Трутовые грибы (М. А. Бондарцева)
Трутовые грибы в подавляющем большинстве - разрушители древесины, причем во многих случаях именно они оказываются первопричиной поражения и последующей гибели живых деревьев. Вегетативное тело гриба (мицелий) развивается и функционирует в древесине живых или мертвых стволов, корней, пней, а реже - в ветвях деревьев и кустарников.
Гифы у трутовых грибов, как правило, тонкие (диаметром 2-5 мкм), бесцветные, изредка буроватые, иногда с более или менее многочисленными пряжками и всегда с поперечными перегородками (септированный мицелий). Вегетативный мицелий способствует распространению грибов в субстрате и их питанию.
Живой мицелий грибов содержит обычно более 90% воды, которую теряет при высушивании. Плодовые тела, особенно с ди- и тримитической гифальной системой, несколько менее водянисты, но процент воды в них все же высок.
Минеральное питание трутовиков как дереворазрушающих грибов определяется характером субстрата. Древесина растущего дерева относительно бедна золой, содержание которой колеблется в пределах 1% массы сухого вещества. Вследствие этого требования трутовиков к минеральным элементам питания очень малы. В золе грибов обычно много Р2О5 (до 40-50%), содержание К2О достигает 20-30%. Остальные элементы представлены единицами процентов или их дробями. Увеличение общей концентрации минеральных веществ в субстрате влияет на содержание золы в грибах, вызывает интенсивный рост и более мощную разрушительную деятельность.
Азот как важнейший элемент питания в органических соединениях более доступен для трутовых грибов, чем минеральный (в виде нитратов и солей аммония). С увеличением азота в древесине возрастают скорость роста грибов и интенсивность их деятельности. Например, древесина, пропитанная 1%-ным раствором пептона, разлагается рядовым трутовиком (Соriolellus serialis) на 50% и больше, чем контрольная. Рост и активность дереворазрушающих грибов в большой степени зависят от содержания азотных соединений в субстрате. Гифы проникают в древесину прежде всего по сердцевинным лучам и в них же больше всего разрастаются, поскольку сердцевинные лучи содержат основную массу азотных соединений. Поглощение азота, связанного в неорганическом ионе, зависит от реакции субстрата и от характера амфолитических цитоплазматических белков, так как в среде более кислой, чем оптимальная, белки цитоплазмы некоторых грибов проявляются как катионы, в менее кислой - как анионы.
Важной составной частью питания грибов, особенно патогенов живых деревьев, служат вещества витаминного характера и стимуляторы роста. Некоторые трутовики, например ложный осиновый, быстро отмирают после рубки ствола, несмотря на то что химический состав древесины, ее физическое состояние и прочие условия сохраняются, но прекращается синтез этих веществ.
Грибы как гетеротрофные организмы получают энергию только в процессе респирации (дыхания), выделяя углекислый газ и воду. Трутовым грибам присущ аэробный тип дыхания. Достаточное поступление кислорода воздуха и одновременный отвод продуктов дыхания являются важным условием для возникновения гнили в растущем дереве. С точки зрения физиологического состояния древесины для деятельности грибов самым подходящим является ядро, которое у растущих деревьев, как правило, заражается.
Рост грибов и выделение ими углекислоты зависят от давления кислорода в атмосфере и от температуры. При меньшем давлении кислорода чем 1,5 атм и температуре 17,5° С гриб перестает расти и его обмен приобретает анаэробный характер. Нижняя граница аэробного дыхания зависит от температуры: при 29,5° С обмен веществ уже меняется при давлении кислорода 1,5 атм. В условиях анаэробного дыхания выделение углекислоты прямо пропорционально давлению кислорода. В таких условиях изменяется весь обмен веществ гриба, весь набор его ферментов. Например, кориолус многоцветный (Coriolus versicolor) может выделять фосфатазу и мутазу и вызывать спиртовое брожение.
Целлюлозоразрушающие и лигнинразрушающие грибы потребляют целлюлозу или лигнин, благодаря чему древесина приобретает красновато-бурую или светлую окраску. Посветление древесины при разложении лигнинразрушающими грибами вызвано окислительными ферментами, которые выделяют грибы в древесину. Потемнение древесины, вызванное целлюлозоразрушающими грибами, зависит от веществ гумусового характера. Некоторые дереворазрушающие грибы выделяют также пигменты, окрашивающие древесину в иные тона. Подобным образом корневая губка в первой стадии разложения окрашивает древесину в оттенки фиолетового цвета. У дереворазрушающих грибов встречается и изменение окраски гиф. Та же корневая губка образует в трахеидах бесцветные гифы. В сердцевинных лучах гифы бурого цвета и вызывают буровато-фиолетовое окрашивание.
Считают, что выделение грибами пигмента связано с недостатком органического или неорганического азота. Это доказывается тем, что на средах с достаточным количеством азота пигмент не выделяется, а в естественных условиях выделение пигмента в массу клеточных оболочек наступает только после определенного времени (когда израсходованы все азотсодержащие питательные вещества в сердцевинных лучах). Однако корневая губка выделяет пигменты как раз при достаточном количестве азота (в сердцевинных лучах), а при переходе в трахеиды, практически не содержащие азота, выделение пигмента прекращается.
Питание и метаболизм трутовых грибов зависят от свойств субстрата или физиологического состояния хозяина.

Рис. 160. Ложный осиновый трутовик (Phellinus tremulae)
Большая часть веществ, необходимых грибам для питания, находится в нерастворимом состоянии, особенно это касается источников углеродного питания. Поэтому огромную роль в жизни грибов играют энзимы, или ферменты, переводящие различные соединения из нерастворимого состояния в растворимое.
Основное значение в разложении древесины имеют экзоферменты, выделяемые грибом в древесину. Продукты ферментативного расщепления доступны грибам и являются для них источником питания и энергии. Эндоферменты не выделяются в субстрат. Оставаясь внутри живых клеток, они обеспечивают их внутренние метаболические процессы. В субстрат они попадают как продукты разложения самой грибной клетки.
Для трутовых грибов как разрушителей древесины наиболее важным экзоферментом следует признать целлюлозу. Все без исключения дереворазрушающие грибы обладают способностью разлагать целлюлозу, ибо этот фермент есть в клетках всех трутовиков.
Наивысшая активность большинства ферментов наблюдается у целлюлозоразрушающих грибов. У лигнинразрушающих грибов наряду с гидролитическими ферментами существуют также специфические ферменты - оксидазы. Из этой группы ферментов наиболее распространены тирозиназа, встречающаяся у некоторых дереворазрушающих и напочвенных агариковых грибов, лакказа и особенно пероксидаза. Очень много пероксидазы выделяют грибы из рода феллинус (Phellinus), особенно ложный трутовик (P. igniarius). Грибная пероксидаза выделяется только вегетативным мицелием и не отмечена в плодовых телах.
Установлено, что проба на оксидазу может считаться надежным критерием при разделении грибов на целлюлозоразрушающие и лигнинразрушающие. Случаи нечеткого проявления реакции представляют исключение, причем наблюдаются не у трутовых грибов.
Деятельность экзоферментов играет основную роль не только в питании, но и в распространении трутовых грибов по субстрату. Проникновению в клетку гифы дереворазрушающего гриба предшествует растворение клеточной оболочки экзоферментами гриба. Отверстие, образующееся в оболочке древесной клетки, всегда несколько шире, чем диаметр гифы. Никогда мицелий трутовиков не распространяется в древесине, разрушая ее механическим давлением. Гифы могут проникать в клетки древесины, раство-ряя оболочку в любом месте. В зависимости от ферментативного комплекса трутовые грибы способны вызывать бурую деструктивную или белую коррозионную гниль.
Целлюлозоразрушающие грибы, не вырабатывающие оксидаз, вызывают деструктивную гниль. Древесина в начальной стадии разрушения изменяет окраску от красноватой до ржаво-красной и, наконец, становится темно-бурой от освобожденного лигнина, хрупкой, легко ломающейся и крошащейся, заметно теряет в объеме и массе. Часто в конечной стадии разрушения древесина растрескивается на крупные или мелкие призматические кусочки. В зависимости от оттенка разрушаемой древесины деструктивная гниль делится на группы красной и бурой. Из широко распространенных трутовиков деструктивную гниль вызывают заборный трутовик (Gloeophyllum sepiarium), белый домовый гриб (Coriolus vaporarius), рядовой трутовик (Coriolellus serialis), пахучий трутовик (Osmoporus odoratus) и др.
Лигнинразрушающие грибы, вырабатывающие оксидазы, разлагают и целлюлозу, и лигнин. Иногда в начальной стадии разложения древесина может временно потемнеть, вообще же она светлоокрашенная, отчего и гниль носит название белой. Влияние различных видов лигнинразрушающих грибов на древесину неодинаково, отчего различается и внешний вид поражения. Иногда древесина белеет равномерно по всей пораженной части, иногда появляются только светлые полосы или ячейки, заполненные неразложившейся целлюлозой (ямчатая гниль). В конечной стадии гниения древесина становится мягкой, волокнистой, часто расслаивается параллельно годичным кольцам, иногда крошится, теряет в массе, но ее объем не уменьшается, поэтому призматического растрескивания не происходит. Ямчатую гниль часто называют также пестрой или куропатчатой.
Рис. 161. Серно-желтый трутовик (Laetiporus sulphureus)
Белую коррозионную гниль вызывает широко распространенный в наших лесах плоский трутовик (Ganoderma applanatum, рис. 163), а также многие грибы родов инонотус (Inonotus) и феллинус (Phellinus). Пестрая гниль характерна при поражении древесины сосновой губкой (Phellinus pini, рис. 162), или корневой губкой (Fomitopsis annosa).

Рис. 162. Сосновая губка (Phellinus pini)

Рис. 163. Плоский трутовик (Ganoderma applanatum)
Некоторые трутовики вызывают поражение древесины, не укладывающееся в классическое понимание коррозионной или деструктивной гнили. Этот тип поражения носит специальное название прелости (задыхания) древесины лиственных пород и вызывается так называемыми штабельными грибами. Задыхание начинается весной или летом, т. е. в теплое время года, с характерного побурения древесины. Побурение начинается с торцов бревна, часто с двух сторон, и в виде конуса продвигается к центру. Затем, как правило, оба конуса сливаются. Прелость - сложный биологический процесс постепенного отмирания древесины (понятие прелости впервые четко разработано А. Т. Вакиным). Первая его стадия - побурение древесины - еще не связана с заражением грибами. Вторая стадия - подпар - уже связана с заражением грибами, древесина имеет не ровную окраску, а полосами, более светлыми, но не белыми. Вторая грибная фаза соответствует третьей фазе разрушения и называется "мраморная гниль" или просто "мрамор". Конечная фаза прелости - мягкая гниль - хорошо заметное на глаз и на ощупь разрушение древесины. Из трутовиков такой тип гнили вызывает настоящий трутовик (Fomes fomentarius, табл. 29) и грибы рода Coriolus.
Взаимоотношения грибов в процессе разложения древесины определяются тем, что гриб, способный разлагать здоровую древесину, подготавливает субстрат для следующего вида. При этом надо помнить, что в процессе истощения питательных веществ гриб, поселившийся первым, становится менее жизнеспособным, тогда как тот, для которого частично разложившаяся древесина является оптимальной средой, приобретает наиболее благоприятные условия для развития и он сравнительно легко вытесняет своего предшественника. Поданным Рипачека, такой парой являются, например, окаймленный трутовик (Fomitopsis pinicola) и пахучий трутовик (Osmoporus odoratus). Первый гриб поселяется на здоровых пнях, иногда даже на живых деревьях. Пахучий трутовик разрушает древесину значительно медленнее, но, как показывают опыты, после месяца развития окаймленного трутовика активность пахучего трутовика на подготовленной древесине значительно возрастает. Однако следует учитывать, что изменение температурно-психрометрических условий изменяет метаболизм грибов, а следовательно, и возможную последовательность развития их.
Размножение трутовых грибов осуществляется преимущественно базидиоспорами. Конидиальная стадия (например, у Tyromyces sericeomollis, Abortiporus biennis) и образование хламидоспор (например, Laetiporus sulphureus, Inonotus heinrichii) наблюдаются чрезвычайно редко.
Базидиоспоры образуются на базидиях. Последние развиваются в гимениальном слое на плодовых телах.
Мицелий трутовых грибов развивается внутри субстрата, в древесине, тогда как плодовые тела всегда образуются на поверхности субстрата, что позволяет базидиоспорам распространяться токами воздуха.
Плодовые тела трутовых грибов довольно крупные, хотя в некоторых случаях они не превышают 1 см диаметром, а иногда достигают всего несколько миллиметров. По форме плодовые тела трутовых грибов можно разделить на следующие основные категории: резупинатные, или распростертые; распростерто-отогнутые, или латерально (боком) прикрепленные, дифференцированные на шляпку и ножку, (табл. 28).
Распростертые плодовые тела имеют вид пленки или пластинки, в большей или меньшей степени плотно приросшей к субстрату и повторяющей особенности его поверхности. Край такого плодового тела может быть плотно приросшим и сходящим на нет, или приподнятым в виде толстого валика, или срастание его с субстратом неплотное, в этом случае при высыхании край отгибается и заворачивается, или по краю, особенно на вертикальном субстрате, образуются наплывы, напоминающие зачаточные боковые шляпки.
