Внешнее и внутреннее строение лишайников (Н. С. Голубкова)
Морфология слоевища лишайников
Слоевище лишайников очень разнообразно по окраске, размерам, форме и строению.
Лишайники окрашены в самые различные цвета: белый, розовый, ярко-желтый, оранжевый, оранжево-красный, серый, голубовато-серый, серовато-зеленый, желтовато-зеленый, оливково-коричневый, коричневый, черный и некоторые другие. Окраска слоевища лишайников зависит от наличия пигментов, которые откладываются в оболочках гиф, реже в протоплазме. Наиболее богаты пигментами гифы корового слоя лишайников и различные части их плодовых тел. У лишайников различают пять групп пигментов: зеленые, синие, фиолетовые, красные, коричневые. Механизм образования их до сих пор не выяснен, но совершенно очевидно, что важнейшим фактором, влияющим на этот процесс, является свет.
Иногда цвет слоевища зависит от окраски лишайниковых кислот, которые откладываются в виде кристаллов или зернышек на поверхности гиф. Большинство лишайниковых кислот бесцветны, но некоторые из них окрашены, и иногда очень ярко - в желтый, оранжевый, красный и другие цвета. Окраска кристаллов этих веществ определяет и окраску всего слоевища. И здесь важнейшим фактором, способствующим образованию лишайниковых веществ, является свет. Чем ярче освещение в месте произрастания лишайника, тем ярче он окрашен. Примером может служить широко распространенный, часто встречающийся на стволах осин лишайник ксантория (Xanthoria parietina, табл. 42, 7). Слоевище у него яркое, красновато-оранжевое. Этот цвет придает ему особое лишайниковое вещество - париетин, которое в виде оранжевых кристаллов покрывает гифы корового слоя. Если ксантория растет на солнце, ее слоевище имеет яркую красновато-оранжевую окраску. В условиях слабого освещения этот лишайник полностью теряет оранжевый цвет и становится серовато-зеленоватым. Как правило, очень ярко окрашены лишайники высокогорий и полярных районов Арктики и Антарктики. Это тоже связано с условиями освещения. Для высокогорных и полярных районов земного шара характерны большая прозрачность атмосферы и высокая интенсивность прямой солнечной радиации, обеспечивающие здесь значительную яркость освещения. В таких условиях в наружных слоях слоевищ концентрируется большое количество пигментов и лишайниковых кислот, обусловливая яркую окраску лишайников. Предполагают, что окрашенные наружные слои защищают нижележащие клетки водорослей от чрезмерной интенсивности освещения.

Таблица 42. Накипные (1,2) и листоватые (3-7) лишайники: 1 - Rhizocarpon geographicum, ареолированное слоевище с темным подслоевищем; 2 - Haematomma ventosum, ареолированное слоевище; 3 - Peltigera aphthosa; 4 - Solorina сгосеа; 5 - Umbilicaria muehlenbergii; 6 - Hypogymnia physodes, слоевище сверху и снизу; 7 - Xanthoria parietina
Интересно, что слоевища многих антарктических лишайников окрашены в черный или другой темный цвет. Это тоже объясняется высокой концентрацией зеленых, синих и фиолетовых пигментов в коровом слое и плодовых телах этих лишайников. Хорошо пигментированные наружные слои антарктических лишайников не только защищают клетки водорослей от чрезмерно интенсивного освещения, в данном случае темная окраска слоевища является приспособительной и служит для притягивания тепловых лучей. Как известно, условия в Антарктике крайне суровы; здесь растениям приходится переносить постоянное воздействие очень низкой температуры и жить фактически без воды. Среднегодовая температура воздуха ледяного континента -16 °С. Зимой она падает до -45 °С, а летом, которое наступает в январе - феврале, только днем поднимается выше нуля, ночью же снова опускается до -10 °С.
Из-за низкой температуры осадки выпадают в Антарктике только в виде снега. В такой форме они не могут быть использованы растениями. Вот здесь-то темная окраска лишайников и приходит им на помощь. Темноокрашенные слоевища антарктических лишайников за счет высокой солнечной радиации быстро нагреваются до положительной температуры даже при отрицательной температуре воздуха. Снег, падающий на эти нагретые слоевища, тает, превращаясь в воду, которую лишайник сразу же впитывает. Таким образом он обеспечивает себя водой, необходимой для осуществления процессов дыхания и фотосинтеза.
Насколько разнообразны слоевища лишайников по окраске, настолько же разнообразны они и по форме. Слоевище может иметь вид корочки, листовидной пластинки или кустика. В зависимости от внешнего облика различают три основных морфологических типа: накипные, листоватые и кустистые лишайники.
Слоевище накипных лишайников имеет вид корочки, плотно сросшейся с субстратом. Толщина корочки очень различна. Она может быть весьма тонкой и иметь вид еле заметной накипи или порошкообразного налета; может быть толщиной 1-2 мм, а иногда бывает и довольно толстой, достигая в толщину половины сантиметра. Как правило, накипные слоевища небольших размеров, их диаметр составляет всего несколько?
миллиметров или сантиметров, но иногда может достигать и 20-30 см. В природе нередко можно наблюдать, как небольшие по размерам накипные слоевища лишайников, сливаясь друг с другом, образуют на каменистой поверхности скал или стволах деревьев крупные пятна, достигающие в диаметре нескольких десятков сантиметров (табл. 42, 7, 2).
Как правило, накипные слоевища плотно срастаются с субстратом сердцевинными гифами. Но у некоторых лишайников прикрепление к субстрату происходит с помощью подслоевища. Подслоевище чаще всего бывает темной окраски и обычно образовано темноокрашенными толстостенными грибными гифами. Оно никогда не содержит водорослей (рис. 290). Черную кайму такого подслоевища нередко можно наблюдать по периферии слоевищ некоторых накипных лишайников или же между бугорками ассимиляционного слоевища (табл. 42, 1).
Наиболее примитивный тип накипного слоевища (и вообще слоевища лишайников) - это слоевище в виде тонкого порошкообразного налета. Оно носит название лепрозного. Лепрозные слоевища очень просты по своему строению. Они состоят из скоплений отдельных комочков - клубочков водорослей, окруженных грибными гифами. Такие комочки легко отрываются и переносятся ветром или живот-ными в другие места, где прикрепляются к субстрату и спустя некоторое время разрастаются в новые лепрозные слоевища.
Лепрозные слоевища чаще всего бывают желтоватого или зеленовато-беловатого цвета и нередко покрывают большие поверхности скал или стволов деревьев. Обычно они развиваются во влажных тенистых местах. Их можно встретить на поверхности отвесных скал в узких и темных горных ущельях, в лесах на сырых гниющих пнях, при основании стволов деревьев, на разлагающихся растительных остатках и мхах или слегка увлажненной почве.
