Спородерма (Н. Р. Мейер)
Споры, в том числе микро- н мегаспоры, окружены оболочкой, которую в специальной литературе называют спородермой (от греч. spora - сев, семя и derma - кожа).
Спородерма выполняет разнообразные функции, связанные с жизнедеятельностью споры. На ранних стадиях развития оболочка обеспечивает избирательное поступление веществ из полости спорангия (или гнезда пыльника) в развивающуюся спору. Покидая материнское растение, споры некоторое (иногда довольно продолжительное) время существуют самостоятельно. В этих условиях на их оболочку, не прикрытую другими клетками, ложится полная ответственность за сохранение содержимого от высыхания, чрезмерного увлажнения или механических повреждений. Кроме этого, являясь наружным скелетом, спородерма сохраняет определенную и строго постоянную для данного вида форму, которая должна отвечать многим факторам: при наименьшей поверхности заключать максимальный объем, в зависимости от характера опыления хорошо держаться в воздухе, на воде, прикрепляться к насекомым, птицам или другим животным-переносчикам. Чрезвычайно важным свойством спородермы семенных растений является "умение узнавать" особей своего вида, что достигается благодаря содержанию в ней специальных гормонов, стимулирующих прорастание пыльцевой трубки.
Большое количество жизненно важных функций, ложащихся на оболочку всего лишь одной клетки, вызвало необходимость постоянно совершенствовать строение спородермы. В результате выработалась чрезвычайно прочная, устойчивая к высоким температурам, химическим реагентам и воздействию микроорганизмов, совершенная по конструкции, форме и скульптуре оболочка (табл. 3). Особенностью наружных слоев спородермы является также их поистине удивительная сохранность в ископаемом состоянии. О существовании многих давно вымерших растений мы знаем только по остаткам их спородермы.
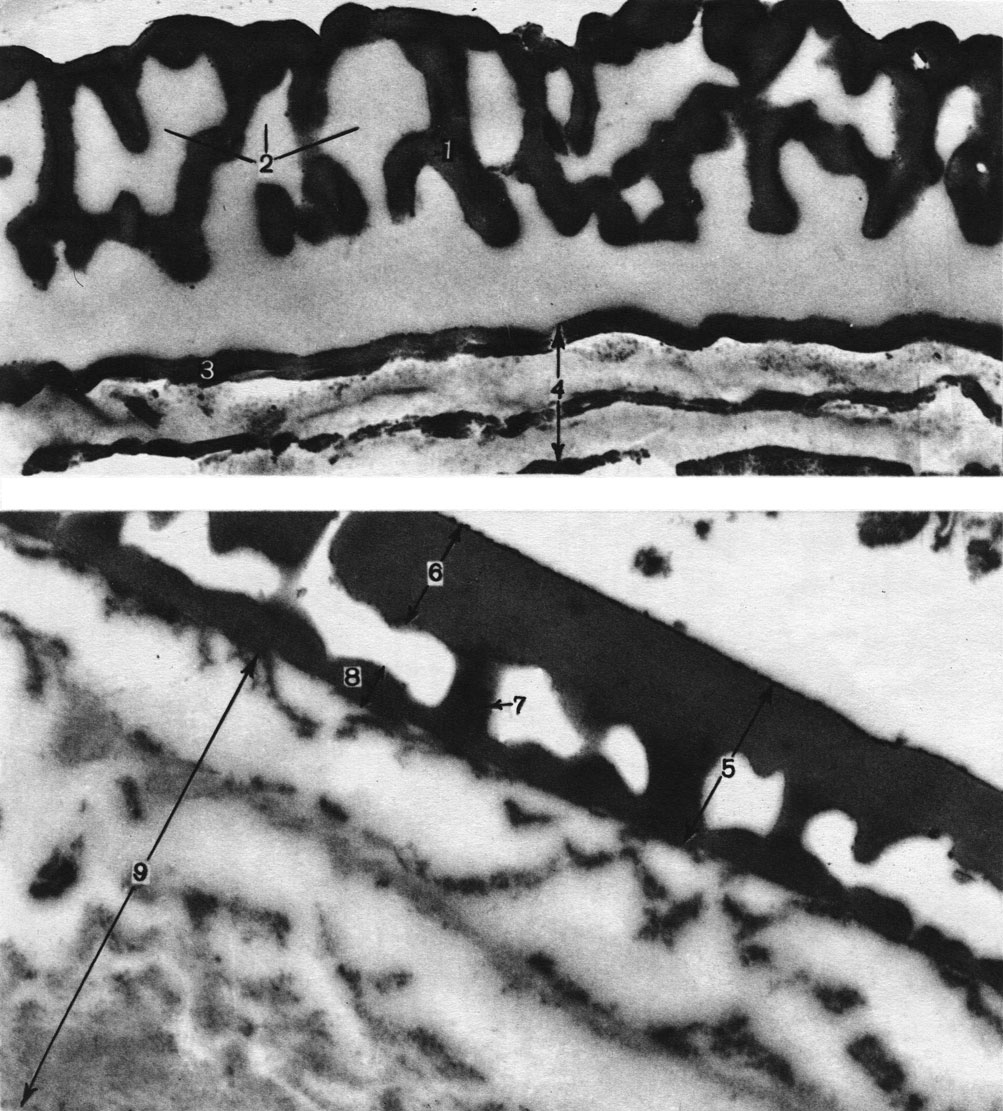
Таблица 3. Спородерма под сканирующим электронным микроскопом (увел, около 20 000): вверху - у сосны черной (Pinus nigra) из голосеменных (1 - ячеистая эктэкзина, 2 - ячеи, 3 - ламелла, 4 - ламеллярная эндэкзина); внизу - у каликантуса фертильного (Calycanthus fertilis) из цветковых (5 - столбиковая эктэкзина, 6 - покров, 7 - столбик, 8 - подстилающий слой, 9 - интина)