Сидячие плодовые тела имеют копытовидную или консолевидную, иногда языковидную форму без ножек, прикреплены боком или широким, нередко суженным основанием, в котором нетрудно угадать прототип боковой ножки. Плодовые тела сидячего типа наименее пластичны, но все же и у них имеются переходы к группе плодовых тел со шляпкой и ножкой, например серно-желтый трутовик (Laetiporus sulphureus) или грибы рода пиптопорус (Piptoporus), или к группе распростерто-отогнутых плодовых тел, например некоторые грибы рода фомитопсис (Fomitopsis), особенно окаймленный трутовик (F. pinicola). Типичные сидячие плодовые тела имеют грибы рода спонгипеллис (Spongipellis), щетинистоволосый трутовик (Inonotus hispidus), настоящий трутовик (Fomes fomentarius) (табл. 29).
Рис. 164. Кориолус многоцветный (Coriolus versicolor)
У некоторых трутовиков суженное основание прикрепленных боком плодовых тел вытягивается в более или менее длинную боковую ножку, длина которой часто зависит от положения субстрата. Трутовик Швейница (Phaeolus schweinitzii), например, у основания стволов и на корнях деревьев образует формы с зачаточной и даже заметно развитой ножкой, тогда как на вертикальном субстрате способен образовывать сидячие плодовые тела. То же относится к некоторым видам рода ганодерма (Ganoderma), особенно к G. resinaceum. В то же время другой вид этого рода - лакированный трутовик (G. lucidum, рис. 165) - всегда образует заметную боковую ножку, хотя и варьирующей длины. Центральную или эксцентрическую, реже боковую ножку образуют также представители некоторых родов трутовых грибов в широком смысле, обитающие на древесине (роды полипорус, бондарцевия) или на почве (роды альбатреллус, болетопсис). Грибы рода грифола (Gryfola) образуют многочисленные боковые или центральные шляпки на разветвленном пеньке.
Рис. 165. Лакированный трутовик (Ganoderma lucidum)
В пределах каждой из перечисленных основных форм плодовых тел можно наблюдать множество переходов и видоизменений, так что точное описание формы плодового тела трутовых грибов, по существу, невозможно. Например, резупинатные плодовые тела в зачаточном состоянии имеют, как правило, более или менее округлую форму, но затем разрастаются, следуя преимущественно поверхности субстрата, так что могут быть сильно вытянутыми, почти линейными, более или менее овальными, с неправильными очертаниями.
Плодовые тела образуют непрерывную гамму переходов из одной формы в другую. В пределах одного семейства и даже рода могут наблюдаться разнообразные по форме плодовые тела.
Размеры плодовых тел трутовиков сильно колеблются в зависимости от погоды и положения и состояния субстрата. Но и здесь наблюдаются известные границы варьирования, особенно в отношении предела максимума. Наименьшие плодовые тела достигают 0,5-1 см в диаметре, например некоторые грибы рода тиромицес (Tyromyces) - тиромицес Лоу (Т. lowei). Наиболее крупные достигают 0,5-1 м диаметром, например плоский трутовик или настоящий трутовик, и массы до 10 кг. Распростертые плодовые тела иногда вытягиваются по субстрату (валежному стволу или бревну) на 1-1,5 м. Однако это крайние пределы, а в среднем, диаметр плодовых тел трутовиков бывает 5-25 см.
Консистенция ткани плодовых тел определяется их анатомическим строением и расположением гиф. У трутовиков наблюдаются плодовые тела пленчатой, восковидной, субжелатинозной, мясистой, кожистой, пробковой, деревянистой, губчатой или волокнистой консистенции, со всеми переходами между этими основными категориями.
В некоторых случаях плодовые тела имеют двухслойную ткань, причем верхний слой шляпки состоит обычно из рыхлой, губчатой ткани, а нижний, примыкающий к трубочкам,- из более плотной и компактной, иногда субжелатинозной ткани.
Окраска плодовых тел трутовых грибов весьма разнообразна, причем цвет ткани, заметный на поперечном разрезе плодовых тел, часто отличается от окраски верхней и нижней поверхности.
Ткань в большинстве случаев белая или светлая - древесинно-желтая, кремовая, розоватая, но имеется большая группа трутовиков со щетинками в гимениальном слое, ткань которых окрашена в оттенки бурого цвета. Сравнительно немногочисленные трутовики характеризуются яркой, киноварно-красной, оранжевой, желтой, глинистой окраской ткани, причем они, как правило, легко группируются в отдельные роды.
Поверхность латерально прикрепленных или имеющих ножку плодовых тел, т. е. поверхность шляпки, может быть покрыта плотной, иногда лакированной или матовой коркой или тонкой, пергаментообразной кожицей, либо часто совсем лишена специального покрова (кутикулы). Однако такая "голая" поверхность не однотипна, она может быть морщинистой, концентрически-бороздчатой, что связано с неравномерным ростом плодового тела, бархатистой, войлочной, волосистой или грубощетинистой, причем покровы шляпки могут изменяться с возрастом: опушение или щетинистость пропадает и т. п. Окраска поверхности различается от белой и светлой (сероватой, бледно-кремовой, древесинно-желтой) до бурой, черной или различных ярких оттенков, причем цвет ткани и поверхности очень редко совпадает. Часто поверхность окрашена зонально, концентрично чередующимися более темными и более светлыми зонами, причем можно наблюдать различные оттенки бурого, коричневого, серого, желтого, оранжевого цветов. При высыхании яркая окраска часто выцветает, бледнеет или плодовое тело становится грязноватым, покрывается неопределенными бурыми или желтыми пятнами или полностью темнеет.
Такие же изменения наблюдаются в окраске гименофора. Гименофор чаще бывает белым или светлоокрашенным, но нежные оттенки розового, желтоватого, лиловатого цветов при высушивании часто изменяются на грязно- серые или темные.
По форме гименофор трутовиков чаще всего трубчатый, т. е. состоит из более или менее длинных, слоистых или однослойных трубочек, плотно сросшихся боками. В некоторых случаях трубочки расщепляются, образуя лопатообразные выросты или шипы, как у церрены одноцветной (Cerrena unicolor) или у ирпекса млечного (Irpex lacteus), или недоразвитие боковых стенок трубочек привело к образованию лаби- ринтовидного (дедалеевидного, по названию характерного гименофора рода дедалея) или пластинчатого (лензитесовидного - по названию рода лензитес) гименофора. Интересно, что трубчатый гименофор может образовывать ежегодно новые слои у многолетних видов, тогда как пластинчатый или дедалеевидный гименофор всегда однослойный, причем при развитии в течение нескольких лет пластинки удлиняются, но не образуют нового слоя.
В тропиках растут грибы из родов цикломицес (Cyclomyces) и циклопорус (Cycloporus), имеющие концентрически-пластинчатый гименофор (в отличие от радиально-пластинчатого гименофора рода лензитес).
Исключение из общего правила составляют плодовые тела печеночницы обыкновенной. Буро-красные, пропитанные красноватым соком, мясистые, языковидные шляпки этого гриба довольно часто встречаются на дубах в лесостепной зоне и на Кавказе. Трубочки в плодовых телах этого гриба - не сросшиеся боками, как у всех остальных трутовиков, а только сближенные, причем каждая развивается на особом основании. В этом случае мы имеем дело с лжетрубочками, представляющими собой на самом деле плодовые тела цифеллоидного типа, объединенные общим основанием - подстилкой, которая в процессе эволюции переместилась в верхнюю часть плодового тела, став так называемой субикулярной (подстилочной) шляпкой.
Анатомическое строение плодовых тел трутовых грибов довольно просто, так как они целиком состоят из гиф и не имеют других типов дифференциации клеток. Однако и при таком простом строении наблюдаются различные способы дифференциации в строении и расположении гиф, что позволяет судить о направлении эволюционного процесса.
В плодовых телах трутовиков обнаружены три основных типа гиф, названных генеративными, скелетными и связывающими.
Морфологические признаки в пределах каждого типа гиф довольно однообразны.
Половое размножение трутовых грибов осуществляется базидиоспорами, развивающимися на базидиях. Для трутовых грибов наиболее характерны булавовидные базидии, реже встречаются мерулиусовидные (у видов с бесцветными гифами и маленькими аллантоидными спорами, род глеопорус - Gloeoporus) и сотовидные (у некоторых видов со щетинками в гимениальном слое). Базидии и базидиоспоры по сравнению с другими элементами в строении плодовых тел наименее изменчивы, им придается большое значение в систематике.
Кроме базидий, в гимении иногда развиваются некоторые стерильные элементы: щетинки, цистиды, цистидиолы.
Щетинки - это особые толстостенные выросты гиф, развивающиеся в субгимении и выступающие между базидиями. Встречаются они только у представителей семейства гименохетовые (Hymenochaetaceae). В гимении грибов рода гименохете (Hymenochaetae), не имеющем трубочек, щетинки выполняют защитную функцию, тогда как у трутовиков с трубчатым гименофором, очевидно, щетинки становятся рудиментом.
У представителей других семейств трутовых грибов время от времени наблюдаются цистиды или цистидиолы. Цистиды - это тонко- или чаще толстостенные булавовидные или бутыльчатовидные окончания скелетных или генеративных гиф, выступающие в гимении между базидиями. Функция цистид до настоящего времени неясна, скорее всего она носит экскреторный характер, особенно если речь идет о цистидах, берущих начало из генеративных гиф. Цистиды, как и щетинки, развивающиеся из скелетных гиф, несомненно, имеют чисто механические, защитные функции или играют роль распорок между базидиями.
Цистидиолы - тонкостенные аналоги цистид или базидий, выполняющие определенные физиологические функции, точное назначение которых еще не выяснено. Кроме того, в гимении наблюдаются иногда базидиолы - недоразвитые стерильные базидии, рассматриваемые иногда так же, как аналоги цистидиол. Различрш в форме, размерах, степени инкрустированности цистид (сплошная или головчатая инкрустация) достаточно постоянны у отдельных видов, поэтому могут быть использованы как систематические признаки.
Оболочки спор (род Anomoporia), цистид (род Amylocystis), гиф (род Amyloporia) иногда содержат растворимый крахмал и в этом случае проявляют амилоидную реакцию с реактивом Мельцера (под действием иода появляется синее окрашивание). Такая реакция более характерна для некоторых групп, стоящих на более низких ступенях развития, а у трутовиков, только для немногих, сравнительно примитивных представителей.
Прорастая, базидиоспоры образуют первичный мицелий, состоящий из гаплоидных одноядерных гиф. После некоторого, обычно краткого, периода роста и развития два первичных мицелия сливаются с образованием вторичного двухъядерного мицелия, который развивается в древесине, вызывая ее разрушение, и на котором позднее образуются плодовые тела. Среди трутовиков имеются биполярные и тетраполярные виды.
Распространение трутовых грибов в природных условиях определяется рядом факторов, важнейшим из которых служит субстрат, т. е. соответствующая древесная порода в определенном состоянии. Лишь немногие трутовые грибы развиваются на почве, подавляющее большинство приурочено к обитанию на древесине. Как правило, трутовые грибы проявляют избирательность к хвойному или лиственному субстрату; узкая (как и слишком широкая) специализация у них бывает редко. Тенденцию к специализации проявляют представители родов пиптопорус (Piptoporus), инонотус (Inonotus), феллинус (Phellinus). В пределах этих родов специализированы преимущественно виды, патогенные для живых деревьев, и значительно реже сапрофиты (пиптопорус). Например, ложный осиновый трутовик (Phellinus tremulae) растет только на осине (см. рис. 160), а ложный трутовик (Pigniarius) - на многих лиственных породах, но на каждой из них развивается специализированная форма. Инонотус скошенный (Inonotus obliquus), стерильная форма которого широко известна под названием чаги, растет преимущественно на березе; трутовик I. rheades обычен на осине.
Как говорилось ранее, способность некоторых видов развиваться только на живых деревьях связана с их потребностью в витаминах, которые вырабатываются в процессе жизнедеятельности дерева. Очевидно, специализация в смысле приуроченности к отдельным породам также связана со специфическими потребностями в питании.
На живых деревьях развиваются лишь сравнительно немногие трутовики, а большая часть их поражает исключительно мертвую древесину. Следовательно, наличие соответствующей породы (или пород) - основное условие для нахождения гриба. Однако наблюдения показывают, что границы распространения древесных и кустарниковых пород шире, чем ареалы развивающихся на них дереворазрушающих грибов. Причины этого могут быть различные. Прежде всего имеет значение возраст дерева при заражении живых деревьев. Проникновению спор гриба внутрь древесины препятствует возникновение раневого ядра. Способность к образованию такого ядра более развита в молодом возрасте, ослабевая с годами. Поэтому старые деревья более подвержены заражению грибами. Условия произрастания деревьев, например горные, несколько изменяют анатомические свойства древесины, что тоже может служить препятствием к поражению трутовыми грибами, хотя непосредственное воздействие окружающих условий на гриб также имеет место.
Помимо чисто физиологических условий (например, состояния дерева, условий для проявления того или иного типа метаболизма, ферментативного комплекса), а также экологических условий, благоприятных или неблагоприятных для роста породы или развития гриба, большое значение имеет историко-географический фактор, т. е. является ли данная местность центром происхождения породы-хозяина или гриба и в каком удалении от этих центров находится.