Примитивно устроенным, хотя и более сложным по сравнению с лепрозным, считается также накипное слоевище в виде отдельных разбросанных бородавочек или зернышек. Здесь в анатомической структуре уже намечается некоторая дифференциация. Водоросли в такой бородавочке не разбросаны по всей толще и обычно отсутствуют в ее нижней части, а в верхней части бородавочки можно заметить скопление гиф, напоминающее коровой слой.
Более высокоорганизованное накипное слоевище имеет вид сплошной плотной корочки. Такие слоевища обычно уже имеют дифференцированную структуру: на поперечном срезе здесь можно различить коровой слой, слой водорослей и сердцевину.
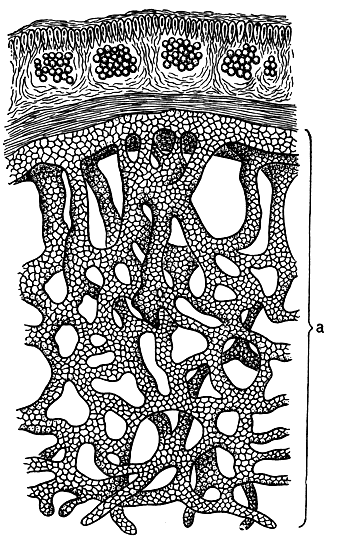
Рис. 290. Поперечный разрез через участок слоевища анции с губчатым подслоевищем (а)
Корочка этих лишайников может быть цельной, гладкой или иметь неровную поверхность - бородавчатую, бугорчатую, с различными шиловидными выростами и т. д. (табл. 42, 1, 2). Нередко слоевище бывает поделено мелкими трещинками на отдельные площадочки, одинаковые по форме и размеру. Эти маленькие площадочки носят название ареол, а сами слоевища называют ареолированными (табл. 42, 2; 43). Лишайники с ареолированной структурой слоевища произрастают только на каменистом субстрате и никогда не встречаются на почве, стволах деревьев, растительных остатках, гниющей древесине и других органических субстратах. Для последних характерно развитие накипных лишайников со слоевищем в виде гладкой, бородавчатой или порошкообразной корочки. Если на них и встречаются трещинки, они обычно бывают неглубокими, неопределенными и никогда не образуют ареол. Особенно характерны ареолированные слоевища для лишайников, произрастающих на поверхности скал в высокогорных районах, пустынях и других областях земного шара с крайними условиями для существования растений. Для таких районов обычны резкие перепады температуры в течение суток, причем на поверхности скал они достигают колоссальных амплитуд - 50-60°. Ареолированная структура слоевища накипных лишайников является приспособлением к перенесению резких колебаний температуры.
Попытаемся проследить за жизнью какого-нибудь наскального пустынного лишайника в течение суток. Каждый день поверхность скалы, на которой растет лишайник, нагревается солнцем до +60, +70 °С и при этом сильно расширяется, а ночью с заходом солнца охлаждается иногда до 0 °С и при этом сильно сжимается. Как же в таких условиях ведет себя лишайник?
На рассвете после холодной ночи на сильно охлажденных за ночь скалах нередко выпадает роса. С первыми лучами солнца наш наскальный лишайник быстро нагревается, гораздо быстрее, чем поверхность скалы, впитывает влагу росы и начинает активно ассимилировать углекислоту, накапливать органические вещества. Такое влажное, разбухшее слоевище заметно увеличивается в размере, в то время как сама поверхность скалы после ночи остается все еще холодной и сильно сжатой. Однако постепенно камень все более и более нагревается и начинает расширяться. Лишайник же с повышением температуры воздуха быстро высыхает, его слоевище резко уменьшается в размере и переходит в характерное для него латентное состояние, когда все процессы в нем замирают. И днем, когда температура скал достигает максимальной величины, на раскаленной и сильно расширенной каменистой поверхности маленький съежившийся лишайник как бы спит. Ночью температура резко падает, поверхность скалы сжимается - гораздо сильнее, чем слоевище самого лишайника. А утром снова на этой сильно сжатой от холода каменистой поверхности происходит расширение слоевища лишайника, увлажненного утренней росой. В результате всех этих изменений, происходящих в течение суток, в слоевище возникают очень сильные напряжения, которые и приводят к появлению на его поверхности многочисленных трещинок. Если бы этого не происходило, резкие изменения в лишайниковом слоевище, противоположные сжатиям и расширениям каменистой поверхности, на которой оно растет, могли бы привести к отрыву слоевища от субстрата. Благодаря ареолированной структуре слоевища эти напряжения ослабляются.
Все перечисленные типы накипных слоевищ являются однообразнонакипными, ибо они одинаковы по своему строению как в центральной, так и в краевой части слоевища. Дальнейшее усложнение в структуре накипных лишайников происходит путем образования переходов к листоватым формам. Особенно часто такие переходы можно наблюдать у ареолированных слоевищ. В этих случаях ареолы, расположенные по периферии лишайника, сильно вытягиваются в радиальном направлении и образуют по краям листовидные лопасти. Такие слоевища имеют вид округлых розеток, в центральной своей части ареольно-потрескавшихся, а по периферии лопастных, и носят название фигурных или радиальных (табл. 44). У высокооргани-зованных зернистых, бородавчатых или гладко-корковых накипных лишайников по периферии слоевища иногда образуется белый или цветной зонированный край. Обычно по окраске он отличается от остального слоевища, так как состоит из радиально растущих гиф микобионта, еще не содержащих водорослей. Позднее водоросли переносятся в этот край из водорослевой зоны двигающими гифами.
Переходной формой между накипными и листоватыми лишайниками является чешуйчатое слоевище, очень характерное, например, для видов, растущих на почве в пустынных областях земного шара (табл. 45; 46, 4, 5). В пустынях на поверхности почвы обычно можно заметить коричневатые, серые, желтоватые и розоватые пятна, образуемые слоевищами чешуйчатых лишайников. Диаметр чешуек колеблется от 2-5 мм до 1 см. Они бывают округлыми, угловатыми, с ровными и волнистыми, иногда лопастными краями. Чешуйки могут быть расположены на некотором расстоянии друг от друга или расти так тесно, что края одной накладываются на поверхность другой. В отличие от типичных накипных слоевищ чешуйки обычно менее плотно срастаются с субстратом, и их легко можно от него отделить. Чаще они прикрепляются отдельными тонкими гифами, отходящими от нижней поверхности. Нередко эти гифы отходят только от какого-нибудь одного края чешуйки, в то время как другой остается свободным. В таких случаях чешуйки приподнимаются и растут не горизонтально, а вертикально. Но иногда они прикрепляются к субстрату только в своей центральной части довольно толстыми тяжами, образованными склеенными сердцевинными гифами. Эти тяжи у лишайников, растущих на почве, могут достигать в длину 0,5-1 см и напоминают маленький разветвленный корешок (табл. 45).
В зависимости от субстрата, на котором произрастают накипные лишайники, среди них различают несколько экологических групп: эпилитные, развивающиеся на поверхности горных пород; эпифлеодные - на коре деревьев и кустарников; эпигейные - на поверхности почвы; эпиксильные - на обнаженной гниющей древесине.
