Размножение красных водорослей
Размножение красных водорослей чрезвычайно сложный и многообразный процесс, отличающий их от других групп водорослей. Различные формы бесполого размножения, сложное строение половой системы, особенно женского репродуктивного органа, особенности развития зиготы, а также многообразие циклов развития - все эти признаки не только придают своеобразие отделу Bhodophyta, но являются основой, на которой строится классификация багрянок, их разделение на порядки, семейства, роды. Поэтому, чтобы составить целостное представление о красных водорослях, нельзя не остановиться на особенностях их размножения.
Собственно говоря, сложным размножением характеризуются представители класса флоридеевых; у бангиевых строение репродуктивной системы намного проще.
Вегетативное размножение флоридеевых происходит посредством образования дополнительных побегов, которые берут начало от подошвы или от стелющихся ветвей. Вертикальное слоевище в определенный период отмирает и остается только базальная часть растения, которая через какое-то время прорастает, образуя новые побеги. Самые примитивные багрянки из класса бангиевых размножаются только вегетативно. У одноклеточных и колониальных форм вегетативное размножение состоит в делении клетки на две или более дочерние клетки.
Бесполое размножение. Для бесполого размножения служат различного рода споры. У низкоорганизованных багрянок бесполое размножение осуществляется моноспорами. Они характерны для всех представителей класса бангиевых, тогда как среди флоридеевых известны за очень редкими исключениями только в порядке немалиевых. Моноспорами называют споры, которые образуются по одной в клетке в результате превращения всего протопласта. Моноспоры не имеют жгутика и оболочки, после выхода из материнской клетки они способны к амебоидному движению. У бангиевых моноспоры образуются в любой клетке слоевища и до своего выхода не отличаются от вегетативных клеток. В классе флоридеевых (там, где они встречаются) моноспорангии образуются на веточках ограниченного роста. У водорослей, построенных по типу свободных нитей, в моноспорангии превращаются конечные клетки коротких боковых веточек. Иногда они образуются группами по два-три на одной и той же клетке-ножке (рис. 151, 1). У водорослей, имеющих более или менее плотное строение и дифференцированный коровой слой, в моноспорангии превращаются внешние клетки ассимиляционных нитей. Моноспоры отличаются от вегетативных клеток яйцевидной или шаровидной формой, богатым содержимым и интенсивной окраской. Созревшие моноспорангии открываются на вершине; голые моноспоры выходят в окружающую среду, через некоторое время одеваются оболочкой и начинают прорастать.
Для большинства флоридеевых наиболее обычны тетраспоры. Они образуются по четыре в клетке, которую называют тетраспорангием. По способу деления различают три типа тетраспорангиев: крестообразно, зонально и тетраэдрически разделенные. В крестообразно разделенных тетраспорангиях вначале вся клетка делится пополам поперечной перегородкой, после чего каждая из двух образовавшихся клеток делится продольной перегородкой, при этом обе перегородки проходят в одной плоскости или под прямым углом друг к другу (рис. 158, 5, 7). В последнем случае, если смотреть на спорангий сбоку, можно различить только три клетки. При зональном делении все три перегородки проходят поперечно и параллельно друг другу, в результате чего тетраспоры располагаются в ряд (рис. 158, 4, 6). В тетраэдрически разделенных тетраспорангиях клеточные перегородки ориентированы не под прямым углом, как при крестообразном делении, а косо (рис. 158, 1).
Способ деления тетраспорангиев - постоянный признак, который закрепился в эволюции за той или иной группой. В порядках немалиевых и гелидиевых имеются только крестообразно разделенные спорангии, у криптонемиевых и гигартиновых - крестообразно и зонально разделенные. В порядке родимениевых встречаются как крестообразно, так и тетраэдрически разделенные спорангии. У боль-шинства церамиевых спорангии делятся тетраэдрически. Как исключение, спорангии делятся только на две части, и тогда мы говорим об образовании биспор. Наиболее характерно образование биспор для кораллиновых.
