Порядок Бластокладиевые (Blastocladiales)
Бластокладиевые грибы живут главным образом в пресных водоемах как сапрофиты на погруженных в воду трупах животных или на растительных субстратах, образуя на них пушок до 1 см и более в длину. Некоторые обитают во влажной почве. Немногие паразитируют на беспозвоночных животных или на других низших грибах.
Строение вегетативного тела, или таллома, представителей этой группы грибов варьирует по степени сложности от простого, почти плазмодиального, до хорошо развитого мицелия.
Бесполое размножение осуществляется при помощи зооспор с одним волочащимся сзади жгутиком. Половое размножение представляет собой слияние подвижных гамет или одинаковых по величине и строению (изогамия), или различных (гетерогамия), образующихся в гаметангиях. При этом подвижные клетки (зооспоры, гаметы) бластокладиевых отличаются от зооспор и гамет других грибов тем, что они более активно движутся, а также наличием хорошо заметной ядерной шапочки, состоящей из массы РНК (см. рис. 4).
У подавляющего большинства имеется чередование поколений - спорофита и гаметофита, одинаковых по форме, но иногда различных по величине. На спорофите образуются зооспорангии с диплоидными зооспорами, дающие новые поколения спорофитов. Кроме того, спорофит образует еще и цисты, или покоящиеся споры (по одной в клетке). Они имеют толстую бурую оболочку. При прорастании, сопровождающемся редукционным делением, из нее выходят гаплоидные зооспоры. После периода активного движения они останавливаются, одеваются оболочкой и вырастают в гаметофиты с гаметангиями. Гаметы, выходящие из гаметангиев, похожи на зооспоры, но мельче их. Они, как было сказано выше, или одинаковы, или различаются по величине, у некоторых и по окраске, и степени подвижности. Зигота, образующаяся в результате слияния гамет, дает начало спорофиту.
Самое простое строение характерно для грибов рода целомомицес (Coelomomyces). Все они - облигатные паразиты, поселяющиеся в полости тела личинок москитов. Такой образ жизни наложил определенный отпечаток на строение их вегетативного тела. Несмотря на то что оно достигает достаточного развития, дихотомически или неправильно ветвится, оно не имеет четко выраженной оболочки и ризоидов. По-видимому, таллом воспринимает питательные вещества из клетки, в которой он поселился, всей своей поверхностью. В определенный момент развития весь таллом преобразуется в толстостенные покоящиеся спорангии, которые прорастают, образуя зооспоры (рис. 11). При этом оболочка покоящегося спорангия растрескивается по вполне определенному месту.

Рис. 11. Бластокладиевые: 1 - целомомицес (Coelomomyces): покоящиеся зооспорангии с зооспорами; 2 - катенария алломицис (Gatenaria allomycis): а - покоящиеся споры и ризомицелий в гифе гриба алломицеса (Allomyces), б - прорастающая покоящаяся спора с цистами у отверстия выводной трубки, в, г - выход гамет, д, е - слияние гамет; 3, 4 - бластокладиелла (Blastocladiella), талломы двух видов: а - незрелый спорангий, б - выход зооспор; 5 - бластокладия (Blastocladia): общий вид таллома с зооспорангиями (а) и цистами (б); 6 - алломицес: общий вид таллома с зооспорангиями (а) и цистами (б)
К сожалению, многое в истории развития этих грибов еще неизвестно: в частности, как
происходит заражение личинок москитов, имеется ли у грибов этого рода строгая специализация, есть ли смена поколений и т. д. Однако уже обнаружено много интересных фактов, свидетельствующих об их большой экологической приспособленности. Например, известно, что толстостенные споры целомомицеса, паразитирующего на личинках москитов, яйца которых приспособлены к перенесению периодов засухи, так же способны длительно сохранять жизнеспособность. Для их успешного прорастания даже требуется предварительное высушивание. Покоящиеся спорангии других видов, хозяева которых развиваются только при постоянном наличии влаги, могут совсем не прорасти, если их подвергнуть высушиванию.
Виды рода целомомицес, которых в настоящее время известно уже более 20, интересны не только своей биологией, но и как объект использования их в биологической борьбе, так как некоторые грибы (например, С. punctatus) паразитируют на личинках малярийного комара.
Грибы более морфологически высокоразвитого рода катенария (Catenaria) паразитируют на коловратках и других мелких беспозвоночных животных, на яйцах микроскопических червей, на некоторых грибах или развиваются сапрофитно на растительных или животных остатках. Они характеризуются трубчатым ветвящимся или неветвящимся талломом, уже одетым оболочкой из хитина и имеющим ризоиды, погруженные в субстрат. У них имеется смена поколений. При образовании из таллома зооспорангиев он разделяется перегородками, причем между зооспорангиями, часто располагающимися в ряд, имеются узкие перемычки (рис. 11). Зооспорангии образуют шейки для выхода зооспор. Цисты формируются чаще всего внутри сегментов гиф. Они отличаются утолщенными светло-коричневыми оболочками и при прорастании образуют также шейку, через которую освобождаются зооспоры. Зооспоры или прямо вырастают в новое бесполое растение (спорофит), или инцистируются и превращаются в гаметангий, из которого выходят четыре одинаковые гаметы, сливающиеся попарно. Двужгутиковая зигота вырастает в новый спорофит.