У подавляющего большинства накипных лишайников слоевище развивается на поверхности субстрата. Однако существует еще одна сравнительно небольшая, но интересная группа лишайников, слоевище которых целиком растет внутри камня или коры дерева. Если такое слоевище развивается внутри камня, его называют эндолитным; если внутри коры дерева - эндофлеодным или гипофлеодным. Эти лишайники можно разделить на две группы. У представителей одной из них слоевище полностью погружено в субстрат и никогда не выступает на его поверхность, изредка выступают лишь плодовые тела лишай-ника; у лишайников второй группы слоевище на поверхности субстрата развивает коровой слой и зону водорослей, а в субстрате - сердцевину и зону с прикрепляющими гифами.
Эндолитные лишайники чаще всего развиваются внутри известковых пород, но могут встречаться и внутри силикатных скал, хотя в таком случае видов, полностью погруженных в субстрат, известно немного. Слоевищные гифы эндолитных лишайников способны проникать внутрь камня на значительную глубину, от 1 до 3 см. Чаще всего гифы и водоросли лишайника при своем продвижении в глубь камня используют мелкие трещинки, щели, но они обладают способностью проникать и внутрь горных пород, совершенно не тронутых разру-шением. Оказывается, гифы эндолитных лишайников выделяют кислоты, растворяющие горные скалы. Благодаря этому они могут разрушить даже такие твердые породы, как гранит, который в этих случаях довольно быстро превращается в тонкозернистую массу, напоминающую глину.
Гифы эндолитных лишайников, проникающие в субстрат, обычно очень тонкие (толщина их всего 1-3 мкм), нежные, с длинными клетками. Часто они не растут прямо, а сгибаются на концах в сторону в виде крючков, охватывающих кусочки субстрата. Иногда на конце этих гиф образуются клетки-щетинки - длинные, тонко заостренные на конце волосковидные клетки (рис. 291).
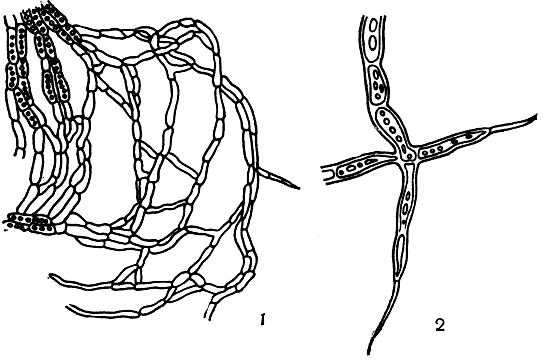
Рис. 291. Гифы эндолитных лишайников: 1 - дуговидно-согнутые гифы, проникающие в толщу камня; 2 - конечные клетки-щетинки этих гиф
Проникая в горную породу, гифы обходят твердые, плохо растворимые минералы и быстро распространяются по более рыхлым и легче растворимым участкам. Так, например, они довольно быстро разрушают слоистые кристаллы слюды. Гифы разъединяют листочки слюды и проникают между ними. Здесь они разветвляются и отодвигают пластинки слюды одну от другой. Постепенно разрастаясь и разветвляясь, гифы образуют между пластиночками грибную плектенхиму. Затем в эту плектенхиму проникают и клетки водорослей, которые размножаются, обвиваются гифами и все больше раздвигают отдельные листочки слюды. Было замечено, что на многих твердых горных породах гифы эндолитных лишайников проникают внутрь камня именно в участках, занятых пластинками слюды, а дальше продвигаются уже благодаря химическому разрушению породы.
Разрушая твердые горные породы, превращая их в зернистую массу, эндолитные лишайники тем самым выступают как одни из пионеров растительности. Они подготавливают поверхность скал для поселения других растений: листоватых и кустистых лишайников, мхов, цветковых растений и т. д. Но вместе с тем эти лишайники играют в жизни человека и отрицательную роль. Особый ущерб они наносят памятникам старины, нередко поселяясь на них и разрушая их. Например, широко известно, какой вред причинили лишайники старым витражам церквей в Западной Европе.
Эндофлеодные лишайники чаще всего поселяются на древесных породах с тонкой или гладкой корой. Слоевище их обычно имеет вид овальных пятен. Длинная ось такого овального пятна обычно размещена горизонтально. Предполагали, что подобная форма слоевища объясняется ростом ствола в толщину. Но оказалось, что она зависит от формы клеток коры дерева. Если они сильно вытянуты в горизонтальном направлении, то и слоевище вытянуто горизонтально. Если клетки коры одинаковы по длине и ширине, то и слоевище эндофлеодных лишайников приобретает округлую форму.
Слоевище этих лишайников обычно постепенно проникает в кору дерева. Первыми по маленьким трещинкам, которые возникают в результате роста дерева в толщину, проникают вглубь гифы микобионта. А через некоторое время туда же проталкиваются и клетки водорослей, которые из округлых временно становятся удлиненными. С появлением водоросли начинается быстрый рост лишайника в ширину и дальнейшее проникновение эндофлеодного слоевища в более глубокие слои коры. Через некоторое время развиваются и плодовые тела, которые у всех эндофлеодных лишайников расположе-ны на поверхности коры дерева.
Обычно гифы эндофлеодных лишайников растут между мертвыми клетками коры, расщепляя их на небольшие участки. Способны ли гифы пробивать оболочку клеток коры дерева, пока еще неизвестно. Однако вряд ли можно допустить, чтобы гифы, проникая внутрь только по трещинкам в коре, могли образовывать столь оформленные слоевища. Трудно также объяснить разделение пробки коры на небольшие участки только механическим воздействием. Скорее всего гифы лишайника оказывают на коровые клетки дерева и химическое воздействие. К этому выводу приводят некоторые наблюдения. Например, в местах контакта гиф лишайника с клетками коры были обнаружены повреждения оболочек клеток пробки, а в некоторых случаях и вообще деформированные одревеснелые оболочки. Кроме того, в этих оболочках очень часто отсутствовала окраска, характерная для лигнина. Поэтому ученые допускают мысль, что гифы лишайников, растущих на коре деревьев и кустарников, обладают целлюлозолитической способностью и содержат ферменты, расщепляющие клетчатку.
Иногда типично эндофлеодные лишайники долго остаются полностью погруженными в субстрат, но с изменением условий освещения становятся поверхностными. Большей частью эти изменения зависят от характера коры. Так, эндофлеодные слоевища лишайников, развивающиеся в тонкой коре ясеня, в условиях хорошего освещения начинают выступать из более глубоких слоев и становятся почти полностью поверхностными. Значит, один и тот же лишайник может быть эндо- и эпифлеодным.
