Анатомия слоевища лишайников
У лишайников в зависимости от анатомического строения различают два типа слоевищ: 1) гомеомерное слоевище, когда водоросли разбросаны по всей толще слоевища; 2) гетеромерное слоевище, когда водоросли образуют в слоевище обособленный слой.
Более примитивным считается слоевище гомеомерного строения. Если рассмотреть поперечный срез такого слоевища под микроскопом, то хорошо видно, что оно образовано беспорядочными переплетениями гиф гриба, среди которых разбросаны отдельные клетки или нити водорослей (рис. 297, 2, 3). Такое строение наиболее характерно для тех лишайников, фикобионтом которых являются сине-зеленые водоросли - носток, глеокапса и некоторые другие. Эти лишайники образуют группу, известную под названием слизистых лишайников. У лишайников, которые имеют фикобионтом зеленые водоросли, слоевище гомеомерного строения встречается очень редко, лишь у наиболее примитивных накипных форм.

Рис. 297. Анатомическое строение слоевища лишайников: 1 - гетеромерное слоевище (а - верхний коровой слой, б - слой водорослей, в - сердцевина, г - нижний коровой слой); 2 - гомеомерное слоевище слизистого лишайника коллема (Collema flaccidum); 3 - гомеомерное слоевище слизистого лишайника лептогиум (Leptogium saturninum) (а - коровой слой с верхней и нижней стороны слоевища, б - ризоиды)
Слоевища слизистых лишайников в сухом состоянии имеют вид темных или даже черных твердых и хрупких корок или пленок. Но они обладают одной интересной особенностью - способностью впитывать огромное количество воды, в 20-30 раз превышающее их собствен-ную массу. При увлажнении они сильно разбухают, ослизняются, приобретают характер студня или желе и становятся оливково-зелеными или грязно-зелеными. Примером может служить влажное слоевище слизистого лишайника лептогиума (Leptogium saturninum), изображенное на цветной таблице 47. Во внешнем облике этих лишайников имеется много общего с некоторыми свободноживущими водорослями. Например, у видов рода коллема (Collema), типичных представителей слизистых лишайников, форма и цвет слоевища очень напоминают черные пленки колоний свободноживущей водоросли носток. Иногда кажется, что в таких слоевищах не грибные гифы образуют переплетения, среди которых разбросаны нити водорослей, а просто в колонии водорослей проникли гифы гриба. Действительно, на формирование внешнего облика этих слоевищ оказывают большое влияние морфологические особенности их фикобионта - сине-зеленых водорослей. Для клеток этих во-дорослей чрезвычайно характерны слизистые желатинные образования, которые формируются чаще всего в результате ослизнения толстых оболочек клеток или как продукт выделения их содержимого. До последнего времени даже считали, что сине-зеленые водоросли пол-ностью определяют форму слоевища у большинства слизистых лишайников. Однако поляризационно-оптические исследования, проведенные в последние годы, показали, что слизистая масса этих слоевищ образована не только водорослью, но и грибом. Значительная часть желатина этих лишайников продуцируется грибным компонентом. Гриб определяет размеры слоевища, формирует лопасти и основные детали общего вида.
Среди слизистых лишайников встречаются как накипные, так и листоватые и кустистые формы. Но, в отличие от других лишайников, у слизистых усложнение морфологического строения почти не приводит к анатомическим преобразованиям в слоевище. Как у накипных, так и у более высокоразвитых листоватых и кустистых форм этих лишайников структура слоевища гомеомерная, т. е. водоросли всегда разбросаны по всей толще слоевища. Лишь у видов рода лептогиум (Leptogium) - высокоорганизованных форм слизистых лишайников - с верхней и нижней стороны слоевища имеется однослойный крупноклеточный коровой слой (рис. 297, 3). У наиболее эволюционно продвинутых представителей этого рода все слоевище целиком состоит из крупноклеточной ткани, но среди этих клеток разбросаны в беспорядке нити ностока.