Вследствие этого в каждом естественноисторическом районе можно установить (по К. Е. Мурашкинскому) три группы пород по отношению к заражению определенными видами дереворазрушающих грибов: 1) наиболее обычно поражаемая порода (породы); 2) породы хотя и нередко поражаемые, но для которых гриб не является повсеместно обычным засельником; 3) породы, случайно поражаемые и только в тех местностях, где они находятся в непосредственной близости к породам 1 и 2 категорий. Действительно, за исключением некоторых видов, не проявляющих избирательности к субстрату, распространение грибов по породам различается в разных географических зонах. Например, широко известный в европейской части СССР на дубе серно-желтый трутовик в Сибири встречается на лиственнице. Лаковый трутовик, который в Сибири растет также на лиственницах, в Ленинградской области встречается на ели, в меньшей степени на осине и ольхе, а на Кавказе обитает преимущественно на дубе. В то же время плоский трутовик и в европейской части СССР, и в Сибири чаще всего встречается на березе и осине.
Наличие подходящего субстрата обусловливает в первую очередь распространение трутовых грибов по типам леса и другим типам местообитаний. В остальном, кроме фактора состояния древесного субстрата (жизнеспособное дерево, угнетенное, сухостой, валеж и т. д.), наибольшее значение имеют основные экологические факторы: температура, влажность, в меньшей степени освещенность.
Температурные границы, при которых способен развиваться гриб, различны не только для разных видов, но даже для разных частей грибного организма: спор, грибницы, плодовых тел.
В естественных условиях мицелий дереворазрушающих грибов всегда многолетний, поскольку он развивается внутри древесины, т. е. в более или менее стационарных условиях. Этого нельзя сказать о плодовых телах, растущих на поверхности и подверженных в первую очередь прямому влиянию окружающей температуры. Поэтому плодовые тела, в отличие от мицелия, разделяются по продолжительности существования на три основные категории:
однолетние плодовые тела, развивающиеся в течение одного вегетационного периода, причем продолжительность их существовашш длится от 1,5-2 недель до 4 месяцев;
однолетние зимующие плодовые тела, способные перезимовывать и возобновлять споруляцию в следующем сезоне, обычно сохраняющие жизнеспособность только после мягких зим;
многолетние плодовые тела, существующие в течение многих лет, причем новый гименофор их может нарастать ежегодно (слоистый гименофор) или новый гимений может развиваться на том же гименофоре (неслоистый гименофор); как правило, плодовые тела со слоистым гименофором существуют более длительные сроки (до 10-15 лет) по сравнению с плодовыми телами, у которых гименофор неслоистый (3-4 года).
Не меньшее значение для развития трутовых грибов имеет влажность. Потребность во влажности субстрата и окружающего воздуха у разных видов неодинакова и для отдельных элементов грибного организма тоже разная. Наивысшая влажность субстрата необходима для прорастания спор.
По отношению к влажности плодовые тела могут быть выделены в основные группы - гигрофилов, мезофилов и ксерофилов.
Для развития грибницы свет не требуется, но плодовые тела при полном отсутствии света или не формируются вовсе, или представляют собой уродливые, часто стерильные образования. В нормальных условиях наиболее теневыносливыми оказываются трутовики с гигрофильными свойствами, а наиболее светолюбивыми - с ксерофильными.
Распространение трутовых грибов по типам местообитаний, таким образом, оказывается в прямой зависимости от наличия определенных пород деревьев (субстрата), а также от соответствующих условий влажности, температуры и освещенности в экологической нише. Дополнительным фактором этого является деятельность человека, которая накладывает отпечаток на состав флоры эксплуатируемых лесов. Немаловажное, хотя обычно недостаточно отмечаемое, влияние на состав флоры имеют взаимоотношения грибов, определяющие сукцессию видов.
В лесных ценозах наибольшее количество видов грибов развивается, естественно, на главной лесообразующей породе. Породы подлеска представляют собой один из главных элементов при выделении типов леса; следовательно, флора дереворазрушающих грибов на них представляет собой главный резерв различий между микофлорой разных типов. Так как из групп грибов, выделенных по отношению к влажности, группа мезофилов самая многочисленная, естественно оказывается, что флора трутовых грибов умеренно влажных типов леса наиболее разнообразна по видовому составу.
В однопородных лесах создаются более благоприятные условия для развития широкого спектра видов, чем в смешанных. Поэтому в чистых типах микофлора более богата и разнообразна, чем в смешанных лесах, где сравнительно небольшое количество видив обильно встречается на всех породах смешанного леса, тогда как специфические для каждой из этих пород виды, как правило, отсутствуют.
Наиболее широкое распространение во многих типах леса имеют грибы с различными сложными типами гифальных систем. Это подтверждает положение о том, что более прогрессивные типы анатомического строения обеспечивают больший диапазон приспособительных возможностей. Некоторое количество видов с мономитическими плодовыми телами (состоящими только из генеративных гиф), развивающимися в течение короткого периода, 2-3 недель, имеют такое же широкое распространение во многих типах леса. Однако микроклиматические условия, сопутствующие им, на редкость однотипны. Поэтому их развитие, особенно массовое, подвержено колебаниям в разные сезоны.
Флора открытых местообитаний отличается от лесной и более близка к флоре вырубок и окрестностей населенных пунктов. Последняя имеет выраженную специфику и отличается отсутствием редких "лесных" видов; обилием широко распространенных видов; преобладанием димитических и тримитических видов; обилием "штабельных" грибов и видов, развивающихся на обработанной древесине.
Специфический состав имеет флора пойм, причем некоторые виды, например душистый трутовик (Trametes suaveolens), одинаково часто встречаются в поймах рек Европы и Сибири.
Специфику каждого типа леса составляют трутовики, растущие на основной лесообразующей породе, но не переходящие из типа в тип. Виды грибов на породах подлеска, даже когда они составляют различия для смежных типов, все же нельзя рассматривать как специфические.
Установление видового состава флоры трутовых грибов как основных дереворазрушителей в пределах определенных типов леса может оказать большую помощь лесному хозяйству при выявлении опасных грибных болезней древесных пород и разработке мер борьбы с ними.
Положение трутовых грибов в системе порядка афиллофоровых нельзя рассматривать вне связи с другими группами. В СССР в настоящее время принята наиболее современная и детально разработанная система А. С. Бондарцева и Р. Зингера (1941, 1943, 1953). Порядок разделен на пять подпорядков в зависимости от характера развития плодового тела. Это позволяет выявить эволюционные линии внутри порядка.
В основе выделения семейств, состав которых в свете современных данных несколько пересмотрен, приняты такие признаки, как характер развития плодового тела и особенности микроскопического строения, имеющие значение в эволюции группы.
В указанной системе трутовые грибы распределяются по следующим семействам (описанным ниже): фистулиновые (Fistulinaceae), пориевые (Poriaceae), гименохетовые (Hymenochaetaceae), ганодермовые (Ganodermataceae), полипоровые (Polyporaceae), скутигеровые (Scutigeraceae), болетопсидные (Boletopsid асеае), бондарцевиевые (Bondarzewiaceae), кортициевые (Corticiaceae), шизофилловые (Schizophyllaceae), лахнокладиевые (Lachnocladiaceae).
Семейство Фистулиновые (fistulinасеае) (М. А. Бондарцева)
В этом семействе известен всего один род печеночница, или фистулина (Fistulina).
Плодовые тела однолетние, латерально прикрепленные, языковидные или консолевидные, у основания часто суженные, сочные, пропитанные красноватым соком, позднее волокнистые и сухие. Гименофор состоит из свободных, не срастающихся боками трубочек, свисающих с нижней стороны шляпки и представляющих собой отдельные плодовые тела цифеллоидного типа. Гифальная система мономитическая. Гифы ткани состоят преимущественно из коротких, бочонкообразновздутых клеток. Часто наблюдаются латициферы, наполненные оранжево-красным соком. Цистид нет. Споры яйцевидные, бесцветные или светлоокрашенные, гладкие, неамилоидные.
Фистулина встречается на живых стволах дубов и каштана съедобного.
Печеночница обыкновенная (Fistulina hepatica) обитает на живых старых дубах или съедобном каштане, как правило, ближе к основанию ствола. Гниль буроватая, неактивная. Встречается в зонах естественного распространения дуба, но нечасто. Плодовые тела съедобны.
Семейство Пориевые (Poriaceae) (М. А. Бондарцева)
Это собственно трутовые грибы. Плодовые тела однолетние или многолетние, распростертые, распростерто-отогнутые, сидячие, иногда с ножковидно-вытянутым основанием, толстые копытовидные или тонкие раковиновидные, различных размеров. Корка на поверхности отогнутых шляпок имеется или отсутствует. Ткань белая, светло или ярко окрашенная, мягко-пленчатой, восковидной, желатинозной, мясистой, волокнистой, деревянистой, пробковой или кожистой консистенции. Гименофор трубчатый или пластинчатый. Гифальная система моно-, ди- или тримитическая. Щетинок в гимении нет, цистиды или цистидиолы имеются или отсутствуют. Споры толсто- или тонкостенные, оттянутые или усеченные у основания, различной формы, обычно не крупные, изредка достигающие 10-12 мкм длины, бесцветные или почти бесцветные, с гладкой или почти гладкой обо-лочкой, изредка амилоидные или декстриноидные.
Пориевые грибы обитают на древесине, иногда переходя с нее на другие растительные остатки или на почву.
Род фибулопория (Fibuloporia) охватывает несколько видов.
Фибулопория моллюска (Fibuloporia mollusca) встречается на разрушенной древесине многих лиственных и некоторых хвойных (ель, сосна) пород, обычно во влажных смешанных лесах. В Советском Союзе зарегистрирован повсеместно, но встречается довольно редко.
Фибулопория кремовая (F. cremea) обитает на валежных стволах и ветках тополя и ивы, в пойменных лесах Камчатки. Ассоциируется с белой гнилью (табл. 28).
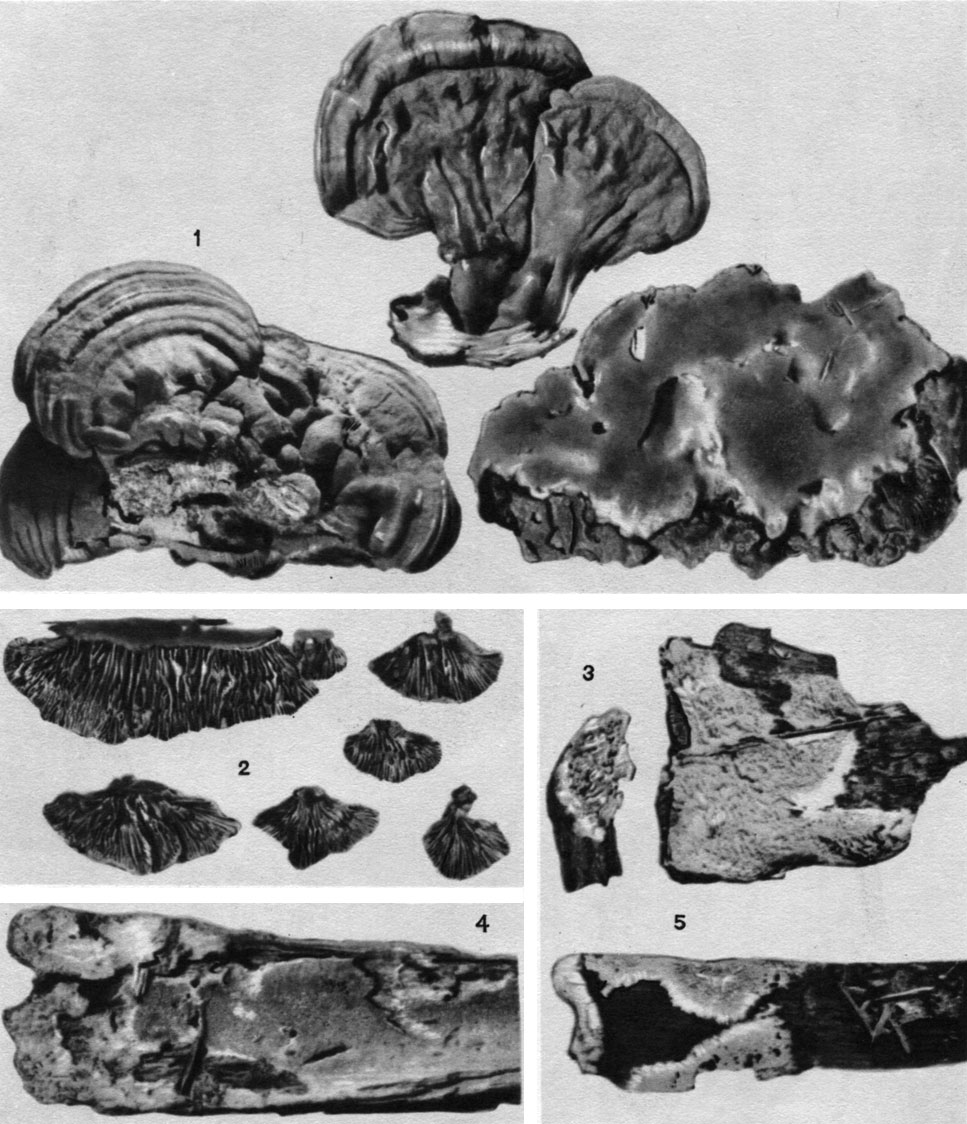
Таблица 28. Плодовые тела трутовых грибов: 1 - слева - плоский трутовик (Ganoderma applanatum); вверху - ганодерма смолистая (G. resinaceum); справа - феллинус черноиолосный (Phellinus nigrolimitatus); 2 - глеофиллум пихтовый (Gloeophyllum abietinum); 3, 5 - фибулопория кремовая (Fibuloporia cremea); 4 - фибулопория маипа (Fibuloporia mappa)
Фибулопория маппа (F. шарра) обитает на валежных стволах хвойных и лиственных пород. В СССР собран только в Ленинградской области. Встречается изредка в Европе и Северной Америке. Ассоциируется с бурой кубической или волокнистой гнилью, пронизанной пленочками мицелия (табл. 28).
