Строение клетки красных водорослей
Своеобразие внутриклеточного строения красных водорослей складывается как из особенностей обычных клеточных компонентов, так и из наличия специфических внутриклеточных включений.
Клеточные оболочки. В клеточных оболочках красных водорослей хорошо различаются два слоя: внутренний, состоящий из целлюлозы, и наружный, построенный пектиновыми соединениями. У форм с грубым жестким слоевищем снаружи клеточных стенок развивается тонкий слой кутикулы. По составу и строению кутикула багрянок отличается от кутикулы высших растений. Так, например, у порфиры она образуется в результате сгущения молекул моносахарида маннозы. У мягких слизистых форм кутикула отсутствует.
Пектиновые вещества красных водорослей представляют собой соли кальция и магния особых пектиновых кислот. Они обладают способностью растворяться в кипящей воде с образованием слизистых растворов. К группе пектиновых веществ относятся также особые коллоидные вещества, которые содержатся в клеточных оболочках и межклетниках многих багрянок. Они представляют собой сложную смесь содержащих серу полисахаридов и носят общее название фикоколлоидов. Фикоколлоиды не растворяются в холодной воде, но хорошо растворяются в кипящей с образованием коллоидных систем. В настоящее время фикоколлоиды красных водорослей, так же как и бурых, являются самыми важными продуктами, получаемыми из морских водорослей. Поскольку эти вещества широко применяются в хозяйственной жизни человека, их химический состав достаточно хорошо изучен. Фикоколлоиды получены из многих видов, в результате установлено несколько их разновидностей. Более всего известны агар, каррагинин, нори, агароиды. Эти вещества отличаются друг от друга по составу и свойствам, но обладают общей желирующей способностью.
Присутствие коллоидных веществ обусловливает способность клеточных оболочек к сильному набуханию после отмирания, благодаря чему они с трудом поддаются окраске при исследовании.
Среди красных водорослей встречаются формы, оболочки которых обызвествлены. Сначала известь откладывается в срединной пластинке, между внешним и внутренним слоем оболочки, проникая постепенно в целлюлозный слой и более или менее сильно пропитывая его. Но даже при сильном обызвествлении внутри всегда остается тонкая мембрана, лишенная извести и отделяющая плазму от известкового с лая.
Состав известковых соединений неодинаков. У кораллиновых откладывается кальцит, у некоторых немалиевых - арагонит. Кроме того, встречаются карбонаты кальция и магния, а также железо.
Рост оболочек происходит следующим образом. Новые слои ее закладываются на вершине протопласта апикальной клетки, в то время как наружные слои по мере роста клетки постепенно разрываются. В результате вся оболочка имеет слоистое строение и, так как новые слои откладываются под определенным углом, по виду напоминает воронку.
Цитоплазма красных водорослей характеризуется большой вязкостью и плотно прилегает к клеточным стенкам. Она очень легко подвергается плазмолизу, и поэтому красные водоросли весьма чувствительны к изменению условий внешней среды, в частности к опреснению.
Ядро. Среди красных водорослей имеется большое число форм с одноядерными клетками. Как правило, это наиболее просто организованные формы. У высокоорганизованных багрянок клетки обычно многоядерные, за исключением молодых клеток слоевища - апикальных, коровых и т. п. Одноядерные клетки ветвей ограниченного роста при определенных состояниях становятся многоядерными. Однако можно найти и такие водоросли, у которых старые клетки центральных нитей одноядерные, а окружающие их более молодые клетки, наоборот, содержат несколько ядер. Репродуктивные клетки - спермации, карпоспоры, тетраспоры - всегда с одним ядром, но яйцеклетки часто окружены многоядерными клетками, даже у форм, все слоевище которых состоит из одноядерных клеток. Ядро у красных водорослей мелкое, имеет четкую ядерную оболочку и ядрышко.