Слизистые лишайники насчитывают небольшое число видов, всего 750, что составляет только 3% от общего числа всех известных видов лишайников. Сравнительно небольшое количество этих лишайников, по-видимому, можно объяснить более поздним вовлечением сине-зеленых водорослей в процесс образования слоевища лишайников. Но также возможно, что захват грибом сине-зеленых водорослей был менее успешным по сравнению с зелеными водорослями. В силу особенностей строения сине-зеленых водорослей, нередко собранных в колонии и обладающих плотными, богатыми слизью оболочками, эти водоросли оказались более трудным объектом для гриба при установлении с ними контакта и при построении слоевища. Безусловно, при захвате этих водорослей и их передвижении гриб испытывает гораздо больше трудностей, чем с одноклеточными и другими формами зеленых водорослей. Может быть, именно этим можно объяснить и примитивное анатомическое строение слизистых лишайников.
Для остального огромного большинства лишайников характерна гетеромерная структура, при которой в слоевищах можно различить дифференцированные слои. При этом чем сложнее морфологическое строение слоевища лишайника, тем сложнее и его анатомическая структура. У этих растений в ходе эволюции усложнение морфологического строения слоевища шло параллельно усложнению его анатомической структуры. Так, в слоевище накипных лишайников, наиболее примитивных форм, можно различить только три анатомических слоя: коровой слой, слой водорослей и сердцевину. Такое же анатомическое строение имеют и некоторые листоватые лишайники, например представители рода пельтигера (рис. 299,1). Но у большинства листоватых лишайников, более эволюционно продвинутых форм, в связи с отделением от поверхности субстрата происходит образование еще одного корового слоя - с нижней стороны слоевища. В таких слоевищах можно различить уже четыре слоя: верхний коровой слой, слой водорослей, сердцевину и нижний коровой слой (рис. 297, 1). У кустистых лишайников с плоскими лентовидными лопастями образуется еще один анатомический слой - слой водорослей с нижней стороны слоевища. У этих лишайников в слоевище можно различить уже пять анатомических слоев: два коровых слоя на верхней и нижней поверхности слоевища, два слоя водорослей, тоже с верхней и нижней стороны лопастей, и сердцевину (рис. 298). И наконец, кустистые лишайники с округлыми лопастями, обладающие наиболее высокоорганизованными слоевищами, имеют радиальную структуру: снаружи лопасти этих лишайников покрыты коровым слоем, под ним расположен слой водорослей, а центральная часть слоевища заполнена сердцевиной (рис. 295).

Рис. 298. Поперечный разрез через лопасть кустистого лишайника цетрарии исландской (Cetraria islandica): а - верхний коровой слой, б - верхний слой водорослей, в - сердцевина, г - нижний слой водорослей, д - нижний коровой слой, е - псевдоцифелла
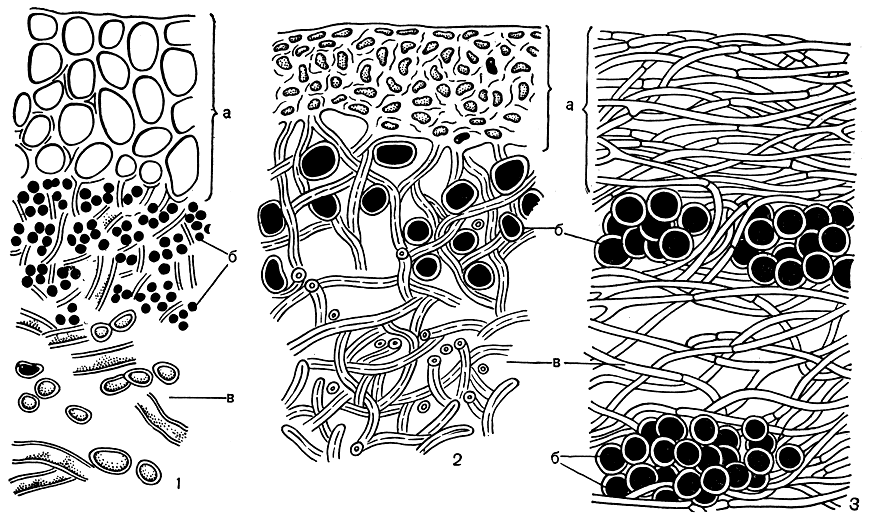
Рис. 299. Строение норового слоя лишайников: 1 - параплектенхимный коровой слой (Peltigera); 2 - прозоплектенхимный коровой слой (Parmelia); 3 - волокнистый коровой слой (Teloschistes). а - коровой слой, б - клетки водоросли, в - сердцевинные гифы
Каждый из перечисленных анатомических слоев слоевища выполняет в жизни лишайника ту или иную функцию и в зависимости от этого имеет совершенно определенное строение.
