения листовых следов, называют листовыми прорывами или лакунами. Амфифлойную сифоностелу, в которой последовательные листовые прорывы значительно отделены друг от друга, называют соленостелой (от греч. solen - канал). Когда в амфифлойной сифоностеле листовые прорывы расположены столь близко, что нижняя часть одного прорыва параллельна верхней части другого, ее называют диктиостелой (от греч. diktyon - сеть). Диктиостела представляет собой цилиндр, состоящий из сети переплетающихся пучков. Отдельный пучок диктиостелы называют меристелой (от греч. meros - часть). Соленостела и диктиостела являются двумя наиболее распространенными типами стелы среди современных папоротников. В эволюционном отношении диктиостела наиболее подвинутый тип, и поэтому она характерна для наиболее прогрессивных семейств.
Листья папоротников, часто называемые (особенно в старой литературе) вайями, во многих отношениях отличаются от листьев хвощевидных и особенно плауновидных. В то время как листовые органы плауновидных представляют собой просто выросты на надземных осевых органах, а листья хвощевидных являются видоизмененными боковыми веточками, у папоротников листья морфологически соответствуют целым крупным ветвям их вероятных предков - риниофитов. Изучая примитивные палеозойские папоротниковидные, палеоботаники не раз обращали внимание на то, что листья у них представляли нечто промежуточное между листьями в обычном понимании и ветвями. У некоторых из них имелись радиально-симметричные листовые органы, которые лишь с большой натяжкой могут быть названы листьями. Одиночное и верхушечное расположение спорангиев у наиболее примитивных папоротниковидных также является убедительным свидетельством веточной природы их листьев. Наконец, об этом же красноречиво говорят такие факты, как характерный для папоротников верхушечный и притом длительный (иногда почти неограниченный) рост их листьев, обычно большие размеры и сложно рассеченная форма листовой пластинки, крупные и сложные листовые следы и наличие листовых прорывов (лакун) в стеле. На основании всех этих фактов делается вывод, что листья папоротников произошли из ветвей в результате их уплощения и ограничения в росте. Радиально-симметричная ветвь постепенно превращается в "плосковетку". По мере уплощения происходит также дифференциация верхней (спинной) и нижней (брюшной) сторон листа. Он становится дорсивентральным. Другим важным результатом уплощения является большее или меньшее срастание отдельных веточек листа, его сегментов. В результате возникает более совершенный тип листа, который гораздо лучше приспособлен для максимального использования света в процессе фотосинтеза.
Листья папоротников отличаются очень большим разнообразием. По своим размерам они колеблются от нескольких миллиметров до 30 м и более в длину и не менее разнообразны по внешней форме и внутреннему строению. В большинстве случаев листья папоротников совмещают обе функции - фотосинтез и спороношение. Но у многих папоротников, например у страусника (Matteuccia struthiopteris), оноклеи чувствительной (Onoclea sensibilis) или у эпифитного тропического рода дринария (Drynaria), листья дифференцированы на стерильные (фотосинтезирующие) и фертильные (несущие спорангии). В крайних случаях диморфизма листьев фертильные листья теряют хлорофилл, и тогда их функция сводится к спороношению, как у сальвинии.
В большинстве случаев листья более или менее явственно дифференцированы на черешок и пластинку. У некоторых папоротников, например у большинства представителей большого семейства полиподиевых (Polypodiaceae), черешки соединены с корневищем посредством особого сочленения, но чаще сочленение отсутствует. Биологическое значение сочленения заключается, вероятно, в том, что оно дает возможность листу более свободно менять свою ориентацию по отношению к меняющемуся направлению падающих на него солнечных лучей. Этот признак важен также при различении групп папоротников. Так, семейства полиподиевых и граммитисовых (Grammitidaceae) различаются, помимо других признаков, тем, что у первого черешок обычно с сочленением, а у второго - без сочленения. Поэтому же признаку можно различать и роды внутри одного семейства.
Для систематики папоротников имеет значение также строение проводящей системы черешка, в частности форма проводящего пучка на поперечном срезе. Например, у осмундовых проводящий пучок черешка имеет на поперечном срезе С-образную форму, а у близких к ним плагиогириевых (Plagiogyriaceae) форма более или менее Υ-образная. Очень важно и число проводящих пучков: 1, 2 или несколько. У некоторых асплениевых (Aspleniaceae) в основании черешка обычно 3-7 проводящих пучков, но у большинства - только 2 пучка, соединяющихся наверху в один.
Интересно отметить, что примитивные девонские папоротниковидные, которые еще не имели типичного папоротникового листа, обладали черешком с радиальной симметрией, по своей внешней форме и внутреннему строению мало отличавшимся от стебля.