Еще одна чрезвычайно интересная группа накипных лишайников - это лишайники с шаровидной формой слоевища. Они широко известны под названием кочующих лишайников. Встречаются кочующие лишайники в засушливых областях земного шара, в равнинных и горных степях, пустынях и иногда в предгорных районах. Слоевище у них комковато-шаровидной формы (табл. 50) и не прикреплено к субстрату. Такие комочки свободно лежат на поверхности почвы, и ветер или животные переносят их с места на место как маленькие перекати-поле. Форма комочков может быть самой разнообразной - от округлой до угловатой, лепешковидной и неправильной. Их поверхность бывает складчатой, бородавчатой, чешуйчатой или покрыта сосочковидными выростами. Кочующий образ жизни в засушливых условиях привел к развитию у этих лишайников толстого и плотного корового слоя. Но на поверхности этого слоя можно заметить небольшие беловатые углубления, называемые псевдоцифеллами. Это органы газообмена - разрывы коры, через которые воздух проникает внутрь слоевища. Обычно в сердцевинном слое этих лишайников между гифами скапливаются кристаллы окисла кальция.
Эти лишайники, главным образом представителей рода аспицилия (Aspicilia), иногда называют также "лишайниковой манной". Когда- то в пустынных областях в голодные годы их добавляли в пищу. В наше время алжирские крестьяне нередко используют эти лишайники как корм для овец.
Слоевище листоватых лишайников имеет вид листовидной пластинки, горизонтально распростертой на субстрате. Наиболее характерна для него округлая форма, которая обусловлена горизонтально-радиальным ростом гиф. В начале образования слоевища гифы листоватых лишайников растут от одного зачатка по радиусам окружности. У взрослых растений также наблюдается краевой рост гиф. Обычно молодые слоевища имеют правильную округлую форму, но позднее они начинают неравномерно разрастаться и форма их делается неправильной. Обычно форма слоевища определяется характером субстрата. Чем ровнее его поверхность, тем более правильную округлую форму имеют слоевища листоватых лишайников.
Наиболее простое слоевище листоватых лишайников имеет вид одной крупной округлой листовидной пластинки, достигающей в диаметре 10-20 см. Такая пластинка нередко бывает плотной, кожистой, окрашенной в темно-серый, темно-коричневый или черный цвет (рис. 292). Слоевище, состоящее из одной листовидной пластинки, носит название монофильного. Монофильное пластинчатое слоевище обычно прикрепляется к субстрату только в своей центральной части с помощью толстой короткой ножки, называемой гомфом (рис. 292, 2а). Такой формой слоевища обладают представители рода умбиликария (Umbilicaria) и некоторые виды рода дерматокарпон (Dermatocarpon). Это ксерофитные формы, широко встречающиеся на скалах в Арктике, Антарктике и высокогорьях. Толстые плотные кожистые слоевища этих лишайников, надежно прикрепленные к поверхности субстрата толстой короткой ножкой, легко переносят действие сильных ветров, снежные бури, ураганы и могут существовать длительные периоды без воды. Иногда такие слоевища состоят не из одной, а из нескольких листовидных пластинок, и тогда их называют полифильными (табл. 42, 3,4,5).
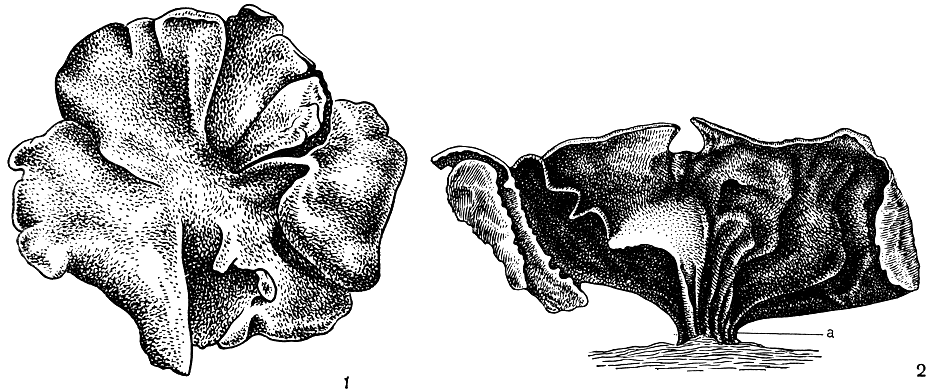
Рис. 292. Монофильное пластинчатое слоевище: 1 - вид слоевища сверху; 2 - вид слоевища сбоку, а - гомф
Нередко пластинчатое слоевище бывает по краям выемчато вырезано или рассечено на широкие доли. Примером могут служить крупнолистоватые слоевища видов рода пельтигера (Peltigera), которые нередко встречаются в лесах на почве среди мха или на основаниях стволов деревьев, поросших мхом (табл. 42, 3). Во влажных старых тенистых лесах на стволах деревьев или мшистых скалах растут причудливо вырезанные крупнолистоватые слоевища лобарий (Lobaria) и стикт (Sticta, табл. 47, 1,4). Эти лишайники обычно довольно рыхло прикрепляются к субстрату всей своей нижней поверхностью, свободными остаются лишь приподнимающиеся кверху края.

Таблица 47. Листоватые лишайники: 1 - Lobaria pulmonaria; 2 - Cetraria richardsonii; 3 - Parmelia caperata; 4 - Sticta wrightii; 5 - Cetraria pinastri; 6 - Leptogium saturninum
Более сложным по строению является листоватое слоевище, рассеченное на множество мелких лопастей. Эти лопасти бывают самого разного размера и формы: узкими и широкими, слабо- и сильноветвистыми, плоскими и выпуклыми, тесно сомкнутыми и разделенными, налегающими друг на друга своими краями или строго отграниченными (табл. 47). Как правило, они собраны в округлые розетки, но иногда образуют слоевища неопределенных, бесконечно разнообразных форм - по внешнему облику они напоминают искусно сплетенные кружева, окутывающие разноцветным чехлом стволы и ветви деревьев (табл. 42, 6).
Характерной особенностью листоватого слоевища лишайников является его дорсовентральное строение, при котором верхняя поверхность отличается по строению и окраске от нижней. Например, у представителей рода пельтигера (Peltigera) верхняя сторона слоевища сероватая или серовато-коричневатая, гладкая или слегка шероховатая, а нижняя - беловатая, розоватая, в центре нередко черно-серая, войлочная, с многочисленными жилками и лохматыми тяжами прикрепительных гиф. У видов рода гипогимния (Hypogymnia) верхняя сторона гладкая, серая, а нижняя - черная. У лишайника солорина скортея (Solorina scortea) слоевище сверху окрашено в серовато-коричневый цвет, а снизу - в яркий, красновато-оранжевый (табл. 42, 4).
Верхняя поверхность слоевища листоватых лишайников бывает ровной, волнистой, ямчатой, голой, глянцевитой или матовой, нередко шероховатой, неровной, покрытой бугорками, бородавочками. Иногда на ней имеются различной формы выросты, реснички, образующие опушение или войлочный налет.