Коровой слой играет в жизни лишайника очень важную роль. Он выполняет сразу две функции: защитную и укрепляющую. Он защищает внутренние слои слоевища от воздействия внешней среды, прежде всего водоросли от чрезмерного освещения. Поэтому коровой слой лишайников обычно бывает плотного строения и окрашен в сероватый, коричневый, оливковый, желтый, оранжевый или красноватый цвет.
Коровой слой служит и для укрепления слоевища. Чем выше слоевище поднимается над субстратом, тем более оно нуждается в укреплении. Укрепляющие механические функции в таких случаях нередко выполняет толстый коровой слой. Гифы корового слоя срастаются своими стенками и могут образовывать очень плотные и сложные плектенхимы - клеточные ткани различного строения. Так, например, если срастаются тонкостенные гифы, поделенные поперечными перегородками на одинаковые по ширине и длине клетки, то образуется ткань настоящей клеточной структуры (рис. 299, 1). Такую ткань называют параплектенхимой. Иногда ее еще называют псевдопаренхимой, так как она по внешнему виду напоминает паренхиму (клеточную ткань) цветковых растений. Однако сходство этих тканей только внешнее, ибо процесс их образования различен. Если паренхима цветковых растений образуется путем последовательного деления одной клетки, то псевдопаренхима лишайников - путем срастания грибных гиф.
У многих лишайников в коровом слое оболочки клеток гиф бывают сильно утолщены, а просветы клеток, заполненные протоплазмой, очень узкими. При срастании таких гиф образуется коровой слой, представляющий сплошную хрящевидную массу, где границы клеток незаметны, а их просветы кажутся почти точками. При этом гифы на своей поверхности выделяют желатин, плотна цементирующий их между собой. Такого типа ткани у лишайников называют прозоплектенхиматическими. Так устроен коровой слой многих кустистых лишайников, например цетрарии исландской, листоватых лишайников рода пармелия и многих других (рис. 299, 2). Плотный хрящевидный коровой слой этих лишайников не только надежно защищает нижележащие слои водорослей, но и хорошо укрепляет приподнимающееся над землей слоевище.
Реже гифы корового слоя лишайников не срастаются, а лежат свободно параллельно друг другу. При этом они могут образовывать два различных типа корового слоя. Если гифы расположены параллельными рядами перпендикулярно поверхности слоевища, то образуется палисадный коровой слой (рис. 300,2).

Рис. 300. Расположение гиф в слоевище лишайников: 1 - сердцевинный слой в слоевище анаптихии; 2 - палисадный коровой слой в слоевище тамнолии. а - коровой слой, б - клетки водорослей, в - сердцевинные гифы
По внешнему виду он напоминает палисадную клеточную ткань в листьях цветковых растений. Такое строение имеет, например, коровой слой тамнолии (табл. 48, 3). Во втором случае (например, У уснеи) гифы корового слоя лежат не перпендикулярно, а параллельно поверхности слоевища и имеют вид удлиненных волокон (рис. 299, 3). Коровой слой такого строения носит название волокнистого.
В слоевищах лишайников верхний и нижний коровые слои могут быть одинаковыми по окраске и строению, но нередко отличаются друг от друга, особенно у листоватых лишайников, для которых характерно дорсовентральное строение слоевища.
На нижнем коровом слое лишайников обычно образуются органы прикрепления. Иногда они имеют вид очень тонких нитей, состоящих из одного ряда клеток. Эти нити называют ризоидами. Каждая такая нить берет начало от одной клетки нижнего корового слоя (рис. 301 ,1). Нередко несколько ризоидов соединяются в толстые ризоидальные тяжи. Их можно видеть, например, на нижней стороне крупнолистоватых слоевищ пельтигер. У листоватых лишайников рода пармелия образуются более толстые прикрепительные тяжи, назы-ваемые ризинами (рис. 301, 2). В образовании ризин принимают участие не только гифы нижнего корового слоя, но и сердцевины. Снаружи они покрыты коровым слоем, а внутри образованы сердцевинными гифами. На самом конце ризины клетки гиф вытягиваются и расходятся в разные стороны, образуя кисточку. Нередко на конце такой кисточки образуется слизистая капелька, с помощью которой слоевище более плотно прикрепляется к субстрату (рис. 301, 3). Иногда на конце ризины образуется особая прикрепительная пластиночка, которой лишайник, как маленькой подошвой, прикрепляется к поверхности коры дерева или камня.