Как нам уже известно, каждая материнская клетка в результате мейоза дает 4 споры, которые довольно долго (у некоторых растений постоянно) сохраняются в тетрадах. Благодаря тому, что споры образуются в тетрадах, спородерма характеризуется определенной полярностью. Поверхность спородермы, обращенную к центру тетрады, называют проксимальной (от лат. proximus - ближайший), а противоположную ей поверхность - диcтальной (от лат. distalis - дальше отстоящий). Центр проксимальной поверхности считают дроксимальным полюсом, а центр пистальной поверхности - соответственно дистальным полюсом. Воображаемую прямую линию, соединяющую проксимальный и дистальный полюсы, называют полярной осью. Линию, перпендикулярную полярной оси и проходящую через центр тетрады, принято называть экваториальным диаметром.
Споры с резко различными поверхностями (проксимальной и дистальной) носят название неравнополярных. В отличие от них равнополярными называют споры с отчетливыми полюсами и неразличимыми между собой дистальной и проксимальной поверхностями. Но нередко вся поверхность спородермы имеет одинаковое строение, так что после распадения тетрады невозможно определить местонахождение полюса и экватора.
В спородерме имеются сквозные отверстия или же места, легко растягивающиеся под давлением. Они носят названия апертурΚ (от лат. apertura - отверстие). Апертуры различают по форме, местоположению, по их числу и строению. На первых этапах формирования спор все типы апертур представляют сквозные отверстия, обеспечивающие контакт цитоплазмы споры с полостью спорангия. В это время они являются основным местом поступления питательных веществ в развивающуюся спору. У бессеменных растений апертуры в течение всей жизни остаются сквозными отверстиями, через которые содержимое споры при прорастании выходит наружу. Для семенных растений характерно, что на определенной стадии их развития места апертур покрываются оболочкой, отличающейся по строению от остальной части спородермы. Обычно здесь структуры, содержащие спорополленин, не имеют между собой прочной связи и легко раздвигаются при набухании цитоплазмы и прорастании споры.
