Порядок фукусовые (Fucales)
Слоевище у фукусовых кустистое, обладает верхушечным ростом посредством деления 1-8 особых верхушечных клеток. Образующиеся при их делении новые клетки растут и снова делятся. Клетки, составляющие осевую часть ветви, делятся слабо и вытягиваются в длину, образуя сердцевину. Проспоры, дающие гаметофиты, развиваются из клеток, отчленяемых верхушечной клеткой (или клетками) в сторону поверхностного ряда клеток слоевища. Обычно такие клетки вытягиваются в высоту и делятся поперечными перегородками. Клетка, превращающаяся в проспору, не удлиняется, наоборот, она округляется, приобретает яйцевидную или грушевидную форму, содержимое ее становится мелкозернистым и однородным. Проспора при развитии гаметофита делится вертикальной и горизонтальной перегородками (пельвеция равновершинная) или наклонными перегородками (аскофиллум узловатый). У проспор остальных фукусовых первая перегородка проходит горизонтально (рис. 147). Верхняя (язычковая) клетка получается меньше нижней. Язычковая клетка делится поперечными перегородками и развивается в волосок с базальной зоной роста. У одних фукусовых он развивается полностью (цистозейра), у других недоразвивается (capгассум).
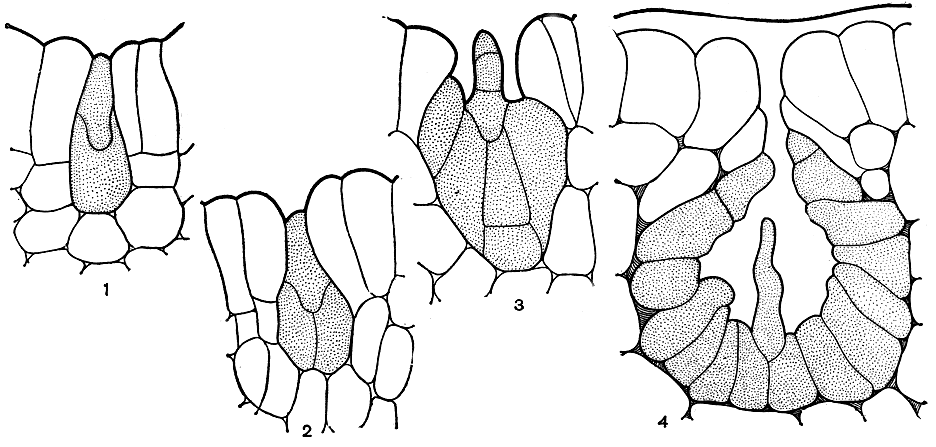
Рис. 147. Начальные стадии развития гаметофита у цистозейры (Cystoseira)
Интересно отметить, что с развития волоска начинается развитие проростков у ряда других бурых водорослей. Нижняя клетка, продукт деления проспоры, делится сначала вертикальной перегородкой. В последующем в новых клетках перегородки закладываются в вертикальной и горизонтальной плоскостях. Одновременно растут и делятся клетки окружающих тканей спорофита, в результате гаметофит оказывается погруженным в углубление. Гаметофиты фукусовых образуют вертикальные разветвленные и простые однорядные многоклеточные нити, часть из них несет гаметангии, а остальные служат парафизами. Ближе к краям у гаметофитов могут расти волоски с базальной зоной роста.
На слоевищах ряда фукусовых встречаются криптостомы и цекостомы. Криптостомы - это ямки, из которых растут многоклеточные волоски с базальной зоной роста. Цекостомы - микроскопические полости под поверхностью слоевища. Слой клеток, выстилающих полости криптостомов и цекостомов, развивается из таких же проспор, как и гаметофиты. Это стерильные гаметофиты, выполняющие физиологические функции. Криптостомы с волосками, видимо, служат для лучшего обмена с внешней средой, в частности для поглощения питательных солей. Криптостомы и цекостомы возникают тогда, когда не закладываются концептакулы, и поэтому они занимают разные участки на ветвях. На границе перехода одних образований в другие встречаются углубления, в которых развиваются гаметофиты промежуточного типа с недоразвитыми гаметангиями и большим количеством волосков наряду с парафизами. Гаметофиты у фукусовых бывают либо рассеяны на больших участках, ничем не отличающихся от стерильных, либо сосредоточены на вершинах побегов, становящихся из-за этого утолщенными и более широкими. Такие части слоевища называют рецептакулами.