Место образования тетраспор у разных групп багрянок также постоянный признак, который используется в систематике. Как правило, тетраспоры занимают положение конечных клеток веточек ограниченного роста, будь то свободные боковые веточки у просто организованных немалиевых или коровые нити у остальных багрянок. Интеркалярное возникновение тетраспорангиев, т. е. в середине клеточных нитей, встречается исключительно редко. Однако в процессе роста тетраспорангиев - а они обычно бывают намного крупнее соседних вегетативных клеток - они оказываются погруженными в коровой слой, и их положение конечной клетки в зрелом состоянии не всегда очевидно (рис. 158, 7). Располагаются тетраспорангии поодиночке или группами. Собрания тетраспорангиев часто имеют вид цепочек. Интеркалярные тетраспорангии образуются из клеток внутренней части коры, как, например, у дилсеи, или в середине вертикальных ассимиляционных нитей у корковых форм. У корковой водоросли круории тетраспорангии представляют собой боковые выросты вертикальных нитей (рис. 158, 4). Из клеток центральной части слоевища формируются тетраспорангии у таких гигартиновых водорослей, как хондрус и иридея (рис. 158, 2). На слоевище тетраспорангии располагаются по-разному. Очень часто они рассеяны по всему слоевищу или приурочены к какой-то определенной его части, например к верхним ветвям. Но нередки случаи, когда образование тетраспорангиев связано со специальными веточками или выростами. Эти выросты располагаются по краю слоевища или на его поверхности у пластинчатых форм и вокруг основных ветвей - у цилиндрических (рис. 158, 3).
У некоторых видов тетраспорангии развиваются в так называемых нематециях. Это специальные образования на поверхности слоевища. Они состоят из вертикальных клеточных нитей, берущих начало от поверхностных коровых клеток, и имеют вид небольших подушечек. Тетраспорангии развиваются среди нитей нематеция, тесно окруженные ими. Они отчленяются от поверхностных клеток корового слоя, как, например, у сквамариевых (порядок криптонемиевых, рис. 158, 5) или от самых нижних веток нитей нематеция, как, например, у эндокладии (порядок криптонемиевых). У филлофоры и некоторых других гигартиновых все клетки нитей нематеция, за исключением самых поверхностных, превращаются в тетраспорангии. Нематеции этих водорослей имеют шаровидную форму, их долгое время считали паразитами и относили к другому роду (рис. 177, 1). У кораллиновых тетраспорангии образуются в специальных вместилищах - концептакулах (рис. 158, 6).
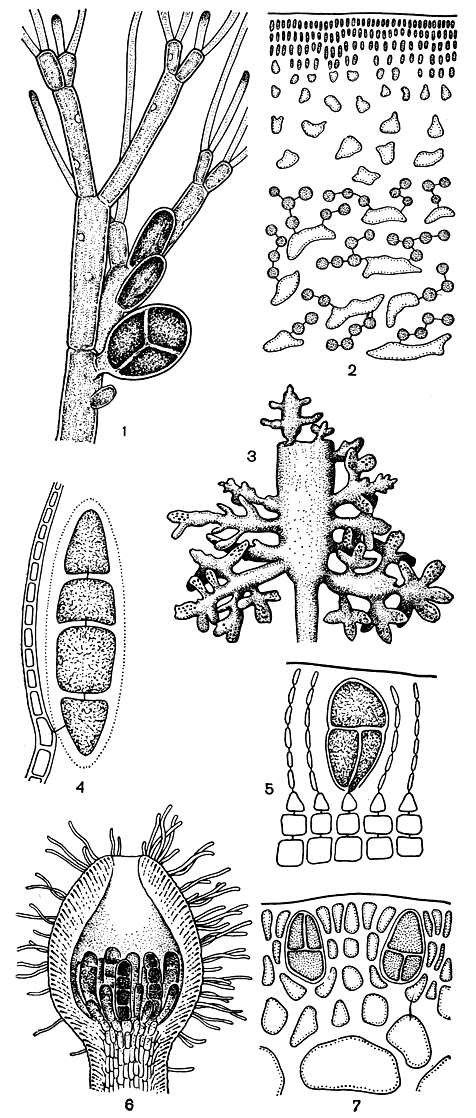
Рис. 158. Тетраспорангии красных водорослей: 1 - Callithamnion; 2 - Iridaea; 3 - веточка Gelidium с тетраспорангиями; 4 - Cruoria; 5 - Peyssonnelia; 6 - концептакул с тетраспорангиями Corallina; 7 - Gracilaria
У некоторых церамиевых и других багрянок известны случаи деления одного спорангия на много частей с образованием полиспор. Иногда встречается до 28 полиспор в одном спорангии. Они соответствуют тетраспорам и ничем, кроме количества, от них не отличаются.
Все бесполые споры багрянок лишены оболочек, у некоторых замечена способность к амебоидному движению. После выхода из спорангия они через некоторое время одеваются оболочкой, оседают и прорастают, как правило, в гаметофиты, которые несут органы полового размножения.
Половое размножение присуще всем водорослям из класса флоридеевых, тогда как в классе бангиевых оно свойственно только наиболее высокоорганизованным формам. У одноклеточных и колониальных бангиевых способность к половому размножению еще не выработалась. Для некоторых флоридеевых известно только бесполое размножение, но отсутствие полового размножения у них всегда результат вторичной утраты.