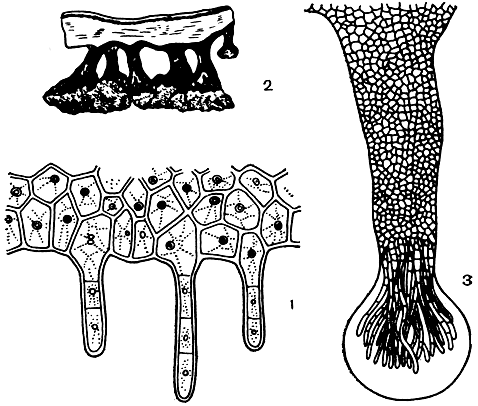
Рис. 301. Органы прикрепления слоевища листоватых лишайников: 1 - нижний параплектенхимный коровой слой с отходящими от него ризоидами (Sticta); 2 - ризины пармелии с прикрепительными пластинками на конце; 3 - ризина с капелькой слизи на конце
Как уже отмечалось, у некоторых листоватых лишайников, имеющих вид более или менее округлой листовидной пластинки, слоевище прикрепляется лишь в своей центральной части с помощью короткой ножки - гомфа (рис. 292). Гомф - это также вырост нижней поверх-ности слоевища, и в его образовании принимают участие нижний коровой слой и сердцевина лишайника. Снаружи эта толстая ножка покрыта параплектенхимным коровым слоем, а внутри ее проходят вытянутые сердцевинные гифы. На конце гомфа, на месте прикрепления к субстрату, эти гифы имеют очень толстые темные оболочки и образуют зубцы, с помощью которых слоевище очень плотно прикрепляется к поверхности скал. Некоторые кустистые лишайники, виды родов уснея, нейропогон и др., прикрепляются к скалам с помощью псевдогомфа, также имеющего вид короткой расширенной на конце ножки. В отличие от гомфа в образовании псевдогомфа принимает участие только сердцевина лишайников. Псевдогомф образован вытянутыми, склеенными сердцевинными гифами, также образующими на конце темные зубцы, с помощью которых лишайник плотно прикрепляется к субстрату.
В зоне водорослей осуществляются процессы ассимиляции углекислоты и накопление органических веществ. Как известно, для осуществления процессов фотосинтеза водорослям необходим солнечный свет. Поэтому слой водорослей обычно размещается вблизи верхней поверхности слоевища, непосредственно под верхним коровым слоем, а у вертикально стоящих кустистых лишайников еще и над нижним коровым слоем. Слой водорослей чаще всего бывает небольшой толщины, и водоросли размещаются в нем так, что находятся почти в одинаковых условиях освещения. Водоросли в слое-вище лишайника могут образовывать непрерывный слой, но иногда гифы микобионта делят его на отдельные участки. Для осуществления процессов ассимиляции углекислоты и дыхания водорослям необходим также нормальный газообмен. Поэтому грибные гифы в зоне водорослей не образуют плотных сплетений, а расположены рыхло на некотором расстоянии друг от друга. Лишь у некоторых пустынных лишайников водоросли окружены плотной грибной тканью клеточного строения, которая защищает их от жаркого и яркого пустынного солнца. Грибные гифы, окружающие водоросли, обычно являются ответвлениями или вершинами гиф сердцевины. Но, в отличие от сердцевинных гиф, они меньшей толщины, обладают более тонкими стенками и часто бывают поделены поперечными перегородками на множество клеток. Иногда, срастаясь, такие гифы образуют в зоне водорослей рыхлые сетчатые переплетения.