Род глеопорус (Gloeoporus) объединяет несколько видов.
Глеопорус двухцветный (Gloeoporus dichrous) обитает на гнилых пнях и крупном валежнике лиственных пород, преимущественно березы, осины, ольхи, дуба, бука, изредка на хвойных или на плодовых телах трутовиков. Встречается этот гриб редко, в основном на территории европейской части СССР и на Кавказе. Вне СССР известен в Западной Европе. Вызывает белую волокнистую гниль. Гниение неактивное.
Глеопорус бахромчатокрайний (G. pannocinctus) обитает на валежнике древесины лиственных пород: ольхе, грабе, липе, клене, буке и некоторых других, изредка на хвойных. Встречается чаще в пойменной зоне, но вообще довольно редок. В СССР известен в Прибалтике, на Северо-Западе, на Кавказе. Вне СССР известен в Западной Европе.
Грибы рода фомитопсис (Fomitopsis) живут на мертвой древесине, иногда на живых деревьях.
Корневая губка (Fomitopsis annosa) обитает на хвойных, изредка лиственных породах (ольхе, березе, буке), на корнях, выступающих из земли, в комлевой части ствола, часто на живых деревьях. В СССР распространен повсеместно (обильно либо редко). Вызывает пеструю напенную гниль хвойных пород (сосны, пихты, реже ели и кедра). Корневая губка - опасный патоген, вызывающий массовое отмирание деревьев.
Окаймленный трутовик (F. pinicola) - многолетний гриб. Он широко распространен в качестве сапрофита на многих лиственных и хвойных породах в умеренной климатической зоне. Изредка этот трутовик является раневым паразитом ослабленных деревьев. Он довольно активен и может вызвать сплошную гниль.
Розовый трутовик (F. rosea) поселяется на отмерших стволах и пнях почти исключительно хвойных пород, из лиственных отмечен на черешне и вязе. Распространен он по всей территории Союза, но преимущественно в европейской части.
В Восточной Сибири и на Дальнем Востоке широко распространен другой, близкий к этому виду - трутовик Каяндера (F. cajanderi), который вызывает бурую гниль.
Грибы рода кориолус (Coriolus) поселяются на лиственных породах, резупинатные формы - на хвойных породах.
Белый домовый гриб (Coriolus sinuosus var. vaporarius) обитает на пнях, стволах и древесине хвойных пород, чаще всего сосны. Он широко распространен как опасный домовый гриб в постройках и вызывает бурую деструктивную гниль. При этом гниение очень активное.
Кориолус красно-бурый (С. cervinus) поселяется на валежнике различных лиственных пород: дубе, ясене, грецком орехе, шелковице, вишне, яблоне, тополе, осине, березе, вязе и других, как исключение - на хвойных. Этот трутовик распространен по всей европейской части СССР и на Кавказе, но преимущественно в южных районах, изредка в Сибири. Он вызывает белую гниль. Гниение активное.
Кориолус волосистый (Coriolus hirsutus) встречается как сапрофит на многих лиственных породах, а как исключение - на ели и пихте. Космополит. Гниль этого трутовика белая, кольцевая. Пораженная грибом древесина распадается на тонкие пластинки по годичным слоям.
Кориолус многоцветный (С. versicolor, см. рис. 164), встречается как сапрофит на многих лиственных породах, в исключительных случаях - на ели. Космополит. Вызывает белую, активно развивающуюся гниль. Этот трутовик типичен на дровах и заготовленной древесине.
Грибы рода дедалея (Daedalea) обитают на древесине.
Дубовая губка (Daedalea quercina, рис. 166) - многолетний гриб. Он поселяется на пнях и мертвых стволах широколиственных пород (дуба, бука, каштана), а также на обработанной древесине. Встречается дубовая губка повсеместно как один из самых обычных грибов. Вызывает активно развивающуюся бурую гниль.

Рис. 166. Дубовая губка (Daedalea quercina). Плодовое тело
Дедалея бугристая (D. confragosa) обитает на различных лиственных породах, особенно иве, березе, ольхе, орешнике. Обычно этот гриб встречается в умеренной зоне европейской части СССР и в Сибири, распространен практически по всей нашей стране. В европейской части преобладает f. rubescens, в Сибири - f. sibirica, на Кавказе и Дальнем Востоке - var. tricolor, хотя возможно во всех этих районах нахождение также других форм и разновидностей. Вне СССР этот трутовик довольно широко распространен в северном полушарии. Он вызывает белую гниль.
Дедалея Диккенса (D. dickensii, рис. 167) обитает на сухостое и валежнике дуба монгольского и березы даурской. Нередко этот гриб встречается в Приморском крае (Дальний Восток). Он вызывает у деревьев белую волокнистую гниль.
Рис. 167. Дедалея Диккенса (Daedalea dickensii). Гименофор
Грибы рода глеофиллум (Gloeophyllum) поселяются на древесине хвойных и лиственных пород.
Заборный гриб (Gloeophyllum sepiarium) обитает на пнях, валежных стволах и ветвях различных хвойных пород, а также на обработанной древесине, особенно в сооружениях открытого типа (мостах, заборах, сваях, шпалах и т. д.). Он может быть занесен на склады и там развиваться как домовый гриб. В виде исключения встречается на лиственных породах (осине). Распространен этот трутовик повсеместно, преимущественно в северном полушарии. Он вызывает бурую, активно развивающуюся гниль.
Глеофиллум пихтовый (G. abietinum) поселяется на древесине хвойных пород, чаще на ели и пихте. Он распространен практически по всей нашей стране, но встречается редко. Вне СССР гриб найден в умеренной и субтропической зонах Старого Света. Этот глеофиллум вызывает бурую, активно развивающуюся гниль. Он встречается в шахтах и подвалах, на обработанной древесине хвойных (табл. 28).
Глеофиллум бревенчатый (G. trabeum) живет на пнях, валежнике и обработанной древесине лиственных, изредка хвойных пород. В СССР встречается редко, но известен для всех районов, кроме Средней Азии и Дальнего Востока. Кроме того, этот гриб встречается в умеренной зоне северного полушария. Он вызывает бурую деструктивную гниль.
Семейство Гименохетовые (Hymenochaetaceae) (М. А. Бондарцева)
Плодовые тела резупинатные, распростерто-отогнутые, сидячие, вееровидные или дифференцированные на шляпку и ножку. Ткань буро окрашенная, чернеющая под действием щелочей (КОН). Гифальная система моно- или димитическая. Пряжки на генеративных гифах отсутствуют. В гимении большинства видов имеются щетинки. Споры часто толстостенные, с гладкой оболочкой, бесцветные или окрашенные.
Гименохетовые грибы встречаются на древесине, а в виде исключения - на почве.
Грибы рода феллинус (Phellinus) поселяются на древесине и часто являются патогенами для живых деревьев.
Ложный трутовик (Phellinus igniarius) обитает на живых деревьях, отмерших стволах и пнях большинства лиственных пород, особенно часто на березе, иве, ольхе, тополе.
Ложный трутовик разделяют на ряд специализированных форм, различающихся по макроморфологическим признакам, приуроченности к определенной породе, незначительным микроскопическим особенностям. Возможно, некоторые из этих форм являются самостоятельными видами, как выделенные ранее Р. tremulae (на осине), P. robustus (на дубе), P. hartigii (на пихте). Этот гриб встречается практически повсеместно и относится к числу самых обычных грибов. Нередко он развивается на живых стволах и вызывает активно развивающуюся белую гниль, пронизанную черными линиями (табл. 29).

Таблица 29. Трутовики: вверху - настоящий (Fomes fomentarius); внизу - ложный (Phellinus igniarius)
Феллинус ржаво-бурый (P. ferrugineofuscus) встречается на валежных стволах и ветвях хвойных пород, особенно пихты и ели. Встречается гриб по всей территории СССР, в европейской части - редко, а в горных районах Сибири довольно обычен. Вызывает желтоватую гниль с расслоением древесины по годичным слоям.
Трутовик Гартига (P. hartigii) растет на хвойных породах, особенно на пихте, реже - на ели, сосне и тисе.
Распространен этот трутовик восточнее Урала, до Сахалина включительно, довольно обычен на Кавказе. В европейской части СССР очень редок: найден только в Карпатах и в Ленинградской области. Очень близок к развивающемуся на дубе P. robustus, отличаясь от него главным образом субстратом и наличием прослоек стерильной ткани между слоями трубочек. Трутовик Гартига вызывает коррозионную бледно-желтую гниль, отграниченную узкими черными линиями от здоровой древесины. Этот гриб - опасный патоген пихты.
Феллипус чернополосный (P. nigrolimitatus) обитает на толстых валежных стволах и пнях хвойных пород, иногда на обработанной древесине. Изредка встречается в европейской части СССР и на Кавказе, чаще - в горных районах Сибири. Этот гриб вызывает пеструю гниль коррозионного типа, с пленками рыжеватой грибницы между годичными слоями пораженной древесины (табл. 28).
Грибы рода инонотус (Inonotus) поселяются на живых стволах и валеже, преимущественно крупном, лиственных и хвойных пород.
Щетинистоволосый трутовик (Inonotus hispidus) обитает на живых стволах многих деревьев, особенно грецкого ореха, платана, шелковицы, дуба, ясеня, вяза и других, начиная с середины лета. Встречается этот трутовик в южных районах (от Курской и Воронежской областей), особенно в Крыму, на Кавказе, в республиках Средней Азии, в Казахстане, кроме того, он найден в южных районах Западной Европы и Северной Америки, а также в Северной Африке. Вызывает белую гниль, отделяющуюся черными линиями от непораженных участков древесины. Плодовые тела возобновляются ежегодно. Особенно часто встречается в садах и парках, причиняя существенный вред. В Средней Азии пигменты плодовых тел используются как красители в ручном ткачестве.
Трутовик лучевой (I. radiatus) обычно поселяется на пнях, сухостое и валежнике ольхи, березы, ивы и других лиственных пород. Трутовик лучевой - широко распространенный вид. Он вызывает белую смешанную (центральную и периферическую) гниль. Загнившая древесина становится волокнистой, пронизанной буроватыми пленками и прожилками мицелия по годичным слоям.
Инонотус Хейнриха (I. heinrichii) встречается на валежных стволах и пнях хвойных пород, особенно пихты, ели и лиственницы. Трутовик распространен на Урале и в Сибири, очень редко встречается в Северной Европе. В Средней Сибири развивается преимущественно в виде скрытой гнили с образованием хламидоспор внутри пораженных стволов; на Алтае часто образует плодовые тела. Вызывает скрытую, активно развивающуюся, белую, волокнистую гниль.
Семейство Ганодермовые (Ganodermатасеае) (М. А. Бондарцева)
Эти грибы обитают на древесине.
Род ганодерма (Ganoderma) объединяет несколько видов трутовиков.
Лакированный трутовик (Ganoderma lucidum, см. рис. 165) развивается у основания ослабленных деревьев, на сухостое и пнях дуба, бука, каштана, ореха, клена, ясеня, ольхи, реже ели и некоторых других пород. Этот гриб распространен почти повсеместно, но преимущественно в южных районах, особенно на Кавказе и в Средней Азии, причем в каждой климатической зоне проявляет наибольшую приуроченность к определенной породе. Гниль, вызываемая трутовиком, белая, неактивная.
Ганодерма смолистая (Ganoderma resinaceum) развивается на живых стволах, пнях и сухостое различных лиственных деревьев (дубе, буке, иве, ольхе) или на хвойных (лиственнице, секвойе). В СССР встречается редко, преимущественно в горных лесах Кавказа, Карпат, Алтая и на Дальнем Востоке. Вне СССР распространен в субтропиках и тропиках. Ассоциируется с белой гнилью (табл. 28).
Плоский трутовик (Ganoderma applanatum) обитает на пнях и мертвой древесине многих лиственных деревьев (особенно часто на березе) и хвойных пород, очень редко встречается на живых деревьях. Этот трутовик - космополит. Он вызывает белую или желтоватую коррозионную гниль корней и оснований стволов (рис. 163, табл. 28).
Семейство Полипоровые (Polyporaceae) (М. А. Бондарцева)
Плодовые тела с дифференцированной боковой, центральной или эксцентрической ножкой. Ткань белая или очень слабо окрашенная. Гименофор трубчатый или ячеистый. Гифальная система димитическая. Споры бесцветные, с гладкой оболочкой.
Эти грибы живут на древесине.
Род полипорус (Polyporus) объединяет несколько видов трутовиков.
Чешуйчатый трутовик - пестрец, или заячник (Polyporus squamosus), встречается на живых и мертвых стволах широколиственных пород в умеренной зоне всего земного шара. Этот трутовик вызывает белую центральную гниль, иногда опасен как раневой паразит. Плодовые тела гриба съедобны.