Половой процесс у всех красных водорослей оогамный. Мужские и женские гаметы лишены жгутиков. При оплодотворении женские гаметы не выходят в окружающую среду, а остаются на растении - гаметофите; мужские гаметы выбрасываются наружу и пассивно переносятся токами воды. В связи с этим женский орган размножения отличается весьма сложным строением.
Мужские гаметы, называемые у багрянок спермациями, представляют собой шаровидные или яйцевидные одноядерные клетки. Спермации бангиевых - голые клетки. Что касается флоридеевых, то здесь вопрос о наличии или отсутствии оболочки у спермациев до конца не выяснен. Известно только, что в момент оседания на женское растение они уже окружены тонкой оболочкой. Во всяком случае, способность спермациев к амебоидному движению позволяет думать, что если не у всех, то у части флоридеевых спермации на первых порах лишены оболочек.
Спермации флоридеевых, как правило, не имеют хлоропластов и бесцветны. У низших представителей класса остатки хлоропласта в мужских гаметах еще присутствуют, но они уже не играют никакой роли при оплодотворении. Постепенную редукцию хлоропласта в спермациях можно проследить уже в классе бангиевых. У простоорганизованных бангиевых, обладающих половым процессом, в спермациях развивается отчетливо видимый хлоропласт, но у высших представителей класса, например у порфиры, он уже сильно редуцирован.
Спермации образуются в клетках, которые функционируют как мужские органы размножения, называют их сперматангиями. В каждом сперматангии образуется по одному спермацию. У бангиевых сперматангии возникают в результате повторных делений многочисленных клеток нитчатого или пластинчатого слоевища. Вегетативную клетку, дающую начало сперматангиям, называют материнской. У бангиевых любая клетка слоевища, за исключением ризоидных, может стать материнской клеткой сперматангиев, хотя в природе плодоношение обычно охватывает только верхнюю часть растения. Из одной материнской клетки образуется 32-128 мелких клеток, каждая из которых дает один спермаций. У всех флоридеевых материнские клетки сперматангиев представляют собой конечные клетки веточек ограниченного роста. В соответствии с этим у флоридеевых, обладающих свободным нитчатым строением, они сидят на вершине боковых веточек (рис. 159, 1), у багрянок с плотным строением в сперматангии превращаются внешние клетки корового слоя. Каждая материнская клетка отчленяет от себя один или несколько сперматангиев (рис. 159, 2, 3). После созревания и отпадения первых сперматангиев материнская клетка может отчленить еще несколько вторичных сперматангиев. По мере усложнения организации флоридеевых материнские клетки сперматангиев также постепенно видоизменяются. Их уже можно отличить от соседних вегетативных клеток по размерам и форме, а также по отсутствию окраски, так как в них происходит редукция хлоропласта, но зато увеличивается количество цитоплазмы.
Только у небольшого числа красных водорослей сперматангии располагаются по слоевищу рассеянно. У большинства же они собраны более или менее обширными группами. Собрания сперматангиев называют сорусами. У нитевидных форм сорусы имеют вид густых пучков, нередко шаровидной формы, расположенных у вершины боковых ветвей. У водорослей с более плотным строением сорусы располагаются на поверхности слоевища в виде подушковидных образований. Иногда они оказываются погруженными в небольшие углубления на слоевище, а у пропитанных известью кораллиновых приурочены к специальным вместилищам - концептакулам (рис. 159, 5). Нередко спермации образуются в специальных нематециях. В этом случае наружные коровые клетки дают начало однорядным параллельным нитям, которые выступают над поверхностью слоевища. Затем клетки этих нитей образуют по нескольку перицентральных клеток, которые служат материнскими клетками и отчленяют каждая по нескольку сперматангиев (рис. 159, 4). У высокоорганизованных церамиевых сперматангии образуются на специальных коротких веточках - стихидиях или на трихобластах, окружая эти веточки наподобие муфты (рис. 159, 6).