Тип апертур характерен для определенных систематических групп. Так, у бессеменных растений апертуры представляют однолучевые или трехлучевые щели, расположенные на проксимальной поверхности и обычно раскрывающиеся по тетрадному рубцу - месту сочленения спор в тетраде. У семенных растений на самых ранних стадиях формирования спородермы на проксимальной стороне микроспоры появляется совершенно новый слой спородермы, отделяющий споры друг от друга, в результате чего тетрадный рубец и проксимальные щели не развиваются. Строение проксимальной и дистальной сторон спородермы оказалось резко различным. Функции регулирования объема и прорастания теперь стала выполнять ее дистальная часть. Так в эволюции растений возникли апертуры совершенно нового типа - дистально расположенные борозды, представляющие специальный проростковый аппарат, имеющий, в отличие от сквозных апертур, сложное строение.
За время существования семенных растений в строении и расположении апертур произошли существенные изменения. Дистальные одиночные борозды превратились в поры (апертуры, имеющие округлую форму). Появился тип спородермы с экваториально расположенными апертурами: несколькими бороздами, порами, сложными апертурами, совмещающими в себе борозды и поры, и т. д. Во многих линиях развития происходит процесс разделения одной апертуры на много мелких, более или менее равномерно расположенных по спородерме.
У наиболее примитивных современных голосеменных и цветковых растений имеется широкая дистальная борозда. В различных эволюционных линиях голосеменных растений наблюдается постепенная редукция дистальной борозды и образование лептомы - дистальной борозды, не имеющей четкого бороздного края и нередко трудно отличимой от остальной поверхности пыльцевого зерна. Пыльцевые зерна специализированных голосеменных снабжены одной поровидной дистальной апертурой или полностью лишены апертур. В последнем случае вся поверхность пыльцевого зерна имеет строение, типичное для апертурной зоны. У цветковых растений выработались более разнообразные апертуры, расположенные, помимо дистальной поверхности, по экватору или равномерно по всей поверхности пыльцевого зерна. Но об этом мы подробнее расскажем в следующем томе "Жизни растений".
Существенным признаком в строении спородермы является также ее форма, определяющаяся многими факторами: соединением спор в тетраде, расположением апертур, свойствами каллозы и т. д. Для бессеменных растений наиболее характерны споры, имеющие тетраэдрическую (плауны, многие папоротники, мхи) или бобовидную, переходящую в эллипсовидную (многие папоротники) форму. Хвощи и большинство моховидных имеют шарообразные споры. Голосеменные унаследовали эллипсовидную и шарообразную формы, в некоторых линиях эволюции трансформировав ее путем образования воздушных мешков (кордаитовые, многие хвойные). Воздушные мешки представляют полые выросты наружного слоя спородермы, они обычно расположены по экватору пыльцевого зерна. Они, как и многие другие структуры спородермы, по-видимому, неоднократно и независимо возникали в различных эволюционных линиях высших растений. Так, хорошо развитые воздушные мешки обнаружены на многих ископаемых спорах, принадлежащих плауновидным и семенным папоротникам.
Воздушные мешки используются в качестве пассивного летательного аппарата, кроме того, представляя собой свободные полости, они позволяют содержимому споры легко менять объем. Некоторые авторы считают, что воздушные мешки помогают пыльцевому зерну правильно ориентироваться на поверхности прорастания.
Спородерма современных растений составлена из нескольких хорошо различимых слоев, имеющих различное происхождение и строение. Самый наружный слой - перина (или периспорий) - характерен для многих спор мхов, папоротников, хвощей. В виде фрагментов он образуется у некоторых голосеменных и, как исключение, представляет самый верхний слой спородермы некоторых очень примитивных цветковых растений. Он формируется из структур, образующихся в клетках стенки спорангия или гнезда пыльника. Следующие слои спородермы - экзина (экзоспорий) и интина (эндоспорий), образуются протопластом самой споры. От клеток стенки спорангия они получают лишь строительный материал. Экзина, так же как и перина, состоит главным образом из спорополленина - высокомолекулярного вещества, обладающего исключительной стойкостью (одного из наиболее стойких в органическом мире). Он вырабатывается в клетках стенки спорангия путем окислительной полимеризации каротиноидов и каротиноидных эфиров. В экзине выделяются два основных слоя: эктэкзина и эндэкзина, а нередко также третий, промежуточный между ними слой - мезэкзина. На формирование эндэкзины и, по-видимому, мезэкзины, в отличие от эктэкзины, расходуется спорополленин, вырабатываемый в цитоплазме самой споры. Поэтому эктэкзину и эндэкзину, помимо морфологических признаков, различают по химическому составу, электронной плотности, флуоресценции и способности к окрашиванию.
