Эволюция проводящей системы (А. Л. Тахтаджян)
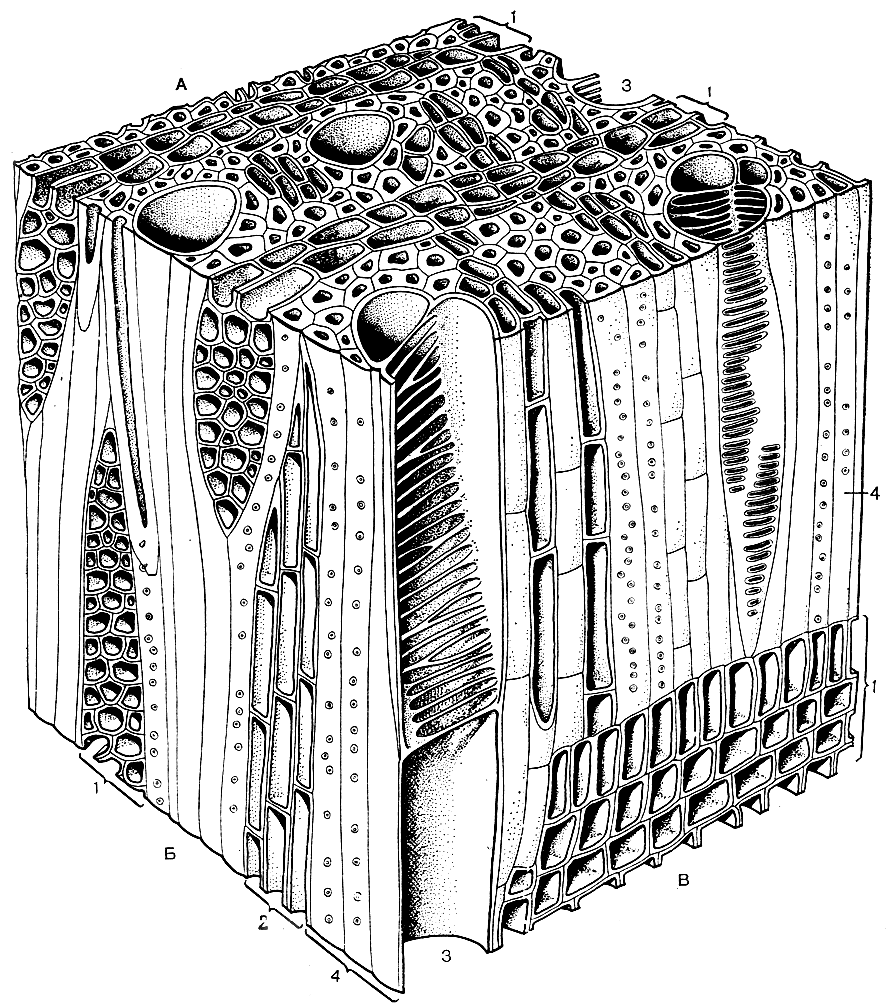
Рис. 1. Трехмерная блок-диаграмма древесины дегенерии фиджийской (Degeneria vitiensis). А - поперечный разрез. Б - тангентальный разрез. В - радиальный разрез: 1 - древесинные лучи; 2 - древесинная паренхима; 3 - сосуд; 4 - волокна
Из вводной главы предыдущего тома мы уже знаем, что проводящая система цветковых растений достигла наиболее высокого уровня эволюционного развития. Проводящая система у цветковых растений оказалась значительно более совершенной, чем у голосеменных (рис. 1), а тем более у папоротников и других групп высших растений. Впрочем, некоторые из наиболее примитивных представителей ныне живущих двудольных, такие, как виды семейства винтеровых (Winteraceae) и роды троходендрон (Trochodendron) и тетрацентрон (Tetracentron), по строению проводящей системы мало чем отличаются от примитивных представителей голосеменных типа современных саговниковых или вымерших беннеттитовых. У названных родов нет сосудов, а имеются только трахеиды. В протоксилеме эти трахеиды с кольчатыми и спиральными утолщениями, в метаксилеме обычно лестничные. Бессосудистые формы имеются также среди немногих травянистых двудольных и у ряда однодольных. Конечно, иногда отсутствие сосудов у травянистых растений - явление вторичное (у рясковых).