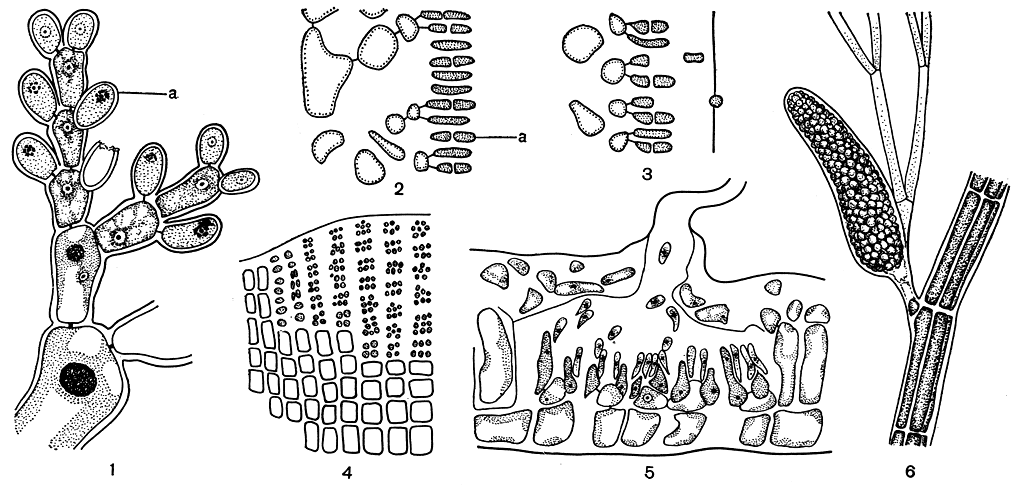
Рис. 159. Сперматангии (а) красных водорослей: 1 - Nemalion; 2,3 - Gelidium; 4 - Peyssonnelia; 5 - концептакул со сперматангиями Fosliella; 6 - Polysiphonia
Женский репродуктивный орган у красных водорослей - оогоний называют карпогоном. Это особая клетка, густо заполненная цитоплазмой и лишенная хлоропласта. Лишь у примитивных представителей флоридеевых и у бангиевых карпогон содержит окрашенный хлоропласт. Карпогон отличается весьма характерной, одинаковой для всех флоридеевых формой (рис. 160, 1). Нижняя, или брюшная, часть клетки имеет конусовидную форму, верхняя вытягивается, образуя более или менее длинный трубчатый вырост. Вырост служит для улавливания спермациев, называют его трихогиной. Благодаря трихогине карпогон легко отличить от остальных клеток слоевища. Ядро карпогона, как правило, находится в брюшной части. Трихогина может быть короткой и кеглевидной или длинной, узкой и спирально закрученной. У бангиевых собственно трихогина еще отсутствует, а иногда карпогоны имеют лишь короткие сосочкообразные выросты - прообраз будущей трихогины. Поэтому карпогон бангиевых до оплодотворения трудно отличить от обычных вегетативных клеток, тем более что им может стать любая клетка слоевища, точно так же как это происходит при образовании спермациев и моноспор (рис. 161, 1).
Далеко не у всех флоридеевых карпогон развивается как отдельная клетка. Гораздо чаще образуется короткая клеточная нить, состоящая из бесцветных клеток, заполненных, как и карпогон, гомогенной цитоплазмой, а уже собственно карпогон развивается как конечная клетка этой нити, или карпогонной ветви (рис. 160, 1). Клетку, от которой отходит карпогонная ветвь, называют несущей или базальной; в состав карпогонной ветви она не входит.
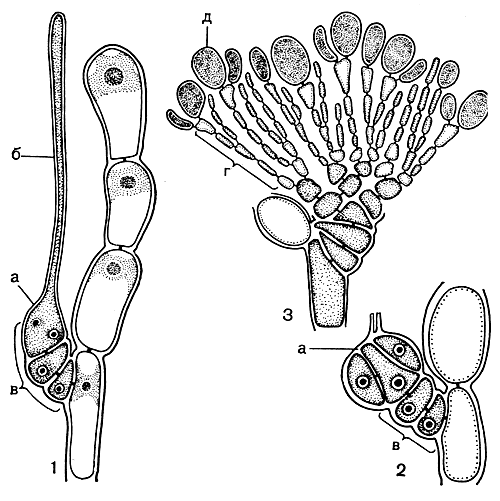
Рис. 160. Карпогонная ветвь (2) и развитие гонимобласта (2, 3) у Helminthora: а - карпогон; б - трихогина; в - карпогонная ветвь; г - нити гонимобласта; д - карпоспоры
Карпогонные ветви флоридеевых играют важную роль в процессе размножения, так как принимают активное участие в развитии карпогона после оплодотворения. Число клеток в карпогонной ветви всегда небольшое, но непостоянное у разных групп багрянок. Если проследить эти изменения от наиболее примитивных к высокоорганизованным представителям класса флоридеевых, то можно заметить, что у низкоорганизованных немалиевых и гелидиевых карпогонная ветвь еще отсутствует и имеется только одна клетка карпогона. У криптонемиевых карпогонная ветвь, будучи еще структурой малоспециализированной, характеризуется, с одной стороны, сравнительной многоклеточностью, а с другой - непостоянным числом клеток. При этом у менее специализированных форм из порядков криптонемиевых и гигартиновых встречаются даже разветвленные карпогонные ветви. В порядках родимениевых и церамиевых число клеток в карпогонной ветви уменьшается до 3-4 и становится уже более постоянным даже для крупных таксонов.