Примитивные листья имеют дихотомическое ветвление, причем равнодихотомическое ветвление эволюционно предшествует неравнодихотомическому. Древний, дихотомический тип ветвления листовой пластинки сохранился и среди современных папоротников. При этом он сохранился не только у ряда представителей такого относительно примитивного семейства, как схизейные, но и у представителей гораздо более развитых семейств, как, например, у довольно специализированного ксерофильного папоротника актиниоптериса южного (Actiniopteris australis), относящегося к адиантовым (Adiantaceae). У подавляющего же большинства современных папоротников листья перистые - однажды, дважды или многократно. Пластинка перистого листа, в отличие от дихотомического, имеет стержень, или рахис (от греч. rhachis - позвоночник), представляющий собой продолжение черешка (рис. 90). Стержень соответствует главной жилке цельного листа. Если лист однажды перистый, стержень несет по бокам по одному ряду сегментов, называемых перьями.
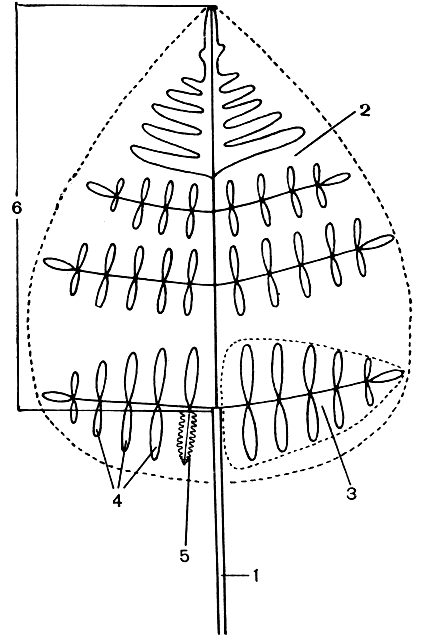
Рис. 90. Строение листа папоротника (схема): 1 - черешок; 2 - пластинка листа; 3 - перо первого порядка; 4 - перышки (или перья второго порядка); 5 - лопасть перышка; 6 - рахис
Перья могут быть цельными или лопастными. Когда лист дважды перистый, стержень листа (в данном случае он будет главным стержнем) несет по бокам вторичные (боковые) стержни, которые, в свою очередь, усажены сегментами (перьями) второго порядка, называемыми перышками. Все сегменты последнего порядка, будь то второго, третьего или последующих порядков, мы называем перышками. Так, если лист трижды перистый, то у него будут перья первого и второго порядков и перышки (перья третьего порядка). У дважды и многократно перистых листьев наряду с главным стержнем имеются стержни второго, третьего и последующих порядков. Отметим, что, в отличие от главного черешка всего листа, черешки перьев второго и последующих порядков называют черешочками. Конечно, перышки часто бывают сидячими, и тогда они, следовательно, лишены черешочка.
Пластинка листа пронизана более или менее сложно разветвленной системой проводящих пучков, которая внешне обычно хорошо выражена в виде так называемого жилкования. Жилка есть не что иное, как выступ ткани листа над проходящим внутри пучком. Жилкование листьев довольно разнообразно, и его особенности имеют существенное значение для классификации папоротниковидных. Не имея возможности описать здесь все огромное разнообразие типов жилкования в листьях папоротников, мы ограничимся только указанием одного из наиболее важных признаков. Так как папоротниковый лист произошел из дихотомической ветви предковых форм, то вполне естественно, что жилкование у примитивных папоротников должно было быть дихотомическим. Отдельные жилки типичного дихотомического жилкования не образуют еще сети, и такое жилкование называют поэтому открытым.
Открытое дихотомическое жилкование сохранилось и у ряда современных папоротников, причем даже у форм довольно подвинутых в эволюционном отношении. Такое жилкование можно наблюдать, например, у ряда представителей семейства гименофилловых (особенно наглядно у трихоманеса почковидного - Trichomanes reniforme). Но открытое дихотомическое жилкование в процессе эволюции уступает более совершенному с точки зрения эффективности водоснабжения листа сетчатому жилкованию. Оно, конечно, возникло не сразу. Постепенно между отдельными ветвями дихотомической системы жилок возникали перемычки, число которых постепенно возрастало. В результате вся система жилкования превратилась в сеть, состоящую из многочисленных ячеек (называемых ареолами), имеющих у разных таксонов различную форму, размер и расположение. Сеть может состоять только из одного ряда ячеек, расположенных вдоль средней жилки пера или перышка, оставляя все остальные жилки свободными. Но у папоротников с более подвинутым типом жилкования сеть занимает почти всю поверхность пластинки и в конце концов даже конечные веточки жилкования, прежде свободно оканчивавшиеся в крае листа, также соединяются между собой. Все эти стадии эволюции жилкования можно наблюдать как у ископаемых, так и у современных папоротников.
К сожалению, мы не имеем здесь возможности рассказать о разнообразном устьичном аппарате листьев папоротников. Но читатель может получить представление об основных их типах, прочитав еще раз об устьицах (рис. 4 и 5). Отметим только, что у папоротников встречаются почти все основные типы устьичного аппарата (за исключением анизоцитного), в том числе и такие, которые характерны только для папоротников (десмоцитный и перицитный) или главным образом для папоротников (полоцитный). Тип устьичного аппарата у папоротников специфичен для определенных таксонов и имеет поэтому значение для их систематики.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'