Хлоропласты. В клетках красных водорослей находится один или несколько хлоропластов. У растений класса бангиевых и у примитивных представителей наиболее низкоорганизованного порядка немалиевых имеется единственный звездчатый хлоропласт с одним пиреноидом. Он обычно занимает осевое положение в клетке и тогда состоит из центрального тела и отходящих от него во всех направлениях отростков (рис. 149, 2). Пиреноид при осевом хлоропласте находится в его центре. Отростки, возникающие из центрального хлоропласта, могут расширяться на периферии и смыкаться между собой, образуя пристенный хлоропласт неправильной или лентовидной формы. Вероятно, пристенный хлоропласт большинства багрянок происходит от осевого в результате утраты центральной части.
Роль пиреноида у красных водорослей не очень ясна. В одних случаях его присутствие связано с отложением крахмальных зерен; в других же пиреноид встречается в клетках, не участвующих в процессах ассимиляции. У более высокоорганизованных форм пиреноид исчезает; этот процесс можно проследить уже в порядке немалиевых.
У красных водорослей, лишенных пиреноидов, хлоропласты бывают двух основных типов - лентовидные и линзовидные (или дисковидные) с многочисленными переходами между ними (рис. 154). Виды, стоящие на более низкой ступени эволюции, обладают обычно лентовидным хлоропластом; для высокоорганизованных форм, наоборот, более характерны линзовидные хлоропласты. То же самое можно сказать и о количестве хлоропластов - число их с усложнением организации увеличивается. Форма хлоропласта не есть нечто постоянное, она может меняться с возрастом, с условиями освещения, с изменением размеров клеток, хотя нередко крупные группы багрянок характеризуются хлоропластом определенной формы. У церамиума в крупных клетках междоузлий хлоропласты вытянутые, лентовидные, а в коротких коровых клетках на узлах - это короткие, неправильно лопастные пластинки. Размножение хлоропластов происходит путем простого перешнуровывания, как у высших растений.
По своему тонкому строению, видимому только под электронным микроскопом, хлоропласты красных водорослей почти не отличаются от хлоропластов других водорослей.
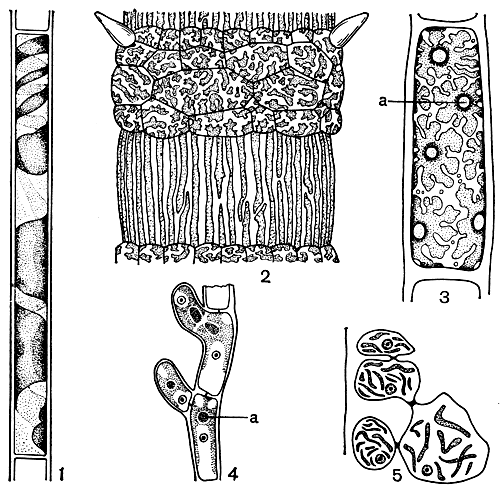
Рис. 154. Форма хлоропластов багрянок: 1 - Audouinella; 2 - Ceramium; 3-4 - Kylinia; 5 - Cystoclonium: a - пиреноид
Пигменты. Багрянки отличаются сложным набором пигментов. Кроме обычных для зеленых растений, растворимых в спирту хлорофилла, каротина и ксантофилла, хлоропласты красных водорослей содержат дополнительные водно-растворимые пигменты билипротеины. Это фикоэритрин и фикоцианин. Зеленые растения, как водоросли, так и наземные, содержат две модификации хлорофилла - сине-зеленый хлорофилл а и желто-зеленый хлорофилл b. У красных водорослей найден только хлорофилл а - универсальный пигмент, характерный для всех растений. Кроме того, у некоторых багрянок обнаружен хлорофилл d, природа которого, однако, остается до сих пор не выясненной. Зеленых пигментов у багрянок по сравнению с высшими растениями содержится немного, и обычно их маскируют дополнительные билипротеины. Замечена некоторая закономерность в изменении количества хлорофилла в зависимости от количества света. Водоросли, приспособившиеся к жизни при малой освещенности в полярных морях, обычно богаче хлорофиллом, чем водоросли южных морей. Точно так же глубоководные водоросли богаче хлорофиллом, чем растущие у поверхности воды.