Почти у всех флоридеевых карпогонная ветвь образуется как специальная боковая веточка во внутренней части корового слоя. Только у низкоорганизованных немалиевых она соответствует обычной боковой веточке слоевища. У багрянок с плотным слоевищем карпогон оказывается погруженным, и только трихогина достигает поверхности слоевища и выступает над ней. У багрянок со свободно нитчатым строением карпогон занимает открытое положение. Строение карпогонной ветви и характер ее образования сильно варьируют у разных групп водорослей и служат одним из главных отличительных признаков в систематике багрянок.
Когда принесенные водой спермации соприкасаются с вершиной трихогины, оболочки их в месте соприкосновения растворяются и ядро спермация перетекает в полость трихогины. Передвигаясь по ней, оно попадает в брюшную часть карпогона и там сливается с его ядром. После оплодотворения карпогон отделяется от трихогины специальной пробкой, и вскоре трихогина отмирает. Оплодотворенная яйцеклетка остается заключенной в оболочку карпогона и собственной оболочки не образует. Этот факт свидетельствует о вероятном отсутствии в эволюции багрянок подвижных женских половых клеток.
Зигота, не проходя периода покоя, вскоре начинает прорастать. Обычно у водорослей она развивается непосредственно в новые растения, принадлежащие уже к бесполому поколению, т. е. в спорофиты. У красных водорослей, в отличие от всех остальных, зигота, прежде чем дать начало спорофиту, претерпевает сложное развитие, в результате которого образуются особые споры, дающие начало спорофиту. Называют эти споры карпоспорами. Они представляют собой голые, лишенные жгутиков клетки, неподвижные или изредка обнаруживающие амебоидные движения. Перед началом прорастания они одеваются оболочкой. Клетки, в которых образуются карпоспоры, называют карпоспорангиями. В каждом карпоспорангии формируется по одной карпоспоре.
У бангиевых процесс превращения зиготы в карпоспоры весьма прост: после оплодотворения карпогон делится с образованием 4-32 карпоспор (рис. 161, 3).
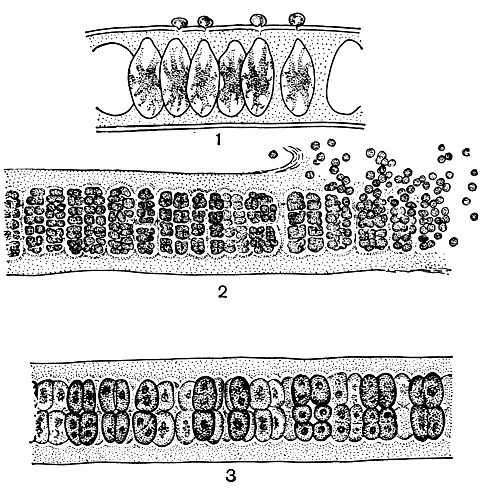
Рис. 161. Поперечный срез пластины Porphyra: 1 - с карпогонами; 2 - с сперматангиями; 3 - с карпоспорами
У флоридеевых развитие зиготы - сложный и многообразный процесс, в котором, кроме карпогона, участвуют многие специальные клетки. Этот процесс протекает неодинаково у разных багрянок, в каждой группе со своими особенностями и многочисленными деталями. Поэтому здесь целесообразно остановиться только на основных чертах развития зиготы.
После оплодотворения карпогона развиваются специальные нити, которые несут карпоспорангии. Эти нити состоят из небольшого числа клеток, называют их нитями гонимобласта. В наиболее простом случае - у водорослей из порядка немалиевых и гелидиевых - оплодотворенный карпогон отчленяет от себя несколько боковых клеток, которые, делясь, дают пучок нитей гонимобласта; конечные клетки этих нитей превращаются в карпоспорангии. Иными словами, применительно к этим водорослям можно говорить о непосредственном развитии нитей гонимобласта из карпогона (рис. 160, 2, 3).