Нижняя поверхность также разнообразна по своему строению, но ее характерной особенностью является то, что она почти всегда образует особые органы, с помощью которых листоватый лишайник прикрепляется к субстрату. В отличие от накипных лишайников, слоевище которых целиком плотно срастается с субстратом, листоватые лишайники обычно довольно рыхло с ним связаны и в большинстве случаев могут быть легко от него отделены. Только очень немногие виды, например представители рода гипогимния, прикрепляются к субстрату прямо нижним коровым слоем. Но и в таком случае не происходит плотного срастания субстрата и всей нижней поверхности лишайника - слоевище прикрепляется отдельными участками нижней поверхности. У огромного большинства листоватых лишайников на нижней стороне слоевища образуются особые органы прикрепления - ризоиды, ризины или гомф. Они образуются тяжами гиф и отличаются друг от друга анатомическим строением. У видов рода анция (Anzia), распространенных в тропических и субтропических областях Азии, Америки, Австралии и Океании, а в СССР встречающихся на Дальнем Востоке, образуется губчатое подслоевище, с помощью которого эти лишайники прикрепляются к субстрату (рис. 290, а).
Листоватые лишайники по сравнению с накипными являются значительно более высокоорганизованными формами. В эволюционном отношении оказалось выгодным отделение слоевища от субстрата. Между ними появился небольшой промежуток, и это дало целый ряд преимуществ. Во-первых, в нем заключена прослойка воздуха, способствующая лучшему газообмену внутренних слоев слоевища. Во-вторых, там дольше задерживается влага, благодаря чему слоевище более длительное время может находиться во влажном состоянии. В-третьих, в узком пространстве между поверхностью субстрата и слоевищем обычно задерживаются различные органические и неорганические вещества, которые могут быть использованы растением.
Но, с другой стороны, отделившись от субстрата, лишайник обрек себя на массу неудобств. Он стал более уязвим по отношению к факторам внешней среды - действию ветра, ударам дождя и снега, нападению животных и т. д. И прежде всего оказалась незащищенной нижняя поверхность лишайника.
Прогрессивная роль отделения лишайника от поверхности, на которой он рос, состояла в усложнении анатомической структуры слоевища и развитии особых органов прикрепления. Прежде всего на слоевище появился нижний коровой слой, обычно отсутствующий у накипных форм лишайников. Огромное большинство листоватых лишайников имеет на нижней стороне слоевища хорошо развитый коровой слой. Исключение составляют крупнолистоватые слоевища пельтигеры. Нижняя поверхность этих лишайников выстлана рыхлорасположенными сердцевинными гифами, которые образуют также длинные пучки ризоидальных тяжей, проникающих в субстрат. Эти лишайники обычно растут на почве, среди мхов. Своеобразное строение нижней поверхности, отсутствие нижнего корового слоя позволяют этим растениям лучше и быстрее втягивать влагу из влажных дерновинок мхов.
Однако отделение слоевища от субстрата привело не только к образованию корового слоя на нижней стороне лишайников, но и к усложнению анатомической структуры всего слоевища. В отличие от накипных лишайников у листоватых форм в слоевищах существует четкая дифференциация анатомических слоев. Как правило, под микроскопом на поперечных срезах таких слоевищ можно различить 4 хорошо дифференцированных слоя: верхний коровой слой, слой водорослей, сердцевину и нижний коровой слой. Особенно большого разнообразия у листоватых лишайников достигает структура коровых слоев, которые выполняют здесь не только защитную, но и укрепляющую роль.
Среди листоватых лишайников также встречаются неприкрепленные, кочующие формы. В степях юга и юго-востока нашей страны и в Монголии на поверхности почвы обычно можно встретить свободнолежащие зеленовато-черные слоевища пармелии блуждающей (Parmelia vagans, табл. 50). Они не прикреплены к субстрату и свободно переносятся ветром с места на место. В горных тундрах Сибири и Чукотки на сухих горных склонах и пологих сопках обитает другой очень красивый кочующий лишайник цетрария Ричардсона (Cetraria richardsonii, табл. 47, 2). Темно-коричневые слоевища его в сухом состоянии сжимаются и свертываются в комки, которые, как миниатюрные перекати- поле, кочуют с помощью ветра с места на место.
Слоевище кустистых лишайников имеет вид прямостоячего или повисающего кустика, реже неразветвленных прямостоячих выростов.
По организационному уровню кустистые лишайники представляют высший этап развития слоевища.
В отличие от накипных и листоватых форм лишайников, для которых характерен горизонтальный рост гиф, у кустистых лишайников наблюдается вертикально направленный рост гиф и верхушечный рост слоевищ. Это позволяет кустистым лишайникам путем изгибов веточек в разные стороны занимать наилучшее положение, при котором водоросли могут максимально использовать свет для осуществления фотосинтеза. Эти лишайники обычно прикрепляются к субстрату только небольшим участком нижней части слоевища. Прямостоячие напочвенные кустистые лишайники чаще всего прикрепляются к почве тонкими нитевидными ризоидами. Прикрепление повисающих слоевищ кустистых лишайников к коре дерева или поверхности скал происходит с помощью псевдогомфа. Последний имеет вид короткой ножки, расширенной на конце в небольшую пяточку (рис. 293); он напоминает по внешнему виду гомф листоватых лишайников, но отличается от него анатомическим строением. Редкое исключение среди кустистых лишайников составляет антарктический лишайник гимантормия траурная (Himantormia ligubris). Слоевище его, имеющее вид черных, как бы мертвых кустиков с разбросанными белыми пятнами, прикрепляется к скалам с помощью подслоевища (табл. 49, 4).
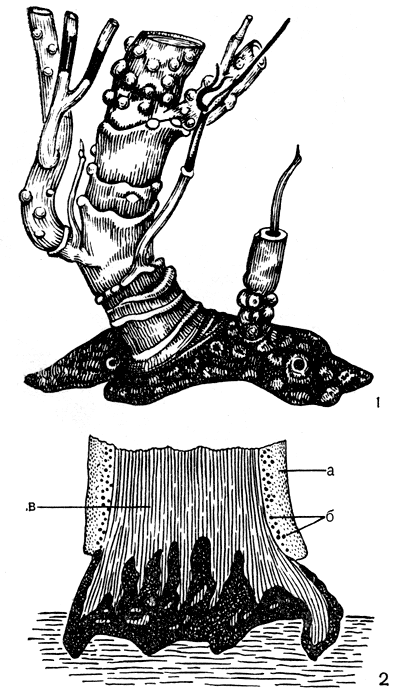
Рис. 293. Псевдогомф: 1 - общий вид псевдогомфа у нейропогона (Neuropogon); 2 - анатомическое строение псевдогомфа: а - коровой слой, б - клетки водорослей, в - сердцевинные гифы
Слоевища кустистых лишайников могут быть разных размеров. Высота самых маленьких составляет всего несколько миллиметров, а наиболее крупных 30-50 см. Повисающие слоевища кустистых лишайников иногда могут достигать колоссальных размеров. Так, длина одного из эпифитных лишайников - уснеи длинной (Usnea longissima), свисающей в виде бороды с ветвей лиственниц и кедров в таежных лесах, составляет 7-8 м (табл. 49, 8).