Интина эластична, бесцветна, не содержит спорополленин и легко разрушается химическими реагентами. Она состоит из целлюлозной основы, пропитанной пектиновыми веществами, имеет белки и ферменты. При прорастании спор интина растягивается.
Морфологические признаки слоев спородермы формируются постепенно в процессе развития спор. Сразу же после мейоза вокруг каждой споры образуется временная оболочка - каллоза (от лат. callus - толстая кожа, мозоль), почти целиком состоящая из полисахаридов. Каллоза имеет особенно большое значение на тех ранних этапах жизни споры, когда последние еще не защищены собственной оболочкой. При непосредственном участии каллозы определяется форма споры. Первоначально каллоза, по-видимому, была "мягкой" оболочкой и недостаточно препятствовала давлению клеток друг на друга, в результате споры многих вымерших и современных растений имеют тетраэдрическую форму. Эволюционно более подвинутые растения выработали прочную каллозу, которая даже при значительном механическом воздействии сохраняет первоначальную конфигурацию. Под такой каллозой споры не испытывают давления со стороны друг друга и после полимеризации спорополленина может быть зафиксирована любая форма спородермы.
Под каллозой закладывается первая собственная оболочка споры - эктэкзина. В ней уже на самых ранних стадиях развития намечаются места апертур и внутренняя структура. У бессеменных растений в образовании эктэкзины принимают участие плазмалемма (наружный более плотный слой цитоплазмы) и прилегающие к ней мембраны цитоплазмы. У семенных растений между каллозой и плазмалеммой формируется специальный слой - матрикс эктэкзины. На матриксе (от лат. matrix - матка) полимеризуются предшественники спорополленина. В настоящее время установлено, что структура эктэкзины целиком зависит от структуры ее матрикса. Во время полимеризации спорополленина происходит лишь утолщение слоя без значительного изменения его строения. В различных линиях высших растений формирование эктэкзины происходит по-разному.
Интина, являющаяся непосредственным продолжением цитоплазмы, также представляет собой сложное образование и нередко состоит из нескольких слоев, отличающихся по химическому составу и электронной плотности.
Уже давно было замечено, что морфологические особенности спородермы, как количественные, так и структурные, довольно постоянны для каждой систематической группы. Не только роды, но даже многие виды настолько хорошо отличаются деталями строения спородермы, что, изучая под микроскопом оболочку споры, можно определить тот род, а часто и вид, к которому она относится. Это обстоятельство уже сравнительно давно используется для изучения истории флоры и растительности, для решения вопросов геологической стратиграфии, археологии, этноботаники. Изучение морфологических особенностей спородермы используется также в медицине (выяснение причин возникновения сенной лихорадки и других аллергий), криминалистике, в установлении ботанического состава пыльцы в перге и меде и пр. Исключительно большое значение имеет сравнительная морфология спородермы для систематики растений. Поэтому изучение спородермы выделилось в самостоятельную ботаническую дисциплину, которую с 1944 г. называют палинологией (от греч. paleno - сыплю, посыпаю и logos - учение). Следует отметить, что предметом палинологии является исключительно спородерма, а не содержимое спор. Поэтому для палинолога не столь существенно, в какой стадии развития находится это содержимое - в стадии одной клетки или в стадии развивающегося гаметофита. Живое содержимое споры и ее дальнейшее развитие являются предметом изучения другой ботанической дисциплины - эмбриологии.