У подавляющего большинства флоридеевых оплодотворенный карпогон вначале сливается с особыми клетками, богатыми питательными веществами, и только после этого развивается гонимобласт. Иногда после оплодотворения карпогон делится, и уже отчленившаяся от него клетка участвует в процессе слияния. Слияние может быть полным или частичным. Клетки, с которыми сливается карпогон, в литературе часто называют ауксиллярными. Но это не совсем верно. По мере того как изучались процессы развития зиготы, было установлено, что клетки, с которыми сливается карпогон, выполняют разную роль и занимают разное положение. Среди них следует различать два типа клеток: просто питающие клетки, которые образуются до или после оплодотворения и служат только для питания развивающегося гонимобласта, и настоящие ауксиллярные клетки. Последние также функционируют как питающие клетки, но их главная и чрезвычайно важная роль состоит в том, что они представляют собой исходную клетку, от которой начинается развитие гонимобласта. Иными словами, у тех багрянок, которые имеют ауксиллярные клетки, гонимобласт развивается не из карпогона, а из ауксиллярной клетки. Этому процессу обязательно предшествует слияние карпогона с ауксиллярной клеткой. Слияние осуществляется при помощи специальных нитей или выростов, которые отходят от карпогона и тянутся, пока не соединятся с ауксиллярной клеткой. Эти нити называют соединительными. Так как далеко не всегда ауксиллярные клетки располагаются вблизи карпогона, соединительные нити могут достигать значительной длины и состоять из большого числа клеток (рис. 176, 4). По этим нитям ядро оплодотворенного карпогона передвигается и переходит в ауксиллярную клетку; слияния ядер при этом не происходит. В этом состоит основное назначение соединительных нитей.
Ауксиллярные клетки отсутствуют в порядках немалиевых и гелидиевых, но имеются в остальных порядках класса флоридеевых. Ауксиллярные клетки характеризуются богатым содержимым и крупными размерами. Место и время их возникновения различны у раз-ных багрянок. Они могут развиваться на нитях корового слоя и от внутренних клеток слоевища, вблизи или в удалении от карпогона, до оплодотворения или после него.
Все эти особенности имеют большое значение для систематики флоридеевых. Можно предположить, что развитие системы ауксиллярных клеток и карпогонной ветви шло независимо в каждом из порядков, но везде связь ауксиллярной системы с карпогоном становилась все более тесной. У багрянок, уже обладающих ауксиллярными клетками, но стоящих еще на низкой ступени организации, они развиваются независимо от карпогонной ветви и бывают удалены от нее на значительное расстояние; образование их не приурочено к процессу оплодотворения: они возникают до него. У этих водорослей соединительные нити развиты очень сильно. По мере дальнейшего развития ауксиллярные клетки начинают возникать в непосредственной близости от карпогонной ветви и образуют с ней уже единую структуру. Соединительные нити заметно укорачиваются, редуцируются и в конечном итоге представляют собой лишь короткий неклеточный отросток карпогона. В тех случаях, когда ауксиллярные клетки развиваются вместе с карпогонной ветвью, образуя единую систему нитей, и оплодотворенный карпогон сливается со своей ауксиллярной клеткой, мы говорим о прокарпе. На ранних этапах развития прокарпа карпогон необязательно сливается с рядом расположенной ауксиллярной клеткой; соединительные нити могут простираться дальше и соединяться с ауксиллярными клетками соседних прокарпов.
На следующей ступени ауксиллярные клетки развиваются от той же клетки, что и карпогонные ветви, и только после оплодотворения карпогона.
Таким образом, можно выделить четыре типа настоящих ауксиллярных клеток:
1. Ауксиллярные клетки развиваются перед оплодотворением как отдельные клетки на специальных клеточных нитях - это тип криптонемиевых водорослей.
2. Ауксиллярными клетками служат обычные интеркалярные клетки слоевища, как правило, клетки внутренней коры, и образуются они перед оплодотворением; это тип гигартиновых.
3. Ауксиллярные клетки отчленяются перед оплодотворением от дочерней клетки несущей клетки карпогонной ветви; этот тип характерен для родимениевых.
4. Ауксиллярные клетки отчленяются от несущей клетки карпогонной ветви только после оплодотворения.
Просто питающие клетки поставляют питательные вещества из материнского растения благодаря полному или частичному слиянию с соединительными нитями или нитями развивающегося гонимобласта. Когда питающие клетки собраны в большом количестве и их собрания хорошо очерчены, можно говорить о питающей ткани (рис. 167, 5ж). Питающие клетки встречаются у всех флоридеевых, тогда как типичные ауксиллярные клетки характерны только для четырех порядков из шести.
Итак, у наиболее примитивных представителей класса флоридеевых развитие гонимобласта связано только с карпогонной ветвью. Затем возникают специальные питающие клетки, которые частично сливаются с развивающимися нитями гонимобласта и обеспечивают их питательными веществами. Позднее образуется ауксиллярная система, которая существует наряду с питающими клетками. Постепенно роль проводника питательных веществ полностью берет на себя ауксиллярная клетка и просто питающая ткань редуцируется. Аук-силлярная система также становится менее громоздкой и более тесно связанной по месту и времени возникновения с оплодотворенным карпогоном.