Каротиноиды красных водорослей представлены α- и β-каротином и ксантофиллами лютеином, зеаксантином и, вероятно, тараксантином. Билипротеинами багрянок являются красный фикоэритрин и голубой фикоцианин. Они близки к пигментам сине-зеленых водорослей, но не идентичны им, так как отличаются по химическому составу. Как показано на многочисленных опытах, количество пигментов у багрянок возрастает с глубиной; при этом количество фикоэритрина возрастает в большей мере, чем количество хлорофилла. Каждый, кто собирал эти водоросли в природе, знает, что окрашенные в красный цвет багрянки растут на глубине и что на мелководье они меняют ок-раску. С увеличением количества света они становятся бледно-красными, затем желто-зелеными, соломенными и наконец полностью обесцвечиваются.
Существует теория так называемой хроматической адаптации, по которой проникновение водорослей на те или иные глубины связано с качеством света, проходящего через толщу воды. Как известно, глубже всего проникают лучи из зеленой и синей частей спектра. Красные пигменты багрянок позволяют им фотосинтезировать в синих лучах, и поэтому, согласно этой теории, они проникают на глубины, недоступные для других водорослей. Однако на практике эта закономерность наблюдается далеко не всегда. Какова же роль билипротеинов в фотосинтезе красных водорослей? В опытах было установлено, что при слабом освещении они участвуют в усиленном поглощении света. Поэтому их можно считать оптическими сенсибилизаторами. Таким образом, проникновение красных водорослей на значительные глубины правильнее объяснить их способностью усваивать малые количества света. В целом багрянки - теневыносливые организмы: слабый свет они способны использовать лучше, чем другие водоросли. Если красная окраска водорослей при слабом свете получает преимущества, то при более сильном, наоборот, интенсивность фотосинтеза багрянок ниже, чем у других водорослей, как раз благодаря наличию красных пигментов. Для защиты от сильного света у багрянок, живущих на небольших глубинах, особенно в тропических и субтропических морях, служат особые иридирующие тельца. Эти мутно-желтые неправильной формы тельца образуются в вакуолях поверхностных клеток слоевища и состоят из мелких зернышек протеиновой природы. Они обладают способностью рассеивать и отражать падающие на них солнечные лучи. При очень сильном освещении иридирующие тельца располагаются под внешней стенкой клетки, в то время как хлоропласт - на внутренней или боковой, и служат своеобразным занавесом для хлоропласта. При попадании растения в условия рассеянного света происходит взаимное перемещение и хлоропласт оказывается у внешней стенки.
Водоросли, обладающие иридирующими тельцами, имеют обычно в падающем свете голубовато-стальной блеск. У некоторых видов в клетках возникают крупные линзообразные тела, которые с понижением освещенности исчезают.
Запасные вещества. В качестве продукта ассимиляции у красных водорослей откладывается полисахарид, называемый багрянковым крахмалом. По химической природе он ближе всего к амилопектину и гликогену и, по-видимому, занимает промежуточное положение между обычным крахмалом и гликогеном. Откладывается багрянковый крахмал в виде мелких полутвердых телец различной формы и окраски. Эти тельца могут иметь форму конусов или плоских овальных пластинок с углублением на широкой поверхности. Часто на них можно видеть концентрические зоны. Зерна багрянкового крахмала образуются частично в цитоплазме, частично на поверхности хлоро-пластов, но они никогда не образуются внутри пластид, в отличие от обычного крахмала зеленых растений. У форм, имеющих пиреноид, последний в какой-то мере участвует в синтезе крахмала.
Кроме багрянкового крахмала, в качестве запасных веществ у красных водорослей откладываются сахара трегалоза, флоридозид, сахароза и др. У некоторых форм в изобилии встречаются многоатомные спирты. Из жиров известны холестерол, силостерол, фукостерол. Содержание жиров меняется в зависимости от условий среды.