Нормальное воспроизведение фукусовых возможно только половым путем, лишь немногие представители имеют стелющиеся ризомы или ризоиды, дающие на конце новые слоевища (например, саргассум Миябе). Антеридии образуются на гаметофитах на концах разветвленных веточек, и в их оболочке различимы два слоя. Антерозоиды выходят наружу в виде пакета, окруженного внутренней оболочкой. Субмикроскопическое строение антерозоидов не у всех фукусовых водорослей одинаково. Антерозоиды фукуса, аскофиллума и пельвеции имеют на переднем конце хоботок (рис. 148). Передний жгутик антерозоида химанталии снабжен на конце шипом, направленным вбок.
Оогонии у фукусовых сидят прямо на поверхности гаметофита или располагаются на одной поддерживающей клетке. Оогонии фукусовых снабжены трехслойной оболочкой, наружный слой называют экзохитоном, промежуточный - мезохитоном, внутренний - эндохитоном. Содержимое выходит в воду, окруженное двумя внутренними слоями. На этой стадии антерозоиды могут проникать в пакет яйцеклеток, но оплодотворения не происходит - этому препятствует поверхность яйцеклеток. Когда они полностью освобождаются от оболочек, происходит оплодотворение. Вслед за этим оплодотворенные яйцеклетки вырабатывают собственную толстую оболочку. У некоторых фукусовых наблюдается оплодотворение и прорастание яйцеклеток на материнском растении. Это бывает у тех представителей, у которых образуется по одной яйцеклетке в оогонии. В простейшем случае яйцеклетки прикрепляются слизью около отверстия концептакула (цистозейра), у других (Sargassaceae, Fucaceae) они прикрепляются слизистым стебельком, тянущимся от оогония и представляющим собой вытянутый мезохитон. Яйцеклетки остаются прикрепленными к материнскому растению 7-16 дней. За это время происходит их оплодотворение и дробление до стадии образования коротких первичных ризоидов, после чего проростки уносятся водой.
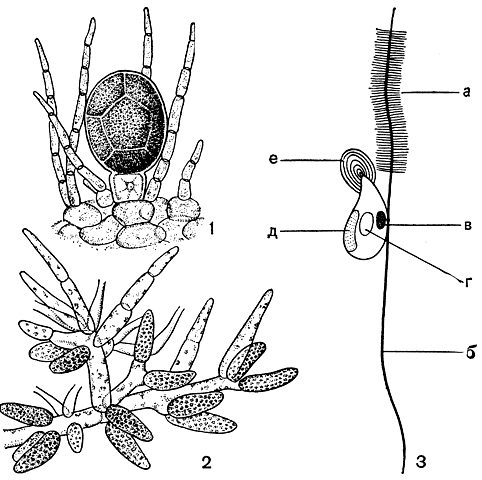
Рис. 148. Фукус пузырчатый (Fucus vesiculosus): 1 - оогоний; 2 - антеридии; 3 - схема строения антерозоида: а - передний жгутик, б - задний жгутик, в - глазок, г- ядро, д - хлоропласт, е - хоботок
Дробление яйцеклеток и развитие проростков у всех фукусовых протекает почти одинаково. После оплодотворения и появления обо-лочки яйцеклетка становится шаровидной, затем яйцевидной, ориентированной узким концом вниз. Первая перегородка проходит поперек и делит яйцеклетку на две примерно равные части (рис. 145, 1-3). В нижней части образуется вторая поперечная перегородка, отделяющая вниз небольшую клетку, называемую ризоидальной, которая дает со временем первичные ризоиды. Последующие перегородки, с третьей по шестую, проходят в вертикальных плоскостях в верхней и средней клетках. Из ризоидальной клетки вырастает от 1 до 32 ризоидов, в зависимости от рода и вида; предварительно эта клетка делится вертикальными перегородками по числу будущих ризоидов. Положение первой перегородки в проростке и направление роста ризоидов определяется, кроме света, соседством других яйцеклеток того же вида. При малом расстоянии между яйцеклетками направление их роста целиком определяется этим фактором, вызывающим рост вершин в противоположные стороны. Установлено, что взаимовлияние яйцеклеток определяется химическими веществами, выделяемыми ими в воду.