Гаметофит, гаметангии и оплодотворение (А. Л. Тахтаджян)
Содержимое споры представляет собой первую клетку гаметофазы, и из нее в результате ряда митотических делений развивается гаметофит. Каждая изоспора дает начало обоеполому многоклеточному гаметофиту, на котором развиваются многоклеточные половые органы (гаметангии) - антеридии и архегонии. Из гетероспор также развиваются гаметофиты, но содержимое микроспор дает начало только мужским гаметофитам, а содержимое мегаспор - женским гаметофитам. В отличие от обоеполых гаметофитов однополые гаметофиты, особенно мужские, состоят из гораздо меньшего числа клеток и характеризуются возрастающей в процессе эволюции высших растений редукцией гаметангиев.
Происхождение многоклеточных гаметангиев высших растений, защищенных слоем стерильных клеток, было связано с условиями наземного существования. В то время как в воде нет необходимости в защите гамет от высыхания, на суше эта задача является одной из важнейших. Не удивительно поэтому, что даже у наиболее примитивных из известных нам высших растений их нежные половые клетки развиваются в особых вместилищах, защищающих их от высыхания. Каково же происхождение этих вместилищ?
Многоклеточные антеридии и архегонии высших растений произошли скорее всего из многоклеточных же половых органов, которые имеются у некоторых водорослей, в частности у зеленых. Но в многоклеточных гаметангиях водорослей все клетки половых органов фертильны и лишены защитной стенки.
Очевидно, такие гаметангии не могли удовлетворить потребностям наземного существования. С переходом водорослевого предка высших растений к наземному образу жизни возникла прежде всего потребность в защите гамет от высыхания. Как в мужских, так и в женских гаметангиях самые наружные клетки должны были в этих условиях стерилизоваться, т. е. потерять способность производить гаметы, и после соответствующих видоизменений превратиться в защитный слой. Предполагают, что именно таким путем, т. е. путем стерилизации наружного слоя клеток, развилась стенка антеридиев и архегониев.
У водорослевого предка высших растений мужские и женские гаметы отличались друг от друга, вероятно, очень слабо и были все снабжены жгутиками. Но у высших растений половая дифференциация усилилась и привела к резкому диморфизму гамет. Женские гаметы (яйцеклетки) накапливают питательные вещества и поэтому более крупны и неподвижны. Мужские гаметы (сперматозоиды) почти совершенно лишены запасных питательных веществ. У низших групп высших растений они сохраняют свою подвижность. Мужские гаметы потеряли свои жгутики у большинства голосеменных и у всех цветковых растений, но здесь это компенсировалось специальным приспособлением для их доставки - пыльцевой трубкой. Такое неравномерное распределение питательного материала между женскими и мужскими гаметами обеспечивает большее число копуляций (слияний) при одной и той же массе, чем если бы эта масса питательных веществ была распределена равномерно между гаметами.
Таким образом, диморфизм гамет имеет большое биологическое значение и усиливался в процессе естественного отбора.
С усилением половой дифференциации количество мужских гамет в гаметангии возрастало. Так получился антеридий высших растений с его многочисленными мелкими сперматозоидами. Количество женских гамет, напротив, уменьшилось. Таким путем возник в конце концов архегоний с одной большой и неподвижной яйцеклеткой. В эволюции архегония стерилизация сказалась весьма сильно, а сама женская гамета лишилась жгутиков и стала неподвижной. Шейковая и брюшная канальцевые клетки архегония также были когда-то гаметами, но в процессе эволюции стерилизовались. У разноспоровых растений в связи с редукцией их гаметофитов редуцируются и видоизменяются также антеридии и архегонии, а у голосеменных родов вельвичия и гнетум и у всех цветковых растений они уже вовсе не образуются.