Полипорус изменчивый, или трутовик изменчивый (P. varius), обычно растет с середины лета до поздней осени на пнях, сухостое и валежнике березы, ольхи, ивы, липы, дуба, ясеня, яблони и других лиственных пород. Изредка встречается на живых деревьях. Обычен в лесах умеренной зоны, встречается в старых парках и садах. Гриб вызывает белую, активно развивающуюся гниль. Известно несколько форм и разновидностей этого очень изменчивого вида, из которых наиболее обычна var. et f. elegans.
Полипорус черноногий (P. meianopus) растет на старом валежнике, опавших ветвях или сухих корнях (прикрытых землей) различных лиственных пород (березе, ольхе, дубе и др.). Как исключение, обитает на хвойных (пихта). Распространен по всем умеренным областям СССР до Дальнего Востока включительно, но встречается редко. Описано много форм этого гриба, некоторые из них значительно отличаются по внешнему виду от основной, более всего напоминая P. ciliatus и отличаясь от него главным образом черной ножкой. Вызывает белую, активно развивающуюся гниль.
Семейство Скутигеровые (Scutigeraceae) (М. А. Бондарцева)
Плодовые тела однолетние, одно- или многошляпочные, иногда сидячие, вееровидные, одиночные, срастающиеся краями шляпок или черепитчатые, с боковой, эксцентрической или центральной, иногда рудиментарной ножкой. Ткань белая или светлая, иногда принимающая на изломе розоватый или зеленоватый оттенок. Гифальная система моно- или димитическая со связывающими гифами. Споры бесцветные, с гладкой оболочкой. Грибы поселяются на корнях и стволах живых деревьев или на почве.
Род летипорус (Laetiporus) представлен грибами, обитающими в основном на древесине, преимущественно на живых стволах.
Серно-желтый трутовик развивается в первую половину лета (с мая) на живых, реже мертвых стволах лиственных (иногда хвойных) пород. Космополит. В европейской части СССР преобладает на дубе, в Сибири - на лиственнице, на Дальнем Востоке - на орехе маньчжурском. Этот трутовик вызывает бурую, призматическую, быстро распространяющуюся, центральную гниль. Молодые плодовые тела съедобны.
Грибы рода алъбатреллус (Albatrellus) обитают на почве.
Овечий трутовик (Albatrellus ovinus) развивается в конце лета и осенью на почве в хвойных лесах, чаще на полянах, опушках, просеках, вырубках. Широко распространен этот трутовик по всей умеренной зоне СССР, особенно в подзоне северной и средней тайги, но встречается обычно нечасто. Растет он, как правило, большими группами, так что может быть собран и как съедобный гриб. Относится к числу грибов невысокой ценности, но, поскольку он растет и в позднеосеннее время, может представлять интерес при отсутствии других грибов.
Алъбатреллус сливающийся (A. confluens) обычно находят в конце лета и осенью на почве в хвойных лесах, преимущественно в северных районах. Встречается редко, но большими группами, так что может быть использован как съедобный гриб. От предыдущего вида отличается главным образом оттенком ткани на изломе и при подсыхании.
Семейство Болетопсидные (Boletopsidaceae) (М. А. Бондарцева)
Плодовое тело с дифференцированной центральной или эксцентрической ножкой, мясистое, однолетнее. Гифальная система мономитическая. Споры дымчатые, угловатые, неамилоидные. Растет на земле. В семействе только один род.
Род болетопсис (Boletopsis) обитает на земле. В роде всего один вид.
Болетопсис малочешуйчатый (Boletopsis subsquamosa) обитает на перегнойной почве в хвойных (преимущественно сосновых) лесах во второй половине вегетационного периода. Широко распространен, хотя встречается единично. Циркумполярный вид.
Семейство Бондарцевиевые (Bondarzewiaceae) (М. А. Бондарцева)
Плодовые тела крупные, вееровидные, соединенные основаниями. Ткань белая или светлая. Гифальная система мономитическая. Генеративные гифы без пряжек. Споры бесцветные, с амилоидными возвышениями в виде шипов или валиков. Обитают на корнях живых деревьев. Семейство включает всего один род.
Род бондарцевия (Bondarzewia) соответствует диагнозу семейства.
Бондарцевия горная (Bondarzewia montana) поселяется у основания или на корнях пихты. Встречается очень редко, преимущественно в горных лесах более или менее южных районов; в СССР - в Закарпатье и на Кавказе. Гниль на раннем этапе светло-бурая, затем с многочисленными узкими пустотами, в последней стадии - трухлявая, волокнистая.
Семейство Кортициевые грибы (Corticiaceae) (Э. Х. Пармасто)
Плодовые тела этого очень крупного семейства имеют весьма простое макроскопическое строение. Нередко это только тонкая распростертая гладкая пленка на нижней стороне валежного ствола. Толщина плодового тела от 30-40 мкм у самых тонких, в сухом состоянии практически невидимых представителей, до нескольких миллиметров у кожистых видов, которые нередко живут до десятка лет и постепенно утолщаются. Консистенция плодового тела паутинистая, мясистая, кожистая или даже деревянистая. Плодовые тела в свежем виде иногда слизистые или желатинозные, а в сухом состоянии твердо-роговидной консистенции. Одни грибы крепко прикрепленные, другие - легко отделимые от субстрата; одни с мучнистым краем, другие с радиально-волокнистым или даже снабженные разветвленными грибными шнурами.
В окраске плодовых тел встречаются всевозможные оттенки желтого, охряного, светло-бурого; нередко они белые или беловатые, реже розовые, лиловые, красные, как исключение темно-синие или зеленые.
Гименофор у большинства видов гладкий, у других - бородавчатый или шиповатый, реже складчатый, как исключение встречаются виды с короткотрубчатым (пористым) гименофором.
Макроскопическое строение плодовых тел настолько однородное, что в старых системах различали только несколько родов, которые включали в большое искусственное семейство телефоровых (Thelephoraceae). В эту группировку соединяли те афиллофоровые грибы, которые имеют гладкие распростертые плодовые тела. С другой стороны, многие кортициевые грибы были по признакам внешнего строения включены в другие, чужие семейства грибов (ежовиковые, трутовые).
Признание важности микроскопических признаков в систематике афиллофоровых грибов привело к полной перестройке таксономии кортициевых грибов. Было обнаружено, что в плодовых телах наблюдаются различные типы гиф; что можно различить десятки своеобразных типов цистид и гифид - клеток, которые вместе с базидиями образуют гимениальный слой. Было обнаружено и несколько типов базидий.
"Нижняя" часть плодового тела, прилегающая к субстрату, называется подстилкой или субикулюмом. (Фактически это не нижняя, а верхняя часть: плодовые тела кортициевых грибов имеют положительный геотропизм, т. е. всегда обращены гимением вниз.) Подстилка состоит из плотного или рыхлого сплетения генеративных гиф, которые у многих видов и родов снабжены пряжками. Различия в химическом составе, в оптической плотности, в типе разветвления гиф - важные для систематики признаки. Под подстилкой наблюдаются более плотно сплетенные тонкостенные гифы - субгимений. На этом образуется палисадный слой базидий, между которыми обыкновенно в большом количестве наблюдаются базидиолы (недоразвитые базидии), гифиды (окончания гиф), а нередко и цистиды самой различной формы (рис. 159). Цистиды могут быть тонко- или толстостенные, по форме - от шиловидных до шаровидных; нередко они инкрустированы кристаллами или смолистым веществом; у некоторых родов цистиды с выростами. Различают глеоцистиды, которые всегда тонкостенные и с мучнистым или мелкокапельчатым содержимым.
У примитивных представителей кортициевых грибов базидии 6-8-споровые. Предполагают, что это показывает на общие с сумчатыми грибами предки афиллофоровых грибов. В большинстве родов базидии 4-споровые, а изредка (у определенных видов) 2-споровые. Количество спор на базидии - весьма характерный для данного вида признак. Тем не менее у большинства исследованных в этом отношении видов среди 4-споровых базидий почти всегда встречаются совсем немногочисленные 2-споровые и исключительно редкие - 1-споровые базидии.
Споры обыкновенно бесцветные, тонкостенные, гладкие, неамилоидные, от цилиндрических или колбасковидных до шаровидных, величиной 3-20 мкм. Имеются и исключения: некоторым родам характерны или шиповатые, или толстостенные, или амилоидные споры.
Кортициевые грибы широко распространены в природе. Большинство видов - лигнофилы: они обитают на валежных стволах и ветках, на пнях, в лесной подстилке, на заготовленной и обработанной древесине. Обыкновенно они малозаметные, у большинства видов и гниение древесины под влиянием гриба протекает медленно. Тем не менее благодаря обилию видов и при-способленности к самым различным местообитаниям кортициевые грибы в общей сложности играют огромную роль в круговороте веществ и энергии в лесных фитоценозах. Вызываемая ими гниль может быть как бурой, так и бе-лой, ибо они вызывают разложение и целлюлозы, и лигнина. Большинство видов - разрушители лигнина, но немало видов - целлюлозоразрушителей. Наряду с сапрофитами имеются виды, способные заражать живые деревья, а также несколько паразитов травянистых растений.
Кортициевые грибы распространены по всему миру, больше их, по-видимому, в неморальных и смешанных лесах умеренных поясов обоих полушарий. В Советском Союзе обнаружено около 375 видов.
Наиболее распространенный паразит среди кортициевых грибов - танатефорус огурцовый (Thanatephorus cucumeris). Плодовые тела этого гриба появляются как нежный белый налет на пораженных стеблях и листьях картофеля, люпина, капусты, некоторых сорняков полей и других травянистых растений. Налет состоит из немногочисленных рыхлоперепутанных бесцветных гиф подстилки и разбросанных на них 4-споровых базидий. Этот гриб известен в фитопатологической литературе под неправильным названием Hypochnus solani и является базидиальной стадией развития известного паразита Rhizoctonia solani, вызывающего ризоктониоз целого ряда культивируемых растений.
Род трехиспора (Trechispora) отличается шиповатыми спорами и большим варьированием строения гименофора (табл. 30). Рядом с видами, имеющими гладкий гименофор, встречаются и другие, с шиповатым гименофором (например, Т. farinacea), которые еще недавно на основе только этого признака считали представителями совсем другого семейства - ежовиковых грибов.

Таблица 30. Плодовые тела афиллофоровых грибов: вверху - трехиспора (Trechispora); внизу слева - рогатик (Clavaria); внизу справа - трубчатый гименофор
В таежных лесах и даже в лесотундре у нас нередко встречается на очень гнилой древесине трехиспора белоснежная (Т. candidissima) - гриб, который внешне не отличается от распростертых трутовых грибов.
Некоторые кортициевые грибы - настоящие почвенные грибы, которые очень обильно растут в гниющей лесной подстилке. Если их почти никогда не отметят в исследованиях по почвенной микробиологии, то в этом виновата применяемая стандартная методика, малопригодная для обнаружения гименомицетов. Своеобразие почвенных кортициевых состоит еще в том, что их плодовые тела образуются на нижней стороне валежных остатков древесины, в ходах кротов и мышей, а иногда даже под маленькими камнями.
Один из таких видов - ателия тонконитчатая (Athelia byssina). Многочисленные белые грибные шнурочки этого вида нередко можно найти в подстилке под моховым покровом в лесах, где реакция почвы почти нейтральная. Еще чаще можно в подстилке кислой реакции хвойных лесов (особенно сосновых боров) видеть шафранно-желтые шнурочки близкого вида - A. bicolor.
На валежных стволах сосны и ели уже через полгода или год после бурелома возникают широко простирающиеся плодовые тела флебии гигантской (Phlebia gigantea). Это восковатые, в сухом состоянии хрящеватые, молочно-белые или сероватые, в краях радиально-волокнистые пленки с неровным гимением, которые при высыхании иногда по краям заворачиваются. Гниющая древесина приобретает желтую или серую окраску, позднее образуются мелкие пустоты. Гриб поражает нередко неокоренные лесоматериалы, иногда встречается как домовый гриб. Поэтому его рассматривают в справочниках и учебниках по лесной фитопатологии как вредный гриб (обыкновенно под названием Peniophora gigantea). Но в последние годы обнаружены некоторые весьма полезные свойства флебии гигантской. Как известно, наиболее опасным для сосны и ели грибом в наших лесах является корневая губка (трутовый гриб Fomitopsis annosa), который может распространяться образующими на зараженных пнях конидиями. Флебия гигантская - антагонист корневой губки, зараженные флебией пни уже недоступны для корневой губки. Сделаны первые удачные попытки применения этого антагонизма; мазанные суспензией спор флебии пни сосны и ели заражаются флебией и не могут уже больше служить возможным источником распространения корневой губки. Следовательно, флебия - гриб, вредный в лесах, где не удаляют вовремя валеж и валеный лесоматериал; там, где лесное хозяйство на должном уровне, гриб полезен как антагонист опасного заболевания леса.
На ветках дубов нередко встречается почти невидимый кортициевый гриб Vuilleminia соmedens, который нередко вызывает массовое высыхание веток. Кора веток отпадает, и из-под шелушащегося эпидермиса выступает плодовое тело - тонкая желтоватая или сероватая пленка, при влажной погоде слегка слизистая, при сухой погоде едва отличающаяся от древесины. Гриб вызывает белую гниль древесины; интересен он тем, что у его плодовых тел много десятков раз в год чередуются период интенсивного спорообразования и период покоя. При сухой погоде жизнедеятельность гриба ослабляется, а через несколько часов после промокания ветки начинается обильная споруляция. В любое время года можно собирать сухие ветки дуба с плодовыми телами этого гриба. Если их мочить полчаса в теплой воде, дать четверть часа подсохнуть и положить на предметное стекло или черную бумагу, то получим уже через 4-8 ч обильный белый налет спор, пригодных для демонстрации или опытов. Поэтому этот гриб - очень подходящий объект для многих, особенно экспериментально-экологических, исследований, тем более что у него крупные споры (длиной до 20 мкм).