Железистые клетки. Особенностью красных водорослей является наличие у некоторых представителей класса флоридеевых особых клеток с бесцветным содержимым, сильно преломляющим свет (рис. 155). В литературе они известны как пузырчатые, или железистые, клетки. Содержимое этих клеток у разных водорослей имеет разную природу; они заполнены иодистыми, реже бромистыми, соединениями. Чаще всего железистые клетки встречаются в порядке церамиевых. У нитчатой разветвленной водоросли антитамнион они сидят на верхней стороне боковых ветвей (рис. 155, 1). При их развитии сначала отчленяется маленькая линзообразная клетка, содержащая небольшое количество плазмы и мелкие красные хлоропласты. Ядро можно проследить лишь на самой ранней стадии развития. Вскоре на дне этой клетки образуется бесцветный светопреломляющий пузырь. Он растет, и вместе с ним увеличивается в размерах вся клетка. В сформировавшейся клетке большая часть занята пузырем, и только в верхней части остается узкий слой плазмы с мелкими хлоропластами (рис. 155, 2-5). Роль пузырчатых клеток не выяснена, хотя на этот счет имеется множество самых различных предположений. Их считают недоразвитыми спорангиями, хранилищем запасных веществ, "плавательными пузырями" и т. п.
Железистые клетки характерны для определенных групп водорослей и поэтому служат важным таксономическим признаком.
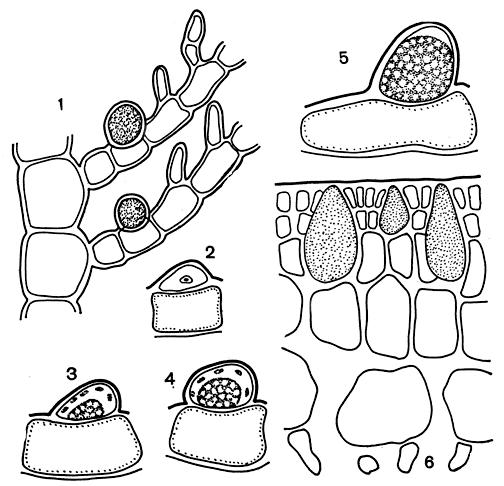
Рис. 155. Железистые клетки красных водорослей: 1 - веточка Antithamnion с железистыми клетками; 2-5 - стадии развития железистой клетки Antithamnion; 6 - поперечный срез Opuntiella с железистыми клетками
Волоски. Образование волосков - широко распространенное явление в классе флоридеевых. Настоящие волоски багрянок следует отличать от волосовидных ветвей или ложных волосков. У видов с однорядным нитчатым слоевищем можно видеть, как конечные клетки боковых веточек удлиняются и обесцвечиваются, приобретая волосовидное строение (рис. 151, 1). Это и есть ложные волоски. Настоящие волоски красных водорослей делятся на два типа: одноклеточные и многоклеточные. Одноклеточные волоски никогда не ветвятся. У однорядных нитчатых форм они образуются из верхушечных клеток ветвей, у многорядных - из поверхностных клеток коры. Клетка будущего волоска отделяется от материнской клетки поперечной перегородкой и сильно вытягивается, достигая в длину нередко миллиметра и более (рис. 156). Она содержит ядро и небольшое количество цитоплазмы. Хлоропласт по мере роста волоска исчезает, и волоски становятся бесцветными. Обычно материнская клетка волоска ничем не отличается от соседних вегетативных клеток, но иногда она намного крупнее и остается хорошо заметной после отпадения волоска. У кораллиновых волоски не являются самостоятельными клетками, а представляют собой только выросты специальных клеток, от которых они не отделяются перегородкой. Эти клетки намного крупнее остальных и известны под названием трихоцитов или гетероцист (рис. 172, 2, 3). Если в классе флоридеевых одноклеточные волоски встречаются довольно часто, то у бангиевых они отсутствуют полностью.