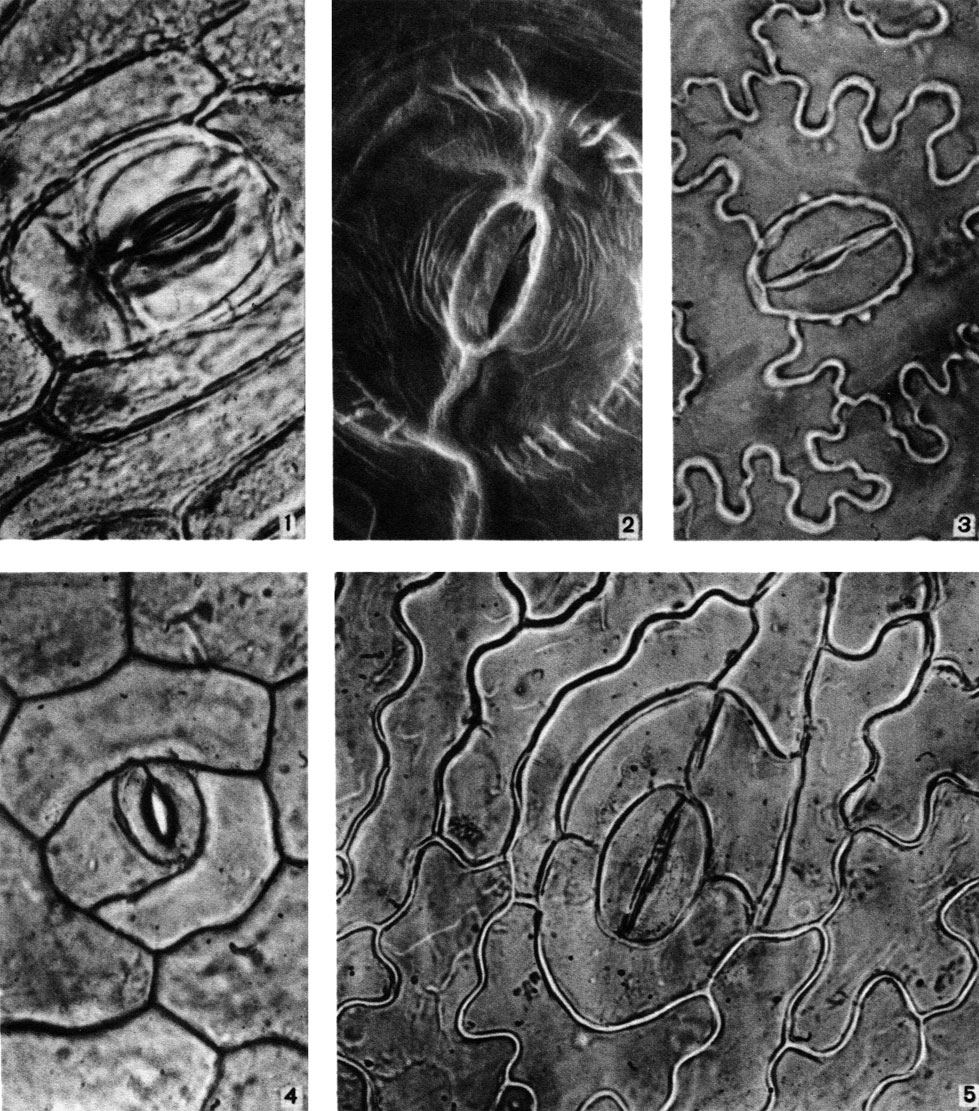
Таблица 2. Типы устьичного аппарата: 1 - тетрацитный у хангуаны малайской (Hanguana malayana) из цветковых (увел, около 900); 2 - аномоцитный у полиподиума обыкновенного (Polypodium vulgare) из папоротниковидных (под сканирующим электронным микроскопом, увел, около 2600); 3 - полоцитный у вудвардии укореняющейся (Woodwardia radicans) из папоротниковидных (увел, около 600); 4 - анизоцитный у пирикветы глянцевитой (Piriqueta nitida) из цветковых (увел, около 900); 5 - ставроцитный у нефролеписа сердцевиднолистного (Nephrolepis cordifolia) из папоротниковидных (увел, около 600). У последнего вида имеются также устьица диацитного типа (см. табл. 1, 4)

Разделение подцарства высших растений на отделы
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'