Слоевища кустистых лишайников чрезвычайно разнообразны по форме. Наиболее простые имеют вид отдельных прямостоячих неразветвленных выростов. Ярким примером может служить слоевище тамнолии (Thamnolia, табл. 48, 3). В тундрах Крайнего Севера или высокогорного пояса на почве среди дерновинок мхов и лишайников нередко можно встретить в беспорядке разбросанные молочно-белые или бледно-розоватые заостренные на конце палочки. Эти простые, вертикально растущие палочки, напоминающие по виду маленькие пики, представляют собой слоевище арктоальпийского лишайника тамнолии червеобразной (Thamnolia vermicularis, табл. 51). Но чаще кустистые лишайники бывают разветвленными и образуют слоевище в виде густых компактных дерновинок (табл. 48; 49, 2, 2, 4, б, 7). Такой формой слоевища обладают очень многие лесные и тундровые напочвенные лишайники. В сухих сосновых борах, в северных и высокогорных тундрах нередко можно наблюдать на поверхности почвы сплошные разноцветные ковры, образованные дерновинками кустистых лишайников. Кустистые слоевища эпифитных лишайников обычно имеют вид лохматого кустика. Но иногда их лопасти бывают очень тонкими, сильно вытянутыми, почти нитевидными. Такие слоевища напоминают по внешнему облику длинные седые или черные бороды (табл. 49, 8).

Таблица 48. Напочвенные кустистые лишайники тундр и сосновых лесов: 1 - Cetraria cucullata; 2 - Cladonia floerkeana; 3 - Thamnolia vermicularis; 4 - Cladonia deformis; 5 - Cetraria islandica; 6 - Cladonia alpestris
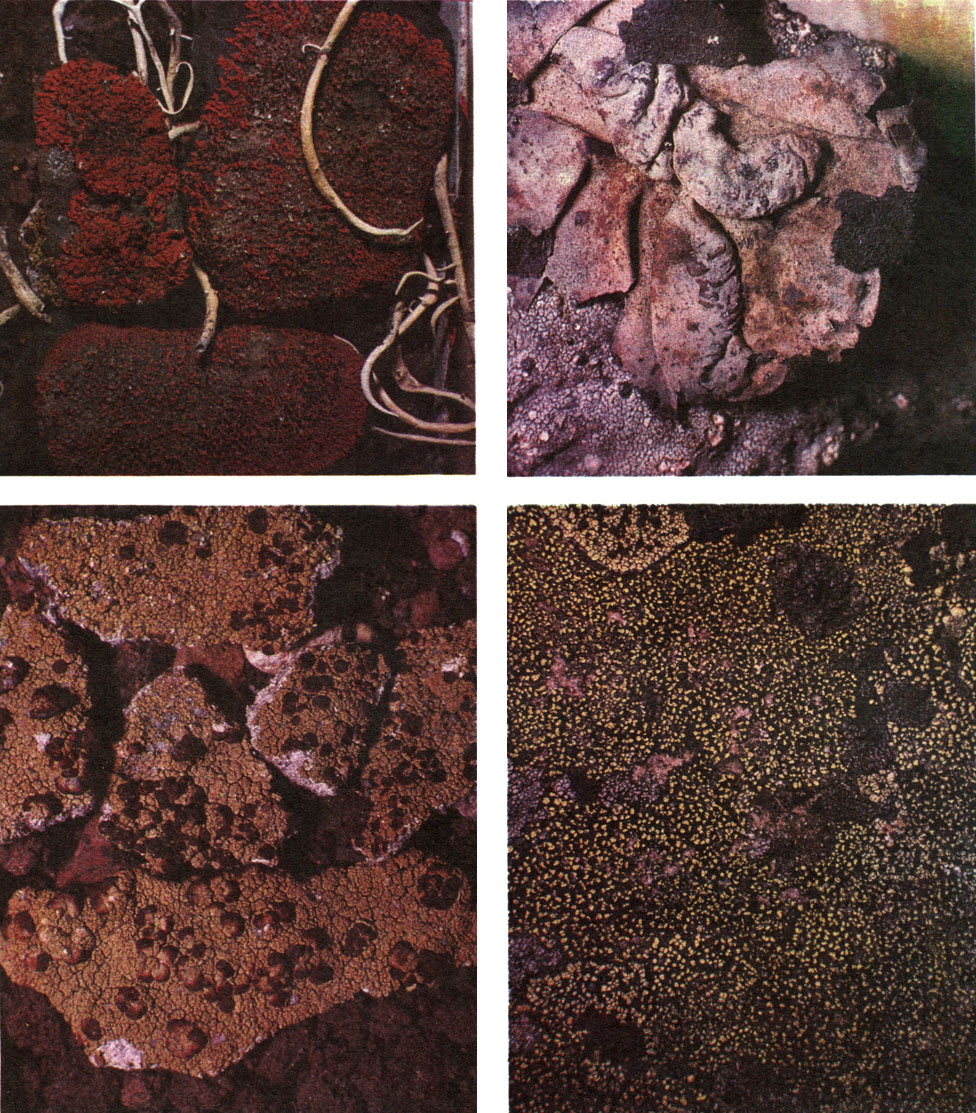
Таблица 51. Арктоальпийские лишайники: вверху слева - Xanthoria elegans и Thamnolia vermicularis; вверху справа - Umbilicaria vellea на скалах; внизу слева - Haematomma ventosum; внизу справа - Rhizocarpon tinei
Среди кустистых лишайников различают слоевища с плоскими и округлыми лопастями. Более примитивным типом строения является слоевище с плоскими лопастями. Эти слоевища ближе всего стоят к листоватым лишайникам, среди них имеются многочисленные переход-ные формы между листоватыми и кустистыми слоевищами. Как и у листоватых лишайников, у кустистых слоевищ с плосколинейными лопастями нередко наблюдается дорсовентральное строение. Примером может служить широко известная цетрария исландская - характерный представитель напочвенных лишайников сосновых лесов, болот и северных тундр (табл. 48, 5). Слоевище этого кустистого лишайника имеет вид рыхлой дернинки высотой до 10 см и образовано плоскими, местами желобчато свернутыми лопастями. Верхняя поверхность этих лопастей коричневая или зеленовато-коричневая, блестящая. Нижняя поверхность окрашена светлее, она светло-коричневая, почти до белой, с многочисленными белыми пятнышками, разрывами коры, которые служат для проникновения воздуха. Но, в отличие от листоватых лишайников, на поперечном срезе этих лопастей можно различить уже не четыре, а пять анатомических слоев. Помимо верхнего и нижнего коровых слоев, слоя водорослей и сердцевины, у этого лишайника имеется еще один слой водорослей, расположенный над нижней корой слоевища. Таким образом, уже у кустистых лишайников с плосколинейными лопастями благодаря вертикальному росту слоевища происходит увеличение поверхности, занятой водорослями, что способствует более интенсивному процессу фотосинтеза.