Под слоем водорослей расположен сердцевинный слой. Обычно сердцевина по толщине значительно превышает коровой слой и зону водорослей. Особенно мощно она развита по сравнению с другими слоями у накипных лишайников. От степени развития сердцевины зависит толщина самого слоевища.
Основная функция сердцевинного слоя - проведение воздуха к клеткам водорослей, содержащим хлорофилл. Поэтому для большинства лишайников характерно рыхлое расположение гиф в сердцевине (рис. 300, 1). Воздух, попадающий в слоевище, легко проникает к водорослям по промежуткам между гифами.
Сердцевинные гифы слабоветвисты, с редкими поперечными перегородками, с гладкими, слабожелатинообразными толстыми стенками и довольно узким просветом, заполненным протоплазмой. У большинства лишайников сердцевина белая, так как гифы сердцевинного слоя бесцветны. Хотя на их поверхности, как правило, откладываются кристаллы лишайниковых веществ, но в преобладающем большинстве они бесцветны и не изменяют окраски сердцевины. Если кристаллы лишайниковых веществ окрашены в тот или иной цвет, то в зависимости от этого и сердцевинный слой приобретает золотисто-желтую, кроваво-красную, серую или другую окраску. Лишайниковые вещества обладают одной важной особенностью: они нерастворимы или очень слабо растворимы в холодной воде. Благодаря этому свойству кристаллы, покрывая поверхность сердцевинных гиф, препятствуют их смачиванию. Поэтому даже во влажном слоевище лишайника его сердцевинный слой может частично оставаться сухим и содержать воздух, необходимый для клеток водорослей.
У некоторых кустистых лишайников, слоевище которых далеко отстает от субстрата, сердцевинный слой, помимо проводящей функции, выполняет еще одну - укрепляющую. Как отмечалось, у большинства лишайников укрепление слоевища происходит с помощью корового слоя, который у высокоразвитых кустистых и листоватых форм постепенно становится все более толстым и плотным. Однако дальнейшее развитие в этом направлении могло бы привести к ухудшению нормальных ассимиляционных процессов: очень толстый и плотный коровой слой препятствовал бы проникновению в слоевище воздуха и света. Поэтому в процессе развития кустистых форм происходит постепенное снижение роли корового слоя в укреплении слоевища и образование механической ткани под зоной водорослей, т. е. в сердцевинном слое.
В сердцевинном слое высокоразвитых кустистых лишайников возникают тяжи, которые расположены вдоль ветвей слоевища и состоят из толстостенных, плотно сросшихся друг с другом гиф. У некоторых лишайников, например из рода летария (Letharia), эти тяжи имеют небольшую толщину; обычно их образуется сразу несколько в сердцевинном слое. Но постепенно в процессе развития кустистых форм происходит слияние этих тяжей в один толстый и плотный центральный тяж. Особенно хорошо развит такой осевой тяж у видов рода уснея, кустистых лишайников, свисающих длинными редкими "бородами" со стволов и ветвей деревьев. Округлые веточки их слоевищ очень тонкие и нежные, но при этом достигают значительной длины, до 10-30 см, а иногда даже до 8 м. К субстрату эти веточки прикрепляются у самого основания; вся остальная часть остается свободной, повисающей. Если бы внутри таких веточек не было плотного осевого тяжа, малейшее дуновение ветра легко могло бы их порвать. Коровой слой усней тонок и непрочен. Если попытаться осторожно растянуть веточку какой-либо уснеи, то можно увидеть, как при этом наружный коровой слой легко разорвется, а в образовавшейся поперечной трещинке обнажится плотный белый тяж. Такой тяж, состоящий из толстостенных с длинными клетками гиф, плотно склеенных хрящевидным веществом, очень прочен и эластичен. Например, тоненькая нежная веточка уснеи (табл. 49, 8) длиной 8 см способна выдержать груз до 300 г.