Как мы видим, Vuilleminia comedens - настоящий ксерофильный гриб. Такие же свойства характерны и для видов рода пениофора (Peniophora), которые встречаются на высохших ветках в кроне деревьев и кустарников и которые особенно характерны для растительности средиземноморского типа. Ксероморфное строение позволяет некоторым видам обитать в суровых условиях северной тайги и лесотундры. Плодовые тела многолетние, утолщающиеся ежегодно; в гимении и в подстилке многочисленные инкрустированные толстостенные цистиды, а нередко и крупные глеоцистиды; гифы так плотно склеены, что при микроскопировании почти неразличимы. В СССР найдено 24 вида. Интересна история изучения пениофоры сосновой (P. pini). Еще недавно этот вид считали весьма редким; сейчас известно, что гриб предпочитает образовывать свои плодовые тела на свежевысохших ветках сосны на высоте 5-10 м и больше от земли.
Плодовые тела лиловатые, но маленькие, меньше 1 см в диаметре, и на валежных деревьях они скоро погибают. Возможно, этот очень распространенный вид может вызывать высыхание веток, но изучать распространение и биологию этого вида в природных условиях весьма затруднительно.
К кортициевым грибам относят и кирпично-красный, а затем черно-пурпуровый распростертый трутовый гриб Meruliopsis taxicola (табл. 31).

Таблица 31. Афиллофоровые грибы: вверху - мерулиопсис тисовый (Meruliopsis taxicola); внизу - стереум жестковолосый (Stereum hirsutum)
Мерулиопсис тисовый (Meruliopsis taxicola) обитает на сухих, не опавших еще валежных ветвях и древесине хвойных пород, чаще сосны. Этот вид обнаружен во всех районах СССР, но встречается редко. Вне СССР известен в Западной Европе, Северной Америке и Австралии.
По хозяйственному значению этот гриб - слабый разрушитель древесины, изредка встречающийся в качестве домового гриба.
Некоторые кортициевые участвуют как грибной компонент в составе базидиолишайников. Это - виды рода диктионема (Dictyonema), найденные в тропиках и субтропиках. В отличие от всех других лишайников один вид из рода диктионема может комбинироваться с несколькими видами водорослей, образуя различные "виды" лишайников.
Семейство Кониофоровые (Coniophoraceae) (Э. Х. Пармасто)
Кониофоровые - небольшая группа грибов с распростертыми темно-коричневыми или бурыми пленчатыми или почти мясистыми плодовыми телами. В родах кониофора (Coniophora) и кониофорелла (Coniophorella) гименофор неровный, неправильно бугорчатый или почти гладкий; гименофор видов рода серпула (Serpula) складчатый, лабиринтовидно-сетчатый или почти пористый. Тесно расположенные складки образуют иногда короткие трубочки, которые отличаются от трубочек трутовых грибов только покрытыми гимением (фертильными) краями.
Из микроскопических признаков - наиболее своеобразное строение спор: они толстостенные, темно-желтые или буроватые и окрашиваются сильно в препаратах, в которые прибавлен реактив "Коттон блу" (хлопчатобумажное синее). Споры прорастают в мицелий, который проникает в древесину и вызывает интенсивное деструктивное гниение. Гриб разрушает целлюлозу, оставляя почти незатронутым лигнин; древесина становится бурой, растрескивается многочисленными трещинами в кубики, которые при надавливании (например, между пальцами) превращаются в мелкий по-рошок.
Самый распространенный из видов семейства - кониофора обыкновенная (Coniophora puteana, табл. 32), которая в старой литературе известна под названием С. cerebella. В лесах таежной зоны нередко можно летом видеть на пнях и валежных стволах хвойных (реже - лиственных) пород вначале белые, затем желтые, а осенью коричнево-бурые пленчатые плодовые тела; со временем средняя часть их становится неровной и покрывается слоем зрелых базидиоспор. Зараженные пни разрушаются, своевременно не вывезенные лесоматериалы теряют свою годность. Споры образуются до глубокой осени и переносятся ветром на значительные расстояния. Нередко этот гриб встречается в сооружениях и постройках: на старых деревянных столбах, на нижней стороне перекрытий крыш, а иногда и как опасный домовый гриб в подвалах и т. д. Для грибов рода серпула характерен складчатый гименофор.

Таблица 32. Афиллофоровые и гетеробазидиальные грибы: вверху слева - рамария Инвала (Ramaria invalii); вверху справа - клавариадельфус ситниковый (Clavariadelphus junceus); в середине слева - серая лисичка (Craterellus cornucopioides); в середине справа - кониофора обыкновенная (Coniophora puteana); внизу слева - дакримицес деликвесценс (Dacrymyces deliquescens); внизу справа - калоцера вискоза (Calocera viscosa)
Большинство из найденных в СССР видов рода серпула встречается в лесах, обыкновенно на древесине хвойных, и то редко. В Предкавказье найден очень редкий вид - серпула лесная (Serpula silvester), которая встречается и в Средней Европе. Некоторые исследователи считают ее диким предком настоящего домового гриба.
Настоящий домовый гриб (Serpula lacrymans) - наиболее вредоносен из всех домовых грибов и наиболее распространен на территории СССР. Он встречается от Прибалтики до Камчатки, известен в Западной Европе и в Северной Америке.
Настоящий домовый гриб - один из тех немногих видов грибов, которые встречаются только в постройках и в природе не наблюдаются. Гриб заражает преимущественно деревянные части нижнего этажа и подвала: полы, стены до высоты около 1 м. Первыми признаками заражения являются затхлый запах в помещениях, невысыхающие, мокрые пятна на стене, а затем и шатающиеся доски пола. При раскрытии пола можно обнаружить ватообразные скопления мицелия гриба, грибные шнурочки и пленки. Вначале они белые, а затем с характерным желтоватым, розоватым, лиловатым и серым оттенком. Часть древесины уже совсем сгнила, часть окрасилась в бурый и стала мягкой.
Гниль распространяется в древесине очень быстро; этому способствует рост грибных шнурков и пленок по поверхности зараженной древесины. Для полного загнивания пола или балки требуется до года, а иногда даже полгода. На нижней поверхности гнилой древесины образуются мясисто-пленчатые плодовые тела, которые могут достигать значительных размеров, длиной до нескольких метров. В сырых помещениях плодовые тела появляются и в нижней части стен внутри комнаты, у основания косяков и т. д. Наличие таких, видимых внутри самого помещения плодовых тел - признак широкого распространения и сильного повреждения деревянных конструкций.
На плодовых телах образуются в массовом количестве базидиоспоры гриба, которые иногда видны в виде коричневой пыли на полу или в углах помещения. На 1 см2 поверхности плодового тела в течение суток может выделиться до 35 млн. спор; наиболее интенсивная споруляция весной и в первой половине лета.
Споры настоящего домового гриба очень легкие. В 1 м3 воздуха в зараженном помещении насчитывается до 2,5 млн. спор. Они переносятся даже небольшим движением воздуха; в воздухе любого города или поселка почти всегда имеются споры настоящего домового гриба. Споры можно случайно перенести на обуви и одежде людей. Особенно опасны зараженные грибом доски и прочие гнилые остатки древесины, которые при ремонте нередко бросают во двор или даже приносят как топливо в незараженные дома.
Для прорастания и дальнейшего заражения древесины споры требуют определенных условий. Это прежде всего большая влажность древесины и окружающего воздуха. Наиболее интенсивное гниение наблюдается при влажности воздуха 90-95% и температуре 18-23 °С. Такие условия могут возникнуть при неправильном строительстве здания, когда недостаточно хорошо проложена изоляция деревянных конструкций от фундамента. Часто наблюдают накопление конденсационной влаги, например около водопроводных труб; попадание дождевой или талой воды в подвалы, под стены. Многие простые деревянные постройки в Сибири и на севере Дальнего Востока построены с цоколем, который служит для теплоизоляции нижней части постройки. Если в наполнитель цоколя будет попадать вода (дождевая), то заражение домовыми грибами почти неизбежно.
Вторая группа причин заражения - небрежность в эксплуатации домов. Несвоевременный ремонт крыши, неисправная вентиляция в сырых помещениях, протечь водопровода и прочие недостатки в уходе приведут к постоянному накоплению сырости и при этом создаются условия, благоприятствующие развитию домовых грибов. При заражении построек домовым грибом необходим срочный ремонт. Опоздание с ремонтом даже только на месяц или два может увеличить потери в несколько раз.
При ремонте следует полностью удалять деревянные конструкции, зараженные грибом. Начальная стадия развития гнили незаметна для невооруженного глаза, поэтому приходится удалять и видимо здоровые доски, граничащие с явно гнилой: вдоль волокон древесины до 0,5 м, а поперек волокон не менее 20 см. Тщательно надо убрать и верхний слой под зараженным полом: гриб может еще долго сохраняться и развиваться в виде мицелия в сырой почве или даже в песке, если там имеется примесь опилок или гумуса.
Зараженные конструкции следует заменить новыми из сухого антисептированного дерева или из негниющего материала. Новый пол нельзя сразу покрывать плотным покрытием (линолеум, релин и другие синтетические материалы), которое препятствует сушке пола. Но главное при ремонте - обнаружить и ликвидировать причины, которые благоприятство-вали развитию домового гриба, создав нужные для его развития условия (сырость, накопление влаги).
Семейство Стереовые (Stereасеае) (Э. Х. Пармасто)
Плодовые тела грибов небольшого семейства распростерто-отогнутые, вееровидные, реже - совсем распростертые, тонкие, всегда кожистой консистенции и с гладким гименофором. Поверхность шляпки почти всегда короткоопушенная, различных оттенков серого, желтого, коричневого (табл. 31 и рисунок 168).

Рис. 168. Стереум (Stereum insignytum)
Все грибы, встречающиеся на территории Советского Союза, с более или менее выраженными ксеробионтными свойствами. Они обитают на мертвой древесине - на пнях, валежных стволах и сучьях, на дровах - по всей лесной зоне; в небольшом количестве - в лесостепи. И даже в парковых насаждениях или в зарослях кустарников в зоне полупустынь. Они выносливы к высыханию и не боятся расти даже на очень сухих солнечных местах.
В нашей стране найдено 19 видов семейства стерео вых. Из них некоторые имеют значение в лесном хозяйстве.
Плодовые тела стереума краснеющего (Stereum sanguinolentum) встречаются нередко в таежных лесах на валежных стволах сосны и ели. Они буровато-серые, кожистые, почти распростертые, только кое-где с небольшими отогнутыми шляпками. Но во влажную погоду при легком прикосновении сероватая поверхность гимения становится в местах дотрагивания кровяно-красной. Такая картина только у трех видов рода стереум: тонкая оболочка верхушки цистидиол растрескивается при прикосновении и их желтое содержимое выделяется на поверхность гимения.
Гриб является сапробионтом, но тем не менее вредным. При подсечке старых елей и сосен обнажается древесина, которая может заражаться некоторыми видами лесных грибов.
Среди них на первом месте стоит стереум краснеющий. Гриб вызывает медленное, но все же заметное гниение древесины. Гнилая древесина приобретает вначале буроватый или красноватый оттенок, а затем белеет. Через несколько лет гниль распространяется на глубину до 5 см.
Следует отметить еще стереум жестковолосый (S. hirsutum, табл. 31). Плодовые тела этого гриба появляются в большом количестве на заготовленных из лиственных пород дровах, которые остались в лесу или на складе. Это многочисленные, нередко черепитчато расположенные, тонкие, полукруглые шляпки, сверху волосистые, зональные, серые, а снизу охряно-желтые, иногда с серым налетом. Древесина вначале желтеет, потом белеет; гниение идет довольно быстро, особенно при повышенной влажности воздуха.

Рис. 169. Стереллум сосновый (Sterellum pini)
Семейство Цифелловые (Сyphellaceae) (Э. Х. Пармасто)
Это небольшая группа малозаметных грибов очень своеобразного строения. Плодовые тела их очень маленькие, высотой меньше нескольких миллиметров и еще меньше в диаметре, бокальчатые, чашевидные, бочковидные или почти цилиндрические; прикрепляются суженным основанием или короткой ножкой. Поверхность плодовых тел пушистая или почти гладкая, от белой до бурой. Устье плодового тела нередко с волосками. Внутренняя полость плодового тела гладкая (редко - складчатая), покрыта гимением.
Плодовые тела цифелловых грибов растут иногда одиночно, но чаще небольшими или большими группами сближенно; между ними растет рыхлый паутинистый мицелий, но не всегда.
Плодовые тела можно найти на остатках древесины, на валежных сучках, мертвых стеблях травянистых растений, на отмерших черешках папоротников. Большинство видов встречаются редко или очень редко.
На территории Советского Союза пока обнаружено два десятка видов, но следует учесть, что эта группа грибов осталась у нас почти не изученной.