Далее в эволюционном развитии кустистых лишайников намечается переход к радиальной структуре слоевища, как наиболее выгодной для лишайника. В цилиндрических лопастях водоросли располагаются по окружности и тем самым достигаются равномерность их освещения со всех сторон и максимальное увеличение ассимиляционной поверхности слоевища. Переход от плосколинейной структуры лопастей к радиальной происходит прежде всего через свертывание плоских лопастей в трубочки. Такого типа лопасти можно наблюдать у напоч-венного кустистого лишайника цетрарии клубочковой (Cetraria cucullata), характерного обитателя северных и горных тундр. Ярко-желтое или бледно-зеленовато-желтое слоевище его образовано желобчато свернутыми, почти трубчатыми лопастями (табл. 48, 1). У более продвинутых в эволюционном отношении кустистых лишайников лопасти слоевища становятся угловато-округлыми и теряют свое дорсоветральное строение. Верхняя и нижняя поверхности этих лопастей одинаково окрашены и аналогичны по своему строению. Но, несмотря на радиальную анатомическую структуру, по форме эти лопасти не цилиндрические, а слегка сдавленные, почти до плоских и лишь в некоторых участках округлые. Лопасти такой формы имеет, например, широко распространенный эпифитный лишайник летария (Letharia thamnodes), напочвенные лишайники рода алектория (Alectoria ochroleuca) и некоторые другие (табл. 49).
Своеобразной переходной формой между листоватыми и кустистыми лишайниками являются слоевища видов рода кладония (Cladonia) - широко распространенных напочвенных кустистых лишайников. По своему строению эти слоевища считаются уже радиально-кустистыми. У большинства представителей рода кладония тело образовано двумя типами слоевищ: горизонтальным - чешуйчатым, реже бородавчатым и вертикальным - различной формы выростами или кустиками, растущими вверх от чешуек (табл. 48,2, 4, 6). При образовании слоевища у кладонии сначала вырастают горизонтальные чешуйки. Эти чешуйки, достигающие обычно в диаметре от 2 до 5 мм, но иногда имеющие вид небольших листиков диаметром до 10-20 мм, сверху зеленоватые, снизу беловатые или желтоватые, собраны в небольшие дерновинки. По анатомическому строению они обычно трехслойные. В них можно различить верхний коровой слой, слой водорослей и сердцевину. Спустя некоторое время на поверхности или по краям этих чешуек появляются вертикально направленные выросты, которые называют подециями. Форма подециев чрезвычайно разнообразна. Они могут быть простыми, неразветвленными, иметь вид шиловидных или роговидных выростов (рис. 294,1, 2). Часто они на конце расширены в виде бокалов (рис. 294, 3). Такие кубковидные подеции называют сцифами . Иногда из центра такого бокальчика вырастает еще один, а из него, в свою очередь, следующий, и таким образом развиваются как бы многоэтажные подеции, образованные сцифами, растущими один из другого (рис. 294, 4). Нередко подеции бывают сильно разветвленными и имеют вид кустика (рис. 294, 5). Такая форма подециев, например, очень характерна для группы кладоний, которую называют ягелем или оленьим мхом. Этими лишайниками на Севере питаются олени. Общее количество лишайников, поедаемых оленями, составляет не менее 50 видов. Но особенно олени предпочитают кладонии. Среди них наиболее "любимыми" являются кладония приальпийская (Cladonia alpestris), кладония оленья (С. rangiferina), кладония лесная (С. sylvatica) и кладония мягкая (С. mitis). У этих видов, в отличие от других кладоний, горизонтальное слоевище не чешуйчатое, а бо-родавчатое и рано исчезает. И слоевище этих лишайников фактически состоит лишь из одних подециев, сильно разветвленных и образующих довольно крупные дернинки.
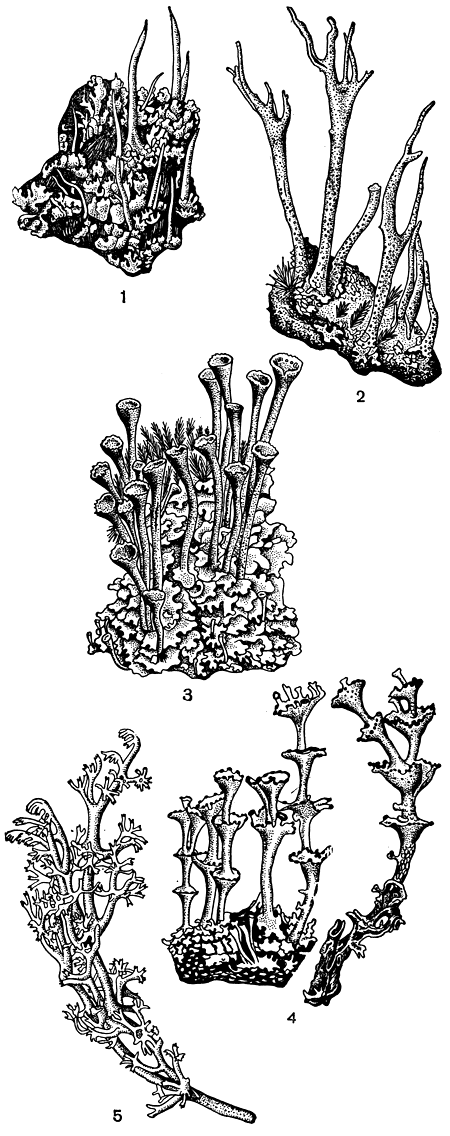
Рис. 294. Различные типы подециев у лишайников из рода кладоний: 1 - шиловидные подеции Cladonia coniocraea; 2 - роговидные подеции Cladonia subulata; 3 - кубковидные подеции (сцифы) Cladonia fimbriata; 4 - пролифицирующие сцифы Cladonia verticillata; 5 - разветвленные педеции Cladonia sylvatica
Подеции кладоний округлые в сечении и имеют радиальную анатомическую структуру. Снаружи в них можно различить коровой слой, далее слой водорослей, расположенный по окружности, а внутри подеция - сердцевину. Но нередко эти анатомические слои бывают нечетко выражены или вовсе не развиты. Так, например, у группы ягелей на поверхности подециев никогда не развивается коровой слой, их поверхность образована рыхлорасположенными сердцевинными гифами (рис. 295). У многих видов кладоний коровой слой развит на поверхности подециев в виде отдельных островков. Слой водорослей также не является непрерывным, а состоит из отдельных разбросанных группок водорослей. Сердцевина подециев, примыкающая к зоне водорослей, обычно образована рыхлорасположенными гифами, а далее по направлению к центру имеет аморфное строение и состоит из плотно склеенных гиф. У большинства видов кладоний в центре подециев образуется внутренняя полость, не заполненная сердцевинной тканью.