Как же воздух проникает в слоевище лишайника? У некоторых форм на поверхности слоевища вовсе не образуется корового слоя, например у кладоний группы ягелей (рис. 295, 2). Поверхность их слоевищ образована рыхлыми сердцевинными гифами. В таком случае воздух беспрепятственно по промежуткам между рыхло-расположенными гифами проникает к водорослям, которые разбросаны отдельными группами вблизи верхней поверхности подециев. У крупнолистоватых лишайников рода пельтигера на нижней стороне слоевищ также не развивается коровой слой, ее образуют рыхло-расположенные сердцевинные гифы. И у этих лишайников воздух свободно проникает в сердцевину и далее по промежуткам между гифами к клеткам водорослей. Но такие случаи являются исключением. У большинства лишайников на поверхности слоевища образуется плотный коровой слой и доступ воздуха во внутренние слои слоевища сильно затруднен. Для осуществления нормального газообмена на поверхности слоевища лишайников образуются особые отверстия - специальные органы, которые служат для аэрации внутренних частей растения.
Например, у тундрового кустистого лишайника цетрарии исландской слоевище покрыто со всех сторон плотным блестящим коровым слоем. Он развивается в результате слияния между собой толстостенных гиф, образующих сплошную хрящевидную массу, в которой лишь кое-где видны просветы клеток неправильной формы (рис. 298, а, д). Проникнуть сквозь такой плотный желатинообразный слой воздух не имел бы никакой возможности, не будь в этом слое особых отверстий. Если внимательно присмотреться к этому лишайнику, то на нижней, всегда более бледноокрашенной поверхности его лопастей даже невооруженным глазом можно заметить беленькие пятнышки неправильной формы (табл. 48, 5). Это разрывы коры, через которые воздух, как через маленькие форточки, проникает внутрь слоевища. Они являются органами газообмена лишайников и носят название псевдоцифелл (или иногда у этого лишайника их называют еще манулами). На поперечном срезе этого лишайника, изображенном на рисунке 298, видно, что в местах образования псевдоцифеллы плотный коровой слой исчезает, а возникшее при этом отверстие заполняется рыхлорасположенными простыми или ветвистыми удлиненными гифами сердцевины. Через промежутки между этими гифами воздух проходит внутрь слоевища. У другого тундрового лишайника - цетрарии сглаженной (Cetraria laevigata), по внешнему облику очень похожей на цетрарию исландскую, псевдоцифеллы имеют иную форму. Верхняя и нижняя поверхности лопастей цетрарии совершенно гладкие, блестящие, без белых пятнышек. Но внимательно присмотревшись, можно увидеть по самому краю трубчато свернутых лопастей узенькую белую кайму. Это также псевдоцифеллы, которые образуются в результате разрыва коры вдоль края лопастей.
Помимо псевдоцифелл, у лишайников образуются и другие органы газообмена. Так, на нижней поверхности листоватых лишайников рода стикта можно заметить круглые правильной формы белые углубления (табл. 47, 4). Эти разрывы нижней коры лишайника, по форме напоминающие маленькие чашечки, называют цифеллами. Дно их выстлано рыхлорасположенными, шарообразными клетками сердцевины, через промежутки между которыми воздух легко проникает в сердцевину (рис. 302). У менегации продырявленной (Меnegazzia pertusa) вся верхняя поверхность слоевища покрыта округлыми или овальными отверстиями, которые носят название перфораций (табл. 52, 2). Их назначение - тоже проводить воздух внутрь слоевища. Перфорации возникают в результате отмирания маленьких участков корового слоя лишайников. У представителей гипогимний перфорации размещены на вершинах лопастей. У некоторых лишайников органы аэрации слоевища имеют вид маленьких точковидных пор, расположенных в нижней коре лишайников. Иногда такие поры возникают на вершине бородавочек, разбросанных на верхней поверхности лишайников.