Особенно широко распространен С. anguillulae, живущий в почве и паразитирующий на нематодах и других животных. Он встречается в европейской части СССР, в ряде стран Западной Европы (Франция, Румыния, Великобритания, Ирландия), в Азии, Африке, Южной Америке, Северной Америке (США) и т. д. С. allomycis - паразит на талломах представителей других родов бластокладиевых грибов - бластокладиелла (Blastocladiella) и алломицес (Allomyces); С. sphaerocarpa живет в воде как сапрофит или на талломах некоторых пресноводных водорослей (на водяной сеточке, спирогире, кладофоре и др.), или на элодее, а во влажной почве поселяется на корнях кукурузы, лука и других высших наземных растений.
Еще выше организованные бластокладиевые сапрофиты в воде или в почве на различных растительных и животных остатках.
У некоторых грибов рода бластокладиелла таллом состоит из простого вздутия, от которого отходят многочисленные ризоиды (рис. 11). Это вздутие у спорофита преобразуется затем или в зооспорангий, или в цисту, а у гаметофита образует гаметы. У большинства же видов таллом представлен более или менее удлиненной клеткой с хорошо развитыми ризоидами в основании. На конце ее или образуется один крупный тонкостенный зооспорангий, отделяющийся перегородкой от остальной части таллома, или внутри тонкостенного вместилища формируется циста, одетая толстой двухслойной оболочкой (рис. 11). В данном случае установлено, что появление цист на талломе связано с неблагоприятными для развития условиями. Эти цисты характеризуются большой стойкостью. Они могут переносить высушивание, промораживание, действие высоких температур и т. д.
Зооспоры выходят из зооспорангия через сосочек или поры и после некоторого периода активного движения прорастают в новые растения - спорофиты. Зооспоры, выходящие из цист, отличаются несколько меньшими размерами. Из них вырастают растения внешне такие же, как спорофиты, но образующие гаметы (гаметофиты), причем двудомные. Зигота, образующаяся при слиянии гамет, вырастает в новый спорофит. У Blastocladiella emersonii при повышении содержания СО2 в среде из цист выходят диплоидные зооспоры, образующие снова спорофиты.
Многие виды рода бластокладиелла хорошо развиваются на жидких и твердых питательных средах в условиях лаборатории, поэтому их усиленно используют для изучения самых разнообразных физиологических, цитологических, генетических и других вопросов, особенно В. emersonii. У нее были получены, например, мутанты с пигментированными зооспорами. Наблюдали передачу цитоплазмы от такой зооспоры к нормальной бесцветной (или наоборот) и установили, что таким образом физиологически активный материал может передаваться от одной зооспоры к другой. Это явление, как известно, бывает у некоторых беспозвоночных животных, например у туфельки, и может иметь значение в развитии и росте соответствующего организма.
Представители рода бластокладия (Blastocladia) были описаны раньше других видов бластокладиевых грибов (1878), так как часто встречаются среди других грибов, поселяющихся на растительных остатках, плавающих в воде. На этом субстрате они образуют белые дерновинки длиной 1 мм. Таллом у видов этого рода более расчленен, чем у бластокладиеллы. У большинства видов он состоит из крупной цилиндрической клетки. От нижнего конца ее отходят хорошо развитые ризоиды, а от верхнего - многочисленные дихотомически ветвящиеся гифы, на которых сидят поодиночке или цепочками удлиненные, тонкостенные зооспорангии и округлые толстостенные цисты (рис. 11). Обычно массовое появление цист на талломе связано с повышенным содержанием СО2, что бывает при сильном обрастании субстрата бактериями.
Зооспоры, образующиеся в цистах, прорастают в такие же талломы, которые дают и зооспоры из тонкостенных зооспорангиев, т. е., по-видимому, у видов рода бластокладия нет смены поколений.
Безусловно, самые интересные и высоко дифференцированные среди бластокладиевых грибов - представители рода алломицес (Allomyces, рис. 11). Они обитают в пресных водах на трупах насекомых и других животных, образуя на них дерновинки длиной 1 см. Впервые A. arbusculus был обнаружен и описан в 1911 г. в Индии, затем виды этого рода были найдены в Северной и Южной Америке, Африке, Южной Европе, на Филиппинских островах, на островах Малайского архипелага и т. д. Ведущий и определяющий фактор в распространении видов, по-видимому, температура, так как они почти не встречаются за пределами 40° северной и южной широты.