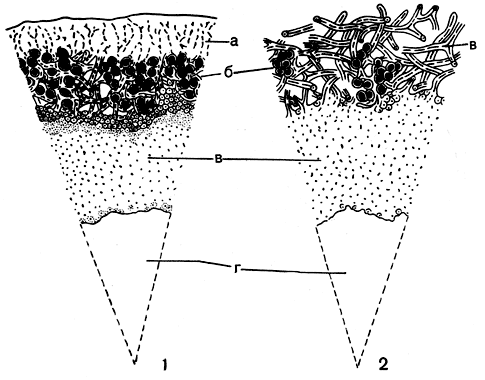
Рис. 295. Секторы поперечных разрезов через подеции кладоний: 1 - Cladonia furcata; 2 - Cladonia rangiferina. a - коровой слой, б - слой водорослей, в - сердцевина, г - центральная полость
Из лишайников, лопасти которых имеют типично радиальную структуру, можно назвать виды рода уснея (Usnea) и многие виды рода алектория (Alectoria). Лопасти этих лишайников, как правило, округлые, тонкие и сильно вытянутые. Чаще всего они образуют длинные, повисающие слоевища в виде редких бледно-зеленых, серых и буро-черных "бород" (табл. 49, 7, 8). На поперечном разрезе округлых лопастей этих лишайников можно различить снаружи плотный, хорошо развитый коровой слой, под ним расположен слой водорослей, а центральная часть занята сердцевиной. Причем у алекторий сердцевина однородная, она образована беспорядочно рыхло расположенными гифами. Но у усней происходит дальнейшее усложнение анатомической структуры слоевища. Сердцевина этих лишайников неоднородна, в ней можно различить периферическую часть, состоящую из рыхлорасположенных гиф, и центральную часть, образованную продольно идущими, плотно склеенными гифами. Эти склеенные между собой толстостенные гифы, расположенные параллельно оси лопастей, образуют так называемый центральный, или осевой, тяж. Осевой тяж придает прочность тонким и сильно вытянутым лопастям этих лишайников (рис. 296).
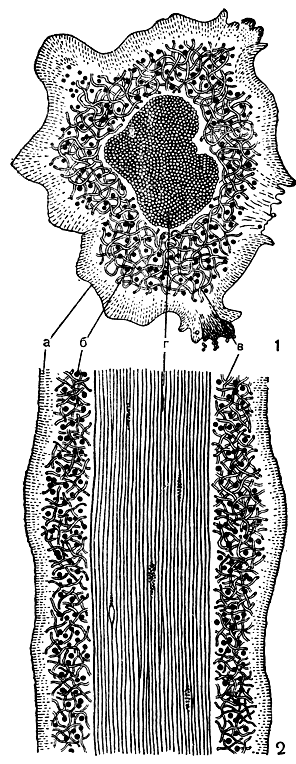
Рис. 296. Поперечный (1) и продольный (2) разрезы через лопасть кустистого лишайника уснея (Usnea): а - коровой слой; б - клетки водорослей; в - периферическая часть сердцевины, состоящая из рыхлорасположенных гиф; г - центральная часть сердцевины - осевой тяж
Интересно, что в Антарктике наиболее широко распространенными кустистыми лишайниками являются виды рода нейропогон (Neuropogon). Этот род очень близок к уснеям, его представители также имеют округлые лопасти с осевым тяжем в центральной части.
Желтовато-зеленоватое, местами зачерненное слоевище этих лишайников имеет вид сильно разветвленного компактного кустика высотой 2-10 см, плотно прикрепляющегося к субстрату расширенной пяточкой псевдогомфа (табл. 49,6). Благодаря морфологическим особенностям строения слоевищ эти лишайники хорошо приспособились к жизни в условиях холодных пустынь Антарктики. Механический тяж, придающий прочность и устойчивость ветвям, и прочное прикрепление к поверхности скал посредством псевдогомфа позволяют этим прямостоячим кустистым слоевищам хорошо противостоять постоянным сильным антарктическим ветрам.
Зачерненные местами ветви этих лишайников и их черные плодовые тела хорошо притягивают тепловые лучи, что также является важной морфологической особенностью для растений, живущих в условиях ледяных пустынь. Виды рода нейропогон играют значительную роль в растительном покрове Антарктики. В прибрежных районах в некоторых местах на скалах они образуют сплошные заросли, иногда к ним присоединяются другие лишайники и мхи, и тогда они образуют сообщества растений, сплошь покрывающие субстрат.
В Антарктике широко распространена своеобразная стелющаяся форма кустистых лишайников, представленная в этом районе видами рода алектория. Темно-коричневые, почти черные слоевища этих лишайников, состоящие из округлых, местами деформированных лопастей, горизонтально распростерты на поверхности скал и мелких камней. Это типично ветроустойчивая форма кустистых лишайников: здесь слоевища не только плотно прижимаются к субстрату, но каждая веточка такого слоевища на своем протяжении в нескольких местах еще прикрепляется к скалам маленькими пяточками. Лопасти этих стелющихся лишайников имеют множество вторичных веточек, а все слоевище напоминает по внешнему виду плотную корку, состоящую из клубков перепутанных деформированных округлых лопастей. Морфологические особенности этих лишайников позволяют им хорошо приспособиться к жизни в условиях ледяного континента. Кустистые виды алектории со стелющейся формой слоевища чрезвычайно широко распространены в Антарктике; они встречаются не только в прибрежных, более благоприятных районах, но и заходят в глубину материка, поднимаются высоко в горы, где условия для жизни растений особенно суровы. Эти лишайники были найдены в крайних пределах существования растительности на земном шаре, всего в 500 км от Южного полюса.
Иногда у крупных кустистых лишайников в условиях тундр и высокогорий развиваются добавочные прикрепительные органы, с помощью которых они прирастают к листьям осок, злаков, к веточкам карликовых берез и другим кустарничкам. Эти добавочные органы прикрепления, называемые гаптерами, могут возникнуть из любой части слоевища. Например, у цетрарии исландской они нередко развиваются из ресничек, окаймляющих края лопастей (табл. 48,5). Если слоевище цетрарии исландской растет вблизи веточки какого-нибудь кустарника, листа злака, мха или даже слоевища другого лишайника и одна из его ресничек долгое время находится в соприкосновении с ними, то такая ресничка начинает вытягиваться и на ее конце возникает плоская пластиночка. С помощью этой пластиночки лишайник прикрепляется к растущему по соседству растению. Причем развитие гаптер и прикрепление слоевища к новому субстрату происходят довольно быстро. Обычно для их формирования достаточно одного вегетационного периода. Таким образом лишайники в суровых условиях северных и высокогорных тундр предохраняют себя от отрыва сильными ветрами и бурями.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'