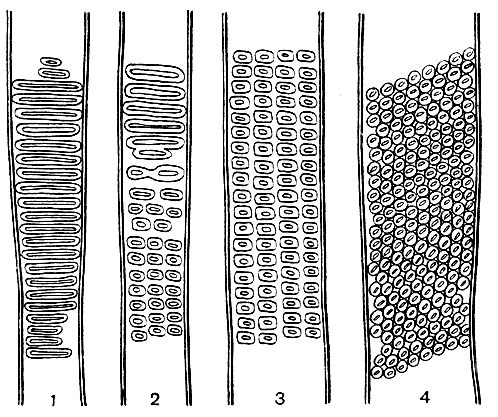
Рис. 2. Различные эволюционные типы боковой поровости сосудов: 1 - лестничная у иллициума мелкоцветкового (Illicium parviflorum); 2 - промежуточная у магнолии вирджинской (Magnolia virginiana); 3 - супротивная у тюльпанного дерева (Liriodendron tulipifera); 4 - очередная у миндаля Фенцля (Amygdalus fenzliana), увел. 400
Лишь у относительно немногих цветковых растений сохранилась бессосудистая ксилема. У подавляющего их большинства наряду с трахеидами имеются также сосуды, являющиеся основными водопроводящими элементами. В отличие от трахеиды каждый членик сосуда (каждый его клеточный элемент) имеет сквозные отверстия, называемые перфорациями (от лат. perforare - пробуравливать). Наиболее примитивные членики сосудов очень похожи веретеновидной формой и заостренными концами на трахеиды. Они очень длинные, узкие, в поперечном сечении угловатые, тонкостенные, не имеют конечной стенки или с очень слабо выраженной и очень косой конечной стенкой (рис. 3). Боковые стенки таких сосудов (рис. 2) имеют еще лестничные окаймленные поры (лестничную поровость), а перфорационная пластинка, т. е. место соприкосновения и сообщения двух соседних члеников, состоит из многочисленных (иногда до 100 - 150) лестничных перекладин, чередующихся с продолговатыми перфорациями. Один из крупнейших анатомов прошлого столетия, немецкий ботаник Антон де Бари (1877) впервые высказал мысль, что лестничная перфорация члеников сосудов возникла из лестничной поровости трахеид в результате исчезновения замыкающих пленок пор в местах соприкосновения налегающих друг на друга стенок соседних трахеид. В эволюционном смысле от лестничной трахеиды до членика сосуда с лестничной перфорацией лишь один шаг, и неудивительно поэтому, что сосуды возникли независимо и гетерохронно в разных линиях развития цветковых растений. Они возникли не только совершенно независимо у двудольных и однодольных, но появились независимо даже в разных группах как двудольных, так и однодольных. Превращение лестничных трахеид в членики сосудов - один из ярких примеров параллельной эволюции.