Большинство представителей рода алломицес имеет смену поколений. При этом спорофит и гаметофит очень сходны по внешнему облику и степени развития. Они состоят из довольно толстых, коротких, ветвящихся гиф, снабженных неглубокими перетяжками и ложными перегородками (там имеются очень широкие просветы - поры). В тех местах, где есть ложные перегородки, на талломе спорофита сидят зооспорангии и толстостенные бурые цисты внутри тонких вместилищ. При этом и зооспорангии и цисты отделяются от таллома настоящими перегородками. Из зооспорангиев выходят диплоидные бесцветные зооспоры, прорастающие затем снова в спорофиты. Весь процесс формирования нового развитого растения занимает 36-48 ч. Зрелые цисты выпадают наружу из разорвавшейся оболочки своего вместилища, наружная оболочка (экзоспорий) их также разрывается, и содержимое, одетое внутренней оболочкой (эндоспорием), высовывается наружу. Из него выходят гаплоидные зооспоры, вырастающие в гаметофиты, внешне очень сходные со спорофитом. На гаметофите образуются, в отличие от других родов, оба типа гаметангиев, обычно один над другим. Мужские гаметангии мельче женских и имеют оранжевую окраску от γ-каротина, а женские бесцветны. Мужские гаметы также окрашены γ-каротином, мельче женских в 2,5-3 раза и более подвижны. Женские гаметы продуцируют половой гормон сиренин, привлекающий мужские гаметы. Мужские и женские гаметы попарно сливаются с образованием зиготы, которая развивается затем в новый спорофит (рис. 12). Такой жизненный цикл установлен для A. javanicus. У некоторых грибов рода алломицес имеются отклонения от описанного цикла развития.
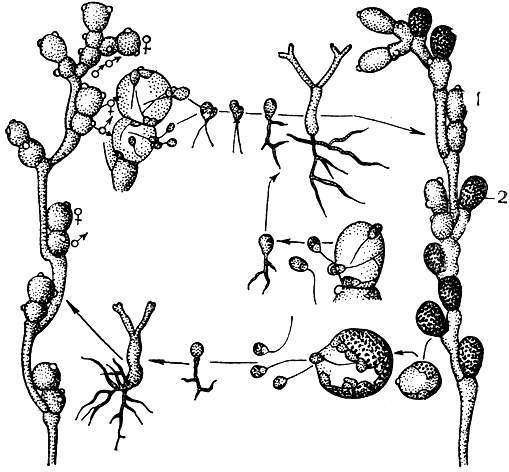
Рис. 12. Бластокладиевые. Алломицес (Allomyces) - схема жизненного цикла: Слева - гаметофит с мужскими (♂) и женскими (♀) гаметангиями; справа - спорофит с зооспорангиями (1) и цистами (2)
Алломицес хорошо растет в культуре на овсяном агаре и некоторых других средах. Поэтому он очень интенсивно изучался и изучается в различных лабораториях мира. Результаты некоторых опытов объясняют многие явления, наблюдающиеся при развитии представителей алломицеса с отклоняющимся циклом. Например, обработка прорастающих зигот колхицином или другими подобными веществами увеличивала частоту прорастания зооспор из покоящихся цист в спорофиты, а не в гаметофиты. Это объясняется тем, что при такой обработке возникает полиплоидность в зооспорах.
При добавлении антибиотика актидиона к среде, на которой выращивался спорофит A. arbusculus, на нем появлялись выросты, несущие гаметангии. Можно было произвольно менять соотношение мужских и женских гаметангиев на гаметофитах, действуя на них веществами, уменьшающими или увеличивающими отношение ДНК и РНК, например тимином. Увеличение количества ДНК вело к появлению больших количеств мужских гаметангиев, и наоборот. Оказалось возможным производить скрещивания некоторых видов алломицеса и получать таким образом гибриды, которые можно было различать друг от друга (в частности, по взаимному расположению мужских и женских гаметангиев и по некоторым другим признакам). Можно было проследить передачу по наследству этих признаков в гаплоидной генерации.
Был установлен такой интересный факт, что A. javanicus, найденный в почве острова Ява, представляет собой природный, естественный гибрид между видами A. arbusculus и A. macrogynes. В 1954 г. он был получен в лаборатории при скрещивании этих видов.
Филогенетически бл астокл а диевые можно рассматривать как результат дальнейшей эволюции хитридиевых грибов, которая шла, во-первых, по пути увеличения размеров центральной части таллома (уже крупного, например, у Macrochytrium botrydiodes). Во-вторых, форма клетки так же изменялась. У бластокладиеллы и бластокладии она стала цилиндрической, а у бластокладия - и ветвящейся, у алломицеса образовался членистый мицелий. В-третьих, осуществлялся переход от изогамии к гетерогамии и выработалась изоморфная смена поколений.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'