Развитие гонимобласта заключается в образовании системы клеточных нитей. Созревая, клетки нитей превращаются в карпоспоры. Различаются багрянки, у которых в карпоспоры превращаются конечные клетки нитей гонимобласта, и багрянки, у которых карпоспоры располагаются цепочкой, так как они образованы рядом клеток. У некоторых флоридеевых все клетки гонимобласта преобразуются в карпоспоры. Зрелый гонимобласт называют цистокарпом. Строго говоря, под цистокарпом следует понимать не только сам гонимобласт, но и окружающую его вегетативную ткань, когда она имеет определенное морфологическое и анатомическое строение и образует обертку гонимобласта. Обертка создается клетками коровых нитей или клетками, расположенными рядом с карпогонными ветвями.
Место возникновения цистокарпов, их форма, характер оболочки, способ выхода карпоспор - все эти признаки имеют важное значение для разграничения родов. Обычно цистокарпы располагаются в молодых частях растений - на ветвях последних порядков или участках близ вершины слоевища. У форм со свободно нитчатым строением они располагаются на поверхности ветвей в виде шаровидных скоплений клеток. Формы с плотным строением характеризуются погруженными или полупогруженными цистокарпами. Они имеют бугорчатую, шаровидную или кувшинообразную форму, и выступая над поверхностью слоевища, часто хорошо заметны даже невооруженным глазом. Для выхода карпоспор в обертке цистокарпа образуются специальные отверстия - одна или несколько пор. Однако пор может и не быть, тогда карпоспоры выходят после разрушения внешней оболочки цистокарпа.
Циклы развития. Как мы видели, красные водоросли размножаются половым и бесполым путем. Оба способа взаимосвязаны и обычно сменяют друг друга в ходе жизненного цикла багрянок. В подавляющем большинстве случаев органы полового и бесполого размножения расположены на разных растениях. Споры, служащие для бесполого размножения (моноспоры, тетраспоры, биспоры и т. п.), образуются на растениях-спорофитах, оогонии и сперматангии - на гаметофитах; при этом у большинства багрянок существуют мужские и женские гаметофиты. Иными словами, половая и бесполая формы развития представлены обычно самостоятельными свободноживущими поколениями. Однако по мере изучения циклов развития водорослей различных видов все чаще отмечаются случаи, когда одна форма развития возникает на другой, и тогда на одном и том же индивидууме можно найти и тетраспоры, и органы полового размножения. Механизмы этого явления пока остаются невыясненными.
В литературе о циклах развития водорослей, особенно в иностранной, гаметофит и спорофит часто называют поколениями. Это было бы верно, если бы половое поколение обязательно сменялось бесполым. Но такая строгая смена происходит далеко не всегда. Сплошь и рядом у разных водорослей и гаметофит, и спорофит могут воспроизводить себя в течение нескольких поколений. Поэтому, касаясь вопроса о циклах развития красных водорослей, мы будем говорить о смене форм развития, а не о смене поколений.
Вопрос о циклах развития красных водорослей - один из самых интересных и трудных в альгологии. Ученых интересует не только характер циклов развития сам по себе, его особенности у разных видов, но, главное, происхождение и эволюция этого явления у водорослей отдела Rhodophyta. Говоря о цикле развития, обычно имеют в виду два аспекта этого явления - морфологическую смену форм развития и цитологическую. Особенности цитологического цикла определяются местом редукционного деления и соотношением диплоидной и гаплоидной фаз. Часто у водорослей морфологические изменения происходят независимо от цитологических, но у красных водорослей эти явления, по-видимому, взаимосвязаны.
У багрянок существует несколько типов циклов развития. В подавляющем большинстве случаев происходит смена трех форм развития: тетраспорофита, гаметофита и карпоспорофита. При этом карпоспорофитом называют гонимобласт с карпоспорами, который принято считать самостоятельным поколением, ведущим паразитический образ жизни на женском гаметофите и потому морфологически редуцированным. В этом цикле имеются две свободноживущие формы развития - тетраспорофит и гаметофиты. Редукционное деление происходит при образовании тетраспор. Следовательно, тетраспоры и возникающие из них гаметофиты представляют собой гаплоидную фазу, тогда как зигота, карпоспорофит и тетраспорофит относятся к диплоидной фазе.