Еще сравнительно недавно считали, что эта группа грибов довольно хорошо выделяется среди других грибов самостоятельным семейством. Последние исследования показали, что это типичная группа искусственной системы. По-видимому, различные грибы могут в ходе эволюции образовывать виды с упрощенными бочковидными мелкими плодовыми телами, которые способны заселять мелкие остатки растений. Считают, что многие цифелловые грибы - это упрощенные до предела и потерявшие способность образовывать пластинки агарикальные шляпочные грибы (Agaricales). Известный нидерландский миколог М. А. Донк сравнил семейство Cyphellaceae с такой "систематической" группой животных, где вместе представлены кенгуру и все остальные прыгающие млекопитающие.
Большинство цифелловых грибов - мезофиты, обитатели более или менее влажных лесов. Но имеется один вид, который встречается как в лесотундре и тайге, так и в полупустынных насаждениях на мелких отмерших или валежных веточках лиственных пород. Это самый обычный представитель группы - цифеллопсис аномальный (Cyphellopsis anomala). Плодовые тела его чашевидные, с вогнутыми внутрь краями высотой 0,2-1 мм и диаметром до 0,4 мм, прикреплены короткой ножкой, тесно покрытые бурыми волосками, растут густыми группами, образуя пятна длиной до 2-3 см. Вся внутренность плодового тела покрыта плотным слоем базидий, носящих по 4 цилиндрические или почти сосисковидные споры длиной 7-10 мкм. Они кожистые, способны полностью высыхать и оживляться после дождя. В условиях северных районов плодовые тела погибают зимой, а в южных областях могут жить несколько лет. Гриб широко распространен на всех континентах мира.
Второй, при этом очень красивый гриб встречается у нас во влажных лесах на основании отмерших корешков страусопера германского.
Плодовые тела растут небольшими группами или даже по 2-5; они цилиндрические, длиной до 2,5 мм, оранжево-коричневые, у основания почти бурые, около вершины белые, гладкие, с короной желтоватых волосков вокруг устья. Это - волдемария шафранно-желтая (Woldemaria стосеа). Гриб найден в СССР, в Швеции и в Северной Америке и является везде редким.
Семейство Шизофилловые (Schizophyllaceae) (Э. Х. Пармасто)
Плодовые тела у некоторых шизофилловых грибов чашевидные, как у цифелловых. У других в ходе эволюции образовались сложные плодовые тела, которые представляют собой группу сближенных плодовых тел на общей подстилке. В роде строматосцифа (Stromatoscypha) в результате этого образуется трубчатый гименофор, который напоминает настоящий гименофор распростертых трутовых грибов. Отличие его в том, что перегородки трубочек у этих грибов - не что иное, как сросшиеся края маленьких чашевидных плодовых тел. В роде щелелистник (Schizophyllum) плодовые тела (шляпки) округлые, почковидные, прикрепляются спинной стороной или боком; гименофор состоит из радиально расположенных складок. Эти складки - сросшиеся края настоящих плодовых тел. Гифальная система плодовых тел мономитическая, гифы с пряжками. Базидии с (2) 4 спорами; споры тонкостенные, гладкие, неамилоидные.
Щелелистник обыкновенный (Schizophyllum commune) из-за некоторого внешнего сходства с агариковыми грибами еще недавно считали настоящим представителем этой группы. Шляпки этого вида диаметром до 5 см, нередко с лопастными краями, тонкие, кожистые, сверху светло-серые, войлочные. Пластинки вееровидно расположенные, пурпурно-серые. При сухой погоде края пластинок раздваиваются и завертываются так, что края соседних пластинок прикасаются и покрывают гимений. При влажной погоде края выпрямляются и гимений стоит открытым. Такой сложный механизм - показатель своеобразного происхождения "ложных пластинок". Не менее интересен тот факт, что такая защита гимения во время засухи позволяет щелелистнику расти в условиях очень сухого климата, даже в полупустыне. Гриб растет во всех частях света, у нас распространен на валежных стволах, пнях и отмерших ветках лиственных, изредка хвойных пород во всех районах флоры, кроме арктического. Среди густого леса встречается, только редко. Вызываемая грибом гниль развивается медленно; иногда гриб встречается и на живых деревьях около ранений ствола.
Семейство Лахнокладиевые (Lachnocladiaceae) (Э. Х. Пармасто)
Это семейство небольшое, но с очень своеобразным микроскопическим строением. Плодовые тела однолетние или многолетние, распростертые, рогатиковые (клавариоидные) или лопатчатые, более или менее кожистые. Гифы плодового тела дифференцированные в два типа: кроме тонкостенных гиф, образующих в гимении базидии, наблюдаются еще скелетные. Это декстриноидные (окрашивающиеся под действием иода в коричневый) толстостенные гифы, не имеющие перегородок, которые разветвляются очень своеобразно - дихотомически или роговидно, а иногда образуют звездчатые астерощетинки. Споры амилоидные или неамилоидные.
В СССР найдены только грибы (15 видов) с распростертыми плодовыми телами. Во всей лесной зоне Советского Союза, но особенно часто в северной тайге и в лесотундре, встречается варария одевающая (Vararia investiens). На нижней стороне валежных веток и стволов хвойных пород, а иногда и прямо на земле в перегное, на опавшей хвое и других остатках древесных пород образуются плодовые тела, толщиной 0,3-1 мм, имеющие своеобразную бледно-яично-желтую окраску. Иногда такие плодовые тела простираются широко и имеют длину до 2 м. По-видимому, этот гриб является в условиях Крайнего Севера важным разрушителем лесного опада и играет там немаловажную роль в круговороте веществ в природе.
На Дальнем Востоке, а также на Урале найден редчайший во всем мире вид - астерострома средняя (Asterostroma medium). Большинство из его тонких (до 0,5 мм) бледно-коричневых плодовых тел состоит из лучистых астерощетинок.
Семейство Рогатиковые (Clavariaceae) (Э. Х. Пармасто)
Для грибов этого семейства характерны следующие признаки: плодовые тела булавовидные или почти цилиндрические, вертикально вверх растущие или разветвленные с простыми или повторно разветвляющимися ветвями, иногда даже коралловидные. Вся поверхность плодового тела, кроме ножки и обращенных вверх участков поверхности ветвей, покрыта гимениальным слоем. Высота плодовых тел - от нескольких миллиметров до двух десятков сантиметров; консистенция мясистая, кожистая, реже слегка хрящеватая или почти деревянистая. Окраска специфична для каждого вида; многие виды желтые различных оттенков или почти белые, но встречаются оранжевые, лиловые, красные, темно-бурые и др.
Гифальная система плодовых тел мономитическая, реже димитическая (генеративные и реже скелетные гифы). Гифы иногда вздутые, с простыми перегородками или пряжками. Субгимений обыкновенно хорошо развитый; гимений у грибов многих видов утолщающийся, несмотря на то что плодовые тела всегда однолетние и обыкновенно скоро разлагающиеся. У базидий гимения 4 (реже 2) стеригмы. В гимении находятся также базидиолы, изредка цистиды или глеоцистиды. Споры бесцветные или желтые, тонкостенные, гладкие, шиповатые, реже с полосатой или спиральной орнаментацией, неамилоидные.
Почти все виды семейства - сапрофиты на лесной подстилке, на очень гнилой древесине, а также (Pistillaria, Typhula) на гниющих стеблях и листьях травянистых растений. Несколько грибов рода Typhula вызывают заболевания культурных растений и сеянцев в лесных питомниках. Представители рода спарассис (Sparassis) вызывают гниль корней лесных деревьев.
Среди рогатиковых грибов большинство несъедобны из-за консистенции и неприятного вкуса. Некоторые из них слабо ядовиты. Но существует много съедобных грибов, например грибная капуста (Sparassis crispa), представители рода рамария (Bamaria). Приятный горьковатый привкус имеют очень крупные мясистые плодовые тела рамарии желтой (Bamaria flava, рис. 170).

Рис. 170. Рамария желтая (Ramaria flava). Плодовое тело
Биология рогатиковых грибов совсем мало изучена. Предполагают, что некоторые из них - микоризообразователи. Плодовые тела появляются у большинства видов не ежегодно, обычно они живут недолго (несколько недель). Большинство видов редкие или очень редкие. Поэтому даже микологи не всегда найдут их во время своих экспедиций. Но бывает, что некоторые грибы рода рамария развиваются в массовом количестве и привлекают внимание любителей грибов яркой окраской и своеобразными коралловидно-разветвленными плодовыми телами.
При гербаризации плодовые тела рогатиковых изменяются до неузнаваемости. Первоначальная яркая окраска меняется при высыхании в буроватый или даже черный цвет, мясистые плодовые тела становятся совсем хрупкими. Это одна из причин, по которой в систематике рогатиковых еще много спорного. Новейшие систематики различают в группе рогатиковых до 4 самостоятельных семейств, но их границы еще неясны. Много идет споров о родстве рогатиковых и лисичковых грибов. Некоторые известные микологи (Корнер, Крайзель и другие) утверждают, что рогатиковые грибы чуть ли не самые примитивные среди гименомицетов: ведь строение их гименофора (отсутствие шипов, трубочек и пр.) не позволяет образованию большого количества спор; строение плодовых тел не защищает базидии и споры от дождя и других неблагоприятных метеорологических факторов (как это наблюдается у трутовых и ежовиковых грибов).
Все приведенные факты соответствуют действительности. Но можно ли сделать из этого вывод, что сложные плодовые тела других афиллофоровых грибов развивались от предков, близких к нынешним рогатиковым? Скорее можно допустить, что рогатиковые грибы - тупик развития одной из линий эволюции примитивных афиллофоровых.
Наиболее распространенный из лесных видов рогатиковых грибов - рамария Инвала (Ramaria invalii, табл. 32). Желто-охряные сильно разветвленные плодовые тела высотой до 5 (8) см растут небольшими группами в тенистых местах на мертвом покрове под сосной или елью, иногда они образуют кривые линии или полные "ведьмины круги".
Зато плодовые тела рамарии желтой (R. flava) крупные, высотой до 30 см и диаметром до 15 см. Они мясистые, серно-желтые или лимонно-желтые, с очень густо расположенными ветвями (рис. 170).
Любимое место для гриба - сосновые боры, где он растет в августе и сентябре. Двух-трех экземпляров рамарии желтой достаточно для вкусного обеда всей семьи.
Кто желает заниматься гербаризацией и определением богатого видами рода рамария, тому следует уже на месте очень точно описать окраску основания, ветвей и их концов. Опытный грибник может по этим признакам уже в при-роде определить вид гриба.
Сложнее узнать многочисленные виды рода клавария (Clavaria, табл. 30), хотя и у них встречаются плодовые тела различных окрасок. Для надежного определения нужно микроскопирование гиф и спор.
Плодовые тела грибов рода клавариадельфус (Clavariadelphus) цилиндрические, булавовидные или трубовидные, охряно-желтые или рыжеватые (табл. 32).
На гниющих листьях и валежных тонких ветках растет в конце лета клавариадельфус ситниковый (С. junceus). Нитевидные бледно- буроватые до 15 см плодовые тела его растут обычно на расстоянии сантиметр или два друг от друга. Это один из тех видов грибов, которые в течение года превращают опавшие листья в гумус.
Клавариадельфус усеченный (С. truncatus, рис. 171) - очень крупный, растущий в одиночку гриб. Плодовые тела его до 15 см высотой и у вершины до 10 см в диаметре. До сих пор этот замечательный вид найден в СССР только в Прибалтике и на Дальнем Востоке.
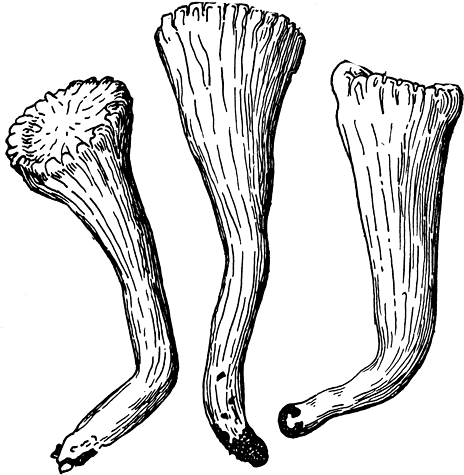
Рис. 171. Клавариадельфус усеченный (Clavariadelphus truncatus). Плодовое тело
Очень своеобразен род мультиклавула (Multiclavula). Их плодовые тела маленькие, только до 3 см высотой и до 2 мм диаметром, булавовидные, простые или немного разветвленные, белые или желтые.
Плодовые тела мультиклавулы весенней (M. vernalis) кремовые, высотой до 2 см, растут большими группами на песчаной земле, покрывая иногда до 4 м2 земли. Земля между плодовыми телами покрыта тонкой пленкой сине-зеленых водорослей или протонемами мхов. Гриб растет в симбиозе с ними, пленка водорослей перепутана рыхлорасположенными гифами мультиклавулы. Мы имеем дело со своеобразным лишайником, мало похожим на остальные.
Некоторые виды тифулы (Typhula) вызывают заболевания культурных растений. В отличие от других рогатиковых грибов у них образуются мелкие бурые или черные склероции, диаметром 1-5 мм, которые погружены в ткань растения-хозяина (субстрата). После смерти растения на склероциях образуются мелкие булавовидные плодовые тела, высотой 2-7 мм и диаметром 1-2 мм.
Тифула инкарнатная (Т. incarnata) вызывает один из разновидностей так называемой снежной плесени хлебных и некоторых других злаковых; тифула клеверная (Т. trifolii) - заболевание клевера и других бобовых.