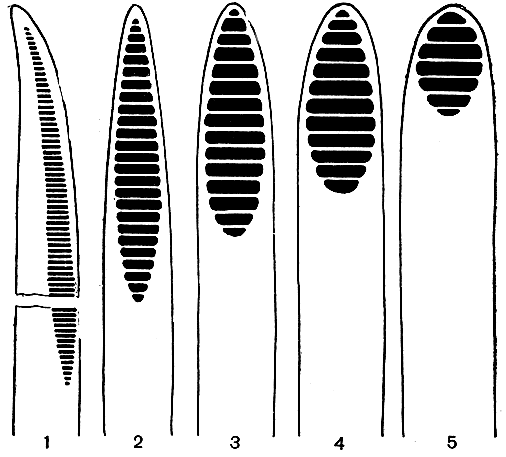
Рис. 3. Стадии эволюции лестничной перфорации (1 - 5) от примитивной перфорации с многочисленными перекладинами (1) до специализированной, имеющей только несколько перекладин (5)
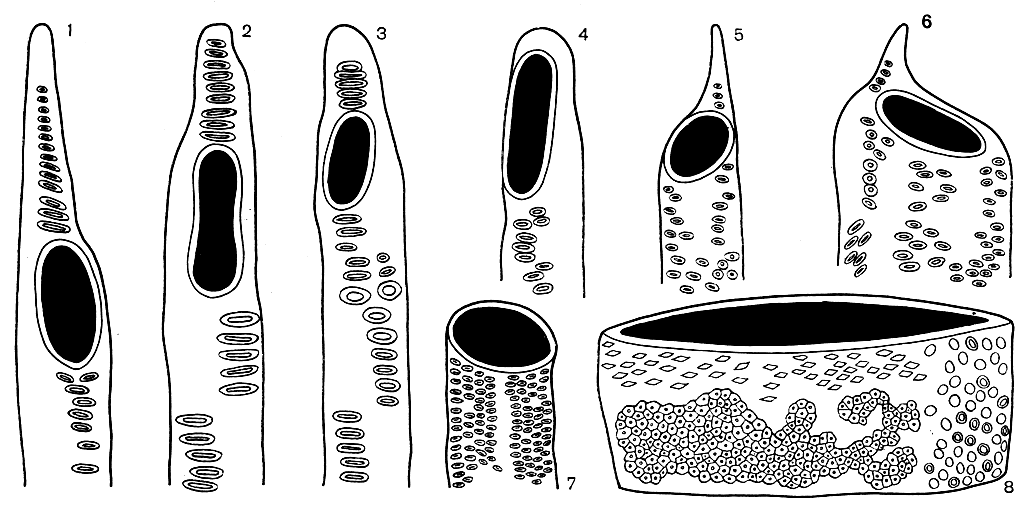
Рис. 4. Стадии эволюции члеников сосудов с простой перфорацией: 1 - магнолия крупнолистная (Magnolia macrophylla); 2 - магнолия лилиецветковая (М. liliflora); 3 - магнолия кобус (М. kobus); 4 - магнолия голая (М. denudata); 5 - яблоня Зибольда (Mains sieboldii); 6 - камфорный лавр (Ciimamomum camphora); 7 - бакхарис лебедолистный (Baccharis halimifolia); 8 - шелковица белая, или тут (Morus alba), увел. 250
Как и все другие структурные элементы, членики сосудов в процессе эволюции совершенствуются. Длина их постепенно уменьшается, они становятся шире и в большинстве случаев приобретают более толстые стенки. Сечение их на поперечном срезе становится округлым, лестничные боковые поры заменяются более или менее округлыми окаймленными порами, которые располагаются сначала в горизонтальных рядах (супротивная поровость), а затем расположение их становится очередным, в виде косых рядов (очередная поровость, рис. 2). Возникают ясно выраженные конечные стенки, на первых этапах эволюции еще очень косые. Постепенно они принимают поперечное положение, т. е. располагаются под прямым углом к длине сосуда. По мере укорочения длины и увеличения диаметра члеников сосуда отверстия в лестничной перфорационной пластинке расширяются, число перекладин уменьшается, и в конце концов после исчезновения всех перекладин образуется одна большая перфорация, называемая простой (рис. 4). Простая перфорация - это наиболее совершенный тип сквозного отверстия между члениками сосудов, так как сопротивление току жидкости сведено здесь к минимуму. Высшим, наиболее совершенным типом членика сосуда является короткий бочонкообразный членик, ширина которого превосходит его длину (рис. 4). Эволюция члеников сосудов - один из самых ярких и наиболее документированных примеров приспособительной эволюции. Это также пример эволюционного ряда, который завершается кульминационным типом, представляющим собой конечное звено в цепи структурных преобразований.