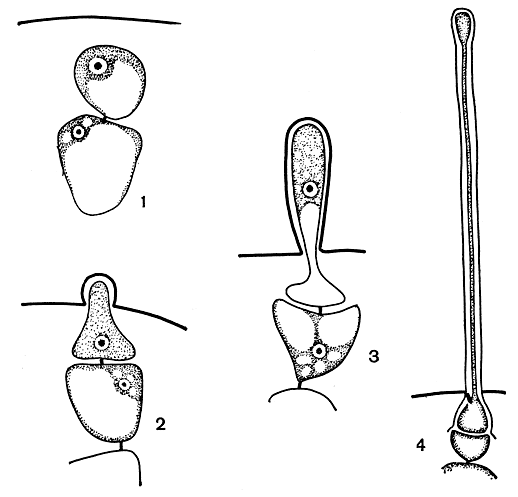
Рис. 156. Стадии развития волоска у Cystoclonium
Многоклеточные волоски обычно более или менее сильно разветвлены. Они встречаются только у некоторых водорослей из порядка церамиевых. Как было показано в опытах, основная роль волосков заключается в том, что они способствуют поглощению питательных веществ из окружающей среды.
Поры. Одна из наиболее интересных особенностей красных водорослей состоит в том, что клетки, слагающие слоевище, соединяются между собой с помощью специальных образований, называемых порами. Между дочерними клетками, т. е. клетками, происходящими от одной материнской, соединение осуществляется посредством первичных пор (рис. 157,1-3). Они формируются в результате неполной перегородки между двумя вновь образующимися клетками. Первичные поры расположены в середине перегородки, в точке, через которую можно провести линию, соединяющую ядра дочерних клеток, и представляют собой тонкую пластинку. Через эту пластинку проходят тяжи, которые соединяют цитоплазму соседних клеток. До самого последнего времени считалось, что первичные поры свойственны только флоридеевым и отсутствуют в классе бангиевых - это был один из принципиальных признаков, на основании которого различали оба эти класса. Но недавно первичные поры были открыты и у представителей класса бангиевых.
Кроме соединения дочерних клеток, у большинства флоридеевых происходит вторичное соединение соседних клеток. Это могут быть клетки одной и той же нити, лежащие рядом клетки соседних нитей или - у паразитических багрянок - клетки паразита и хозяина. Способностью к вторичному соединению обладают более высокоорганизованные представители класса. Она отсутствует в порядке немалиевых и у примитивных водорослей других порядков. Соединение соседних клеток, между которыми первоначально никакой связи не было, осуществляется вторичными порами (рис. 157, 4-8). Образование вторичных пор - процесс довольно сложный. Начинается он с того, что ядро в клетке делится и одно из дочерних ядер перемещается к периферии клетки. В том месте, куда попадает ядро, образуется небольшой вырост, который затем отчленяется, как маленькая клетка, соединенная с первой посредством первичной поры. Вытягиваясь, эта дополнительная клетка достигает соседней и сливается с ней. Теперь первичная пора, соединяющая дополнительную клетку с первой, становится вторичной. Иногда две соседние клетки соединены несколькими вторичными порами. Роль этих соединений до конца не выяснена; вероятно, они способствуют переносу питательных веществ от одной клетки к другой. Хотя способ образования первичных и вторичных пор различен, строение их сходно. И все же не у всех багрянок поры устроены одинаково. В одних случаях между клетками имеется непосредственная цитоплазматическая связь, в других - она гораздо менее очевидна.
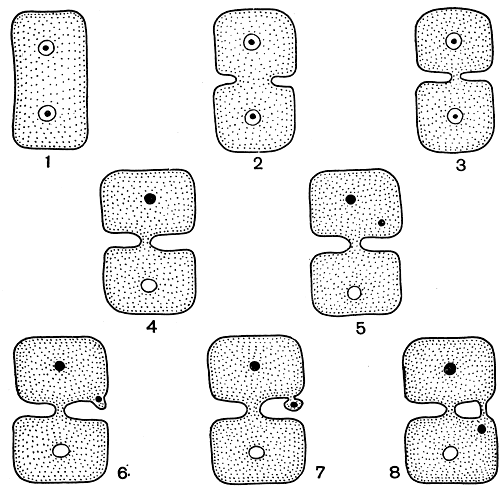
Рис. 157. Схема образования первичных (1-3) и вторичных (4-8) пор у красных водорослей
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'