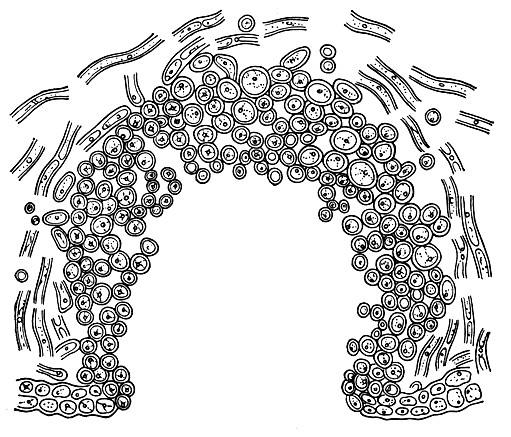
Рис. 302. Поперечный разрез через цифеллу (Sticta)
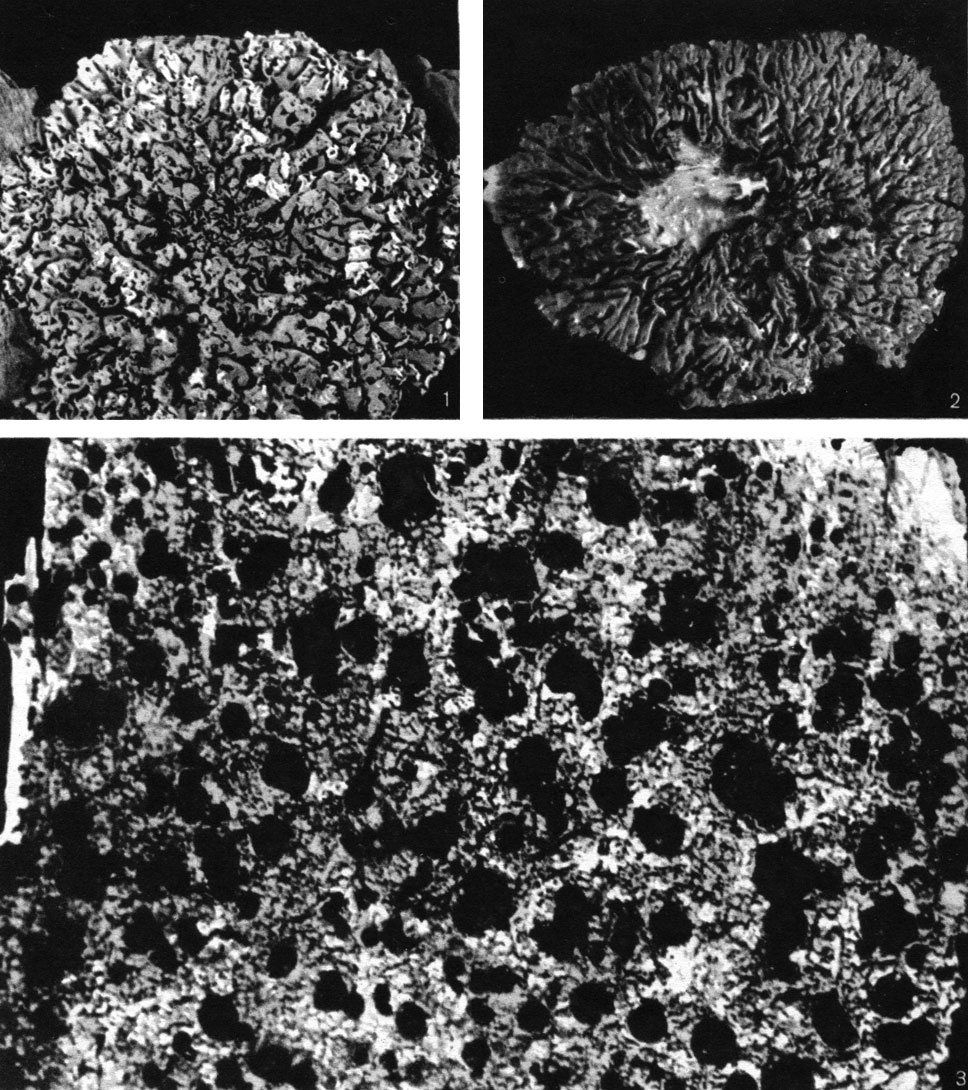
Таблица 52. Эпифитные лишайники: 1 - Hypogymnia physodes, внешний вид листоватого слоевища с губовидными соралями на концах лопастей; 2 - Menegazzia pertusa, внешний вид листоватого слоевища с округлыми отверстиями на поверхности; 3-Mycoblastus sanguinarius, внешний вид накипного слоевища с лецидеевыми апотециями
В том случае, когда на поверхности слоевища лишайников не образуются специальные органы газообмена, на помощь приходят различные трещинки и разрывы в коровом слое. Эти трещинки могут возникнуть как в результате особенностей роста слоевища, так и в результате повреждения слоевища ветром, животными, человеком.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'