Эволюция ситовидных трубок цветковых растений также начинается с очень примитивных типов, близких к ситовидным клеткам голосеменных. Членики ситовидных трубок отличаются от ситовидных клеток голосеменных главным образом наличием ясно выраженных ситовидных участков, представляющих собой более тонкие места (углубления) первичной стенки, пронизанные порами, через которые протопласты соседних члеников сообщаются по средством связующих тяжей. Предполагают, что поры обычно возникают из каналов плазмодесм путем их ферментативного "рассверливания". Эти ситовидные участки представляют собой видоизмененные первичные поровые поля обыкновенных паренхимных клеток. Связующие тяжи ситовидных участков значительно толще плазмодесм первичных поровых полей, и, кроме того, каждая пора в ситовидном участке обычно содержит маленький каллозовый цилиндр, через который проходит тяж (каллоза - полисахарид, состоящий из остатков глюкозы, соединенных в спиральную цепочку). У голосеменных связующие тяжи еще тонкие и похожи на обыкновенные плазмодесмы, но у цветковых растений они достигают часто значительной толщины. В процессе эволюции происходит постепенное утолщение связующих тяжей и окружающих их каллозовых трубок.
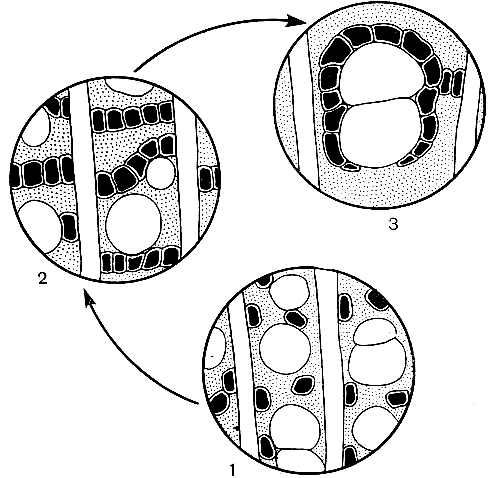
Рис. 5. Схема эволюции основных типов древесинной паренхимы: 1 - диффузная; 2 - метатрахеальная; 3 - вазицентрическая, или околососудистая
На более ранних стадиях эволюции ситовидных трубок все ситовидные участки данного членика одинаковы, но затем начинают выделяться участки с более развитыми каллозовыми трубками. Такие, более специализированные ситовидные участки обычно локализуются на определенных стенках члеников, чаще всего на конечных. Части стенки, несущие такие, более специализированные ситовидные участки, называются ситовидными пластинками. Ситовидная пластинка может состоять из нескольких или многих ситовидных участков (с лестничным, сетчатым или иным их расположением). Такая пластинка носит название сложной. Если ситовидная пластинка состоит из одного ситовидного участка, ее называют простой.
Наиболее примитивные формы ситовидных трубок состоят из довольно длинных узких и заостренных члеников с очень косыми конечными стенками и с более или менее одинаковыми ситовидными участками на конечных и боковых стенках. Подобно конечным стенкам члеников сосудов, конечные стенки члеников ситовидных трубок в процессе эволюции постепенно принимают все менее наклонное положение и в конце концов часто становятся поперечными, т. е. располагаются под прямым углом к боковым стенкам. Одновременно с этим происходит постепенная локализация ситовидных участков на конечных стенках. При этом сложные ситовидные пластинки переходят в простые, более приспособленные для транспорта ассимилятов в растении. Этот процесс аналогичен превращению лестничной перфорации члеников сосудов в простую. В обоих случаях совершенствуется механизм передвижения жидкостей.
Наконец, в процессе эволюции цветковых растений происходило уменьшение длины и увеличение диаметра члеников ситовидных трубок, что, однако, не привело здесь к тем очень коротким и широким бочонкообразным отдельностям, которые встречаются у высокоспециализированных сосудов.
Наблюдается определенная корреляция в эволюции ситовидных трубок и сосудов, и, как правило, уровень специализиции ситовидных трубок соответствует уровню развития сосудов.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'