Семейство Лисичковые (Cantharellaceae)
Лисичковых долго относили к агариковым (пластинчатым) грибам. Действительно, у целого ряда из них плодовые тела шляповидные и трубковидные, со складчатым гименофором. У других же гименофор неровный, желобчатый или почти гладкий. Многие виды желтые, оранжевые или охряные, а другие - буроватые, сероватые.
От настоящих агариковых грибов лисичковые отличаются двумя признаками. Первый из них хорошо заметен невооруженным глазом: вместо тонких пластинок гименофор состоит из вильчатых складок, край которых тупой или даже округленный.
Второй признак цитологический и уловим только сложным препарированием и окрашиванием препарата, но имеет большое значение для понимания эволюции гименомицетов. Как известно, в базидии происходит слияние ядер дикариона, затем следуют два деления, в результате чего образуются четыре ядра, которые идут через стеригмы в образующиеся споры. Деление ядер может произойти на плоскости, перпендикулярной к мысленной оси базидии, но у немногочисленных гименомицетов веретена деления копуляционного ядра находятся продольно. Такой тип базидии называют стихальным, и он характерен именно для семейства лисичковых. Раньше в этом явлении видели доказательство происхождения "стихобазидиальных" гименомицетов от сумчатых грибов: ведь такое же деление ядер происходит в сумке аскомицетов. В настоящее время большинство микологов в этом цитологическом признаке не видят древность лисичковых. Но в то же время это - явное доказательство в пользу отличия лисичковых от всех агариковых, у которых стихобазидий нет.
В семействе только 4 рода и около 100 видов, которые можно найти во всех частях света. В Советском Союзе известно 3 рода и около 10 видов.
Всем известна желтая лисичка (Cantharellus cibarius, рис. 172). Плодовые тела ее можно встретить в хвойных и смешанных лесах от запада до востока СССР, от Карелии до Кавказа (на юге). Плодовые тела яично-желтые, диаметром до 10 см и высотой до 12 см.

Рис. 172. Лисичка желтая (Cantharellus cibarius)
Плодовые тела этого вида растут очень медленно, особенно при сухой погоде. Гриб поселяется в местах с малоразвитым травянистым покровом, быть может, поэтому его можно найти почти только под деревьями или около них. В опытах с искусственным заражением (в лабораторных условиях) установлено, что лисичка образует с сосной микоризу; в природе, по-видимому, может расти и в связи с другими древесными породами.
Лисичка - один из наиболее известных съедобных грибов, которые используют в свежем и консервированном виде (но не в сушеном или соленом). Кроме приятного вкуса, гриб важен большим содержанием некоторых витаминов - B1 (не меньше, чем в дрожжах!), PP. Содержание микроэлементов (цинка, меди) це меньше, чем у агариковых грибов.
Как с культурным шампиньоном, так и с лисичкой проведены успешные опыты по выращиванию мицелия методом глубинного культивирования. Таким путем можно получить ценный пищевой продукт с высокой биологической продуктивностью. Препятствием промышленного производства для нужд человека является только отсутствие специфического вкуса и аромата у "искусственных грибов".
Менее известным для грибника является представитель другого рода - серая лисичка (Craterellus cornucopioides). Плодовые тела его высотой 5-12 см и диаметром 3-8 см воронковидные или трубовидные, с отвернутым, иногда лопастным наружным краем, полые, с очень тонкой пленчатой мякотью, внутри черно-коричневые или почти черные, снаружи покрыты сероватым низкоскладчатым или слегка морщинистым гимением. Растет гриб обыкновенно большими группами летом и осенью в лесах от Прибалтики до Дальнего Востока, а на юге - до Закавказья. Это хороший съедобный гриб, он пригоден как в свежем, так и в сушеном состоянии (табл. 32).
Внешний облик гриба не особенно привлекателен, а после варки он становится совсем черным. Интересны названия этого вида у разных национальностей: немцы зовут его Totentrompete (труба смерти), а англичане - Horn of Plenty (рог изобилия).
Семейство Ежовиковые (Hydnaceae) (Э. Х. Пармасто)
Еще недавно, когда ведущим при систематике гименомицетов считали форму плодовых тел и гименофора, в это семейство включали все грибы, имеющие шиповатый или бородавчатый гименофор. К настоящему времени стало ясно, что этот признак - прекрасный пример конвергенции, который наблюдают у очень многих афиллофоровых грибов, имеющих самое различное происхождение.
В результате углубленных микроскопических исследований к семейству ежовиковых отнесен единственный вид гиднум выемчатый (Hydnum repandum). Его плодовые тела имеют пенек и центральную или эксцентрическую ножку. Шляпка диаметром 3-12 см выпуклая, от белой до светло-охряной, на нижней стороне густо расположенные шипы. Для этого вида, как и для всего семейства, характерны гладкие бесцветные споры, которые не окрашиваются под влиянием иода (неамилоидные).
Плодовые тела растут с лета до осени под деревьями в хвойных и лиственных лесах всего северного полушария. Гриб малоизвестный, но когда шляпки еще молодые, он вкусен и съедобен.
Близко к ежовиковым небольшое семейство герициевых (Hericiaceae). Грибы отличаются амилоидными спорами и наличием своеобразных гиф, имеющих маслянистое содержимое (глеоцистидные гифы). Плодовые тела или в виде прикрепленной боком шляпки, или кустисто-коралловидно-разветвленные, с гименофором в виде висячих шипов.
Во всей лесной зоне у нас летом и осенью растет на валежнике и пнях лиственных пород, особенно березы, гериций коралловый (Hericium coralloides). Его плодовые тела коралловидно-разветвленные, величиной до 40 см, белые или желтоватые, с густо расположенными очень хрупкими шипами, длиной до 1,5 см.
Семейство Телефоровые (Thelephoraceae) (Э. Х. Пармасто)
До начала второй четверти настоящего столетия телефоровыми грибами называли все гименомицеты, имеющие распростертые плодовые тела с гладким или тонкозернистым гименофором. Более углубленное изучение микроскопического строения этой большой группы грибов доказало разнородность телефоровых грибов.
В настоящее время самым важным признаком семейства считают строение и окраску спор. Они могут быть шиповатые или даже с выростами, явно бугорчатые, коричневатые, буроватые или бурые.
Строение плодовых тел может быть самым различным: они распростертые, с гладким или шиповатым гименофором; рогатиковидно-разветвленные; шляповидные, с центральной ножкой и гладким, морщинистым или шиповатым гименофором.
Предпочтение в систематизации этой группы грибов микроскопических признаков казалось многим микологам неприемлемым. Но изучение химического состава плодовых тел подтвердило положение Н. Патуйара, высказанное уже в 1900 г.: у всех изученных представителей семейства, независимо от формы плодового тела, найдено весьма специфическое соединение - телефоровая кислота (один из, терфенилхинонов).
Телефоровые грибы распространены во всем мире в местах, где имеются леса; они обитают на древесине, на опаде, на почве. К 18 их родам принадлежит около 200 видов.
Грибы рода томентелла (Tomentella) растут на гнилой древесине, валежных стволах, разрушенных уже другими грибами пнях хвойных и лиственных деревьев. Обычно встречаются все оттенки ржавого, коричневого, бурого цветов вплоть до черного (белые, желтые, зеленые, серые, розовые). Плодовые тела всегда распростертые, с мучнистой или слегка зернистой поверхностью. Они так малозаметны, что даже в крупных микологических гербариях представлены очень скудно. Зато они очень красивы даже при небольшом увеличении микроскопа: хорошо видны гифы, базидии, своеобразные шиповатые или почти звездчатые споры.
Каково значение такой своеобразной оболочки спор? Можно думать, что споры, снабженные длинными шипами, более медленно падают в воздухе (из-за большего трения) и поэтому переносятся ветром дальше, чем гладкие споры. Опытов по сравнению распространения гладких и шиповатых спор, к сожалению, до сих пор почти не проведено.
В сухих сосновых лесах и особенно на лесосеках часто встречается телефора наземная (Thelephora terrestris, рис. 173). Ее плодовые тела раковинчатые или почти воронковидные, кожистые, темно-коричневые, с бугорчатым сероватым гименофором. Когда плодовое тело образуется рядом с сеянцем сосны (или какого-либо другого дерева), оно может обволакивать сеянцы и приводить их к гибели. При этом нельзя называть гриб паразитом: он пользуется сеянцем только как подпором. Тем не менее в лесных питомниках этот гриб может вызывать серьезные повреждения.
Когда Thelephora terrestris не найдет подходящей опоры, гриб может образовать на нижней стороне валежных веток или даже небольших камней распростертые плодовые тела. В таком виде он ничем не отличается от видов рода томентелла (Tomentella); следовательно, один и тот же гриб может проявляться в различных жизненных формах.
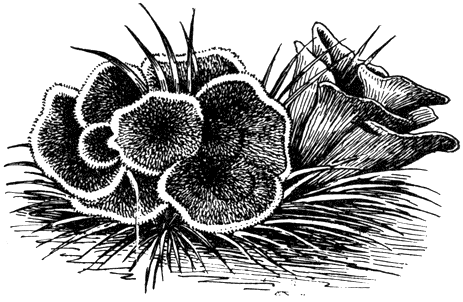
Рис. 173. Телефора наземная (Thelephora terrestris). Плодовые тела
Еще более заметны для наблюдателя природы грибы небольшой группы родов, которых по форме плодового тела иногда называют ежовиковыми грибами. У них шляпка с центральной или эксцентрической, реже боковой ножкой; на нижней стороне шляпки висят густо расположенные шипы.
В роде гиднеллум (Hydnellum) у нас известно 8 видов. Все эти грибы имеют кожистую или деревянистую консистенцию, толстую шляпку и центральную ножку.
В хвойных и смешанных лесах в конце лета и осенью часто встречается гиднеллум плотный (Hydnellum compactum). Обратноконусовидные плодовые его тела (до 12 см в диаметре) растут вередко сросшимися (по 2-3), с войлочной кремовой или коричневатой поверхностью, которая всегда имеет фиолетовый или голубоватый оттенок. Ткань гриба в разрезе с многочисленными голубоватыми полосами. Шипы гименофора бледные, затем ржаво-коричневые.
Не менее часто встречается в хвойных лесах и другой гриб - гиднеллум ржавчинный (Hydnellum ferrugineum, рис. 174), который отличается коричневатыми и коричневыми шляпками без фиолетового оттенка и буроватыми полосами в ткани. Растущие плодовые тела нередко водянистые, содержат жидкость ржавого цвета, которая нередко выступает каплями на поверхности шляпки. Это явление, хорошо известное как гуттация у цветковых растений, наблюдается у целого ряда дереворазрушающих афиллофоровых грибов. "Излишки" воды возникают, по-видимому, в результате интенсивного процесса обмена веществ во время быстрого роста плодовых тел. Известно также, что конечными продуктами разложения древесины и прочих органических соединений у этих грибов служат углекислый газ и вода.
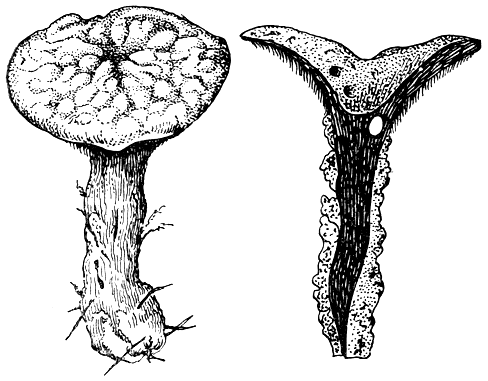
Рис. 174. Гиднеллум ржавчинный (Hydnellum ferrugi- neum). Плодовые тела (справа - в разрезе)
Третий близкий, растущий в таких же местообитаниях вид - гиднеллум оранжевый (Hydnellum aurantiacum) - отличается от описанных меньшими плодовыми телами, до 6 см в диаметре, и оранжевой или слегка коричневатой поверхностью. Шипы гименофора беловатые с оранжевым оттенком, у старых плодовых тел буроватые.
Род саркодон (Sarcodon) имеет мясистые плодовые тела довольно больших размеров, шляпки нередко до 20 см в диаметре. Грибы растут на почве в хвойных и смешанных лесах.
Наиболее распространенный у нас вид - саркодон черепитчатый (Sarcodon imbricatus, рис. 175). Шляпка 5-20 см в диаметре, коричневая, покрытая крупными, более темными, черепитчато расположенными чешуйками. Шипы до 7 мм длиной, светло-коричневые. Ткань горьковатая. Гриб встречается начиная с сентября в хвойных (особенно сосновых) лесах, иногда в большом количестве. Молодые плодовые тела съедобные, но не особенно вкусные.

Рис. 175. Плодовые тела грибов рода саркодон: слева - саркодон черепитчатый (Sarcodon imbricatus); справа - саркодон светло-бурый (S. fuligineo-albus)
Почти такие же плодовые тела, но нередко с пурпуровым оттенком и покрытые только едва заметными чешуйками имеет саркодон лощенный (S. laevigatus). Иногда плодовые тела даже совсем гладкие, растет в сосновых лесах с июля по сентябрь.
Третий близкий вид - саркодон светло-бурый (S. fuligineo-albus, рис. 175) - имеет шляпку несколько меньших размеров (до 10 см в диаметре); поверхность голая, вначале беловатая, затем от кремовой до грязно-охряной. В отличие от большинства видов светло-бурая ткань этого саркодона приобретает при высыхании сильный запах кумарина (напоминает запах сушеных корней цикория). Гриб приурочен к сосновым лесам, где растет с конца лета до начала осени.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'