Такой цикл развития представлен у багрянок двумя разновидностями. Гаметофиты и тетраспорофит могут иметь совершенно одинаковое морфологическое строение. Этот тип был впервые открыт у полисифонии, поэтому его так называют - тип Polysiphonia. У других багрянок слоевище тетраспорофита редуцировано по сравнению с гаметофитом и сильно отличается от него по строению. Часто в таких случаях спорофит представлен стелющимися или корковидными растениями. Ярким примером подобного цикла с так называемой гетероморфной сменой форм развития служит цикл развития порфиры. У этой водоросли пластинчатые крупные гаметофиты, т. е. то, что мы обычно называем порфирой, сменяются микроскопическими стелющимися спорофитами, построенными из однорядных нитей.
Среди филлофоровых (порядок гигартиновых) наряду с видами, цикл развитиц которых проходит по типу полисифонии, есть виды с очень своеобразным укороченным циклом. На крупных свободноживущих растениях - гаметофитах после оплодотворения развиваются нематеции с тетраспорами, а карпоспорофит отсутствует. Этот цикл можно объяснить морфологической редукцией не только тетраспорофита, но и карпоспорофита. Последний редуцируется до зиготы, которая непосредственно превращается в карпоспору. Та прорастает как паразит тут же, не отделяясь от гаметофита, в тетраспорофит, тоже морфологически редуцированный. Он напоминает нити нематеция, собранные в виде подушек и шариков на поверхности слоевища. До тех пор пока не был изучен цикл развития этих водорослей, редуцированные тетраспорофиты принимали за самостоятельные организмы.
Редукция гаметофитов в цикле развития красных водорослей встречается гораздо реже. Обычно она выражается в недоразвитии гаметофита и прорастании его на тетраспорофите.
До последнего времени считали, что у красных водорослей имеются два принципиально отличных типа циклов развития. Кроме уже описанного, в котором участвуют три формы развития, а редукционное деление происходит при образовании тетраспор, существует другой, при котором тетраспорофит отсутствует, а имеется только одна свободноживущая форма развития - гаметофит с паразитирующим на нем карпоспорофитом. Полагали, что редукционное деление в этом цикле происходит сразу после оплодотворения, в самом начале прорастания зиготы, и, следовательно, весь цикл проходит в гаплоидной фазе. Однако недавние исследования показали, что единственное место, где происходит редукционное деление, - тетраспорангии, и поэтому в цикле развития всегда представлены две цитологические фазы - гаплоидный гаметофит и диплоидные карпоспорофит и тетраспорофит. Кроме того, оказалось, что у большинства видов, которые, как считалось, в природе были представлены только гаметофитом и карпоспорофитом, имеется и тетраспорофит. Эти тетраспорофиты - очень мелкие, сильно редуцированные растеньица с весьма отличающейся от гаметофита и карпоспорофита структурой, поэтому исследователи либо не замечали их, либо считали другими организмами.
Таким образом, в цикле развития всех флоридеевых происходит смена трех форм развития, представленных как в гаплоидной, так и в диплоидной фазе. Но, как мы видели, не всегда все эти формы развиты одинаково и являются свободноживущими организмами. Какой же тип цикла развития следует считать наиболее примитивным и исходным для всего существующего многообразия циклов развития багрянок? Большинство исследователей сходится на том, что первоначальный цикл развития заключался в смене трех морфологически одинаковых форм развития, которые существовали независимо одна от другой. Зигота покидала материнское растение и прорастала в свободноживущий карпоспорофит.
Следует сказать, что такого рода цикла развития, принимаемого учеными за исходный, в природе не существует. Это - гипотетический цикл, выведенный на основе логических умозаключений. Принимая за исходное состояние свободное существование морфологически раз-витого карпоспорофита, ученые считают, что позднее в процессе эволюции карпоспорофит перешел к паразитическому образу жизни на гаметофите, в результате чего возник цикл развития типа полисифонии. И наконец, дальнейшая эволюция привела к образованию гетероморфных циклов развития, когда гаметофит и тетраспорофит заметно различаются по строению и по степени морфологического развития. Можно ли безоговорочно согласиться с этими представлениями об эволюции циклов развития красных водорослей? Отсутствие гипотетического исходного цикла развития в природе даже у самых примитивных современных водорослей из класса флоридеевых заставляет усомниться в действительном его существова-нии в истории багрянок. Кроме того, ничего подобного нет в других отделах водорослей, где, как правило, зигота развивается в спорофит, и цикл развития состоит из смены двух форм - гаметофита и спорофита. Зигота багрянок не образует собственной оболочки и развивается в стенках карпогона. Не говорит ли это о том, что зигота никогда не покидала гаметофит?
Таким образом, гипотезу об эволюции циклов развития красных водорослей, принятую в настоящее время большинством специалистов, нельзя считать до конца обоснованной. Не исключена вероятность того, что карпоспорофит багрянок - более позднее эволюционное образование, и первоначальный цикл состоял из двух форм развития.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'