Представители порядка фукусовых отличаются большим разнообразием. Они растут во всех морях от Арктики до Антарктики, кроме Каспийского и Аральского морей.
Семейство хормозировые (Hormosiraceae)
Единственный представитель этого семейства - хормозира Банкса (Hormosira banksii) - имеет дихотомически-разветвленное слоевище, состоящее из шаровидных пузырей, соединенных короткими и узкими перетяжками (рис. 143, 3). Органом прикрепления служит диск. Рост у слоевища верхушечный, осуществляется посредством 4-8 клеток, которые располагаются в небольшом углублении на вершине каждой ветви. Концептакулы образуются в поверхностном слое пузырей. Гаметофиты обоеполые, в оогониях созревает по 4 яйцеклетки. Распространена хормозира у берегов Новой Зеландии, острова Тасмания и в нетропических водах Австралии, растет в литоральной зоне. В Новой Зеландии хормозиру используют в качестве корма, особенно для овец.
Семейство нотейевые (Notheiaceae)
Это семейство включает один род с одним видом. Нотейя аномальная (Notheia anomala) вырастает из концептакулов австралийских фукусовых хормозиры и ксифофоры, а также из концептакулов своих же особей. Высота слоевища достигает 12 см, побеги прямые, цилиндрические; боковые ветви вырастают со дна концептакулов. Рост верхушечный, посредством трех клеток. В концептакулах развиваются почти исключительно оогонии с 8 гаметами. Антеридии с 64 антерозоидами встречаются крайне редко. Одно время ставили под сомнение сам факт их существования. В 1960 г. Низамуддин и Уомерсли, изучая нотейю в культуре, обнаружили, что из оогониев, называемых ими макроспорангиями, выходят макроспоры с двумя жгутиками. Макроспоры прорастали в разветвленные нити с волосками, наподобие гаметофитов сперматохнуса (порядок хордариевые). На этих нитях развивались однорядные многогнездные спорангии, снова производящие макроспоры. Судьба этих спор осталась невыясненной. Зооиды, выходящие из антеридиев, не сливались с макроспорами и не прорастали. Низамуддин и Уомерсли перенесли нотейю в порядок хордариевых в качестве самостоятельного семейства.
Семейство химанталиевые (Himanthaliaceae)
К этому семейству относится один род с одним видом. Химанталия удлиненная (Himanthalia elongata) известна в странах Западной Европы как "ремневидная водоросль" или "водоросль-пуговица" (рис. 143, 4, 5). Ее слоевище состоит из воронковидного или дисковидного образования, прикрепленного эластичным стебельком с подошвой ко дну. На второй или третий год жизни из углубления в центре вырастает 1-4 дихотомически-разветвленных рецептакула длиной до 2-3 м и шириной 1-2 см. Рецептакулы обладают верхушечным ростом, который осуществляется посредством одной верхушечной клетки.
Химанталия интересна тем, что в оогониях развивается по одной очень крупной яйцеклетке (длиной до 0,5 мм), и тем, что после выхода гамет растение обычно отмирает целиком. В отличие от других фукусовых яйцеклетка химанталии при развитии в слоевище спорофита уплощается, становится вогнутой с нижней стороны и прикрепляется к грунту краями. Позднее с нижней стороны вырастают нитевидные ризоиды. В последующем развитие идет, как у других фукусовых.
Химанталия растет на камнях и скалах в нижнем горизонте литорали в умеренно прибойных местах в Атлантическом океане от берегов Южной Норвегии до северного побережья Марокко; наиболее обычна в Норвегии, Ирландии, Великобритании и Северной Франции.
Семейство фукусовые (Fucaceae)
Слоевище у фукусовых кустистое, прикрепляется к грунту диском. Рост ветвей осуществляется посредством одной верхушечной клетки, имеющей вид четырехгранной усеченной пирамиды. Сердцевина ветвей состоит из неплотно сомкнутых тонкостенных нитей и толстостенных гиф. Оогонии с 1-8 яйцеклетками. Ветви плоские или цилиндрические, без "листьев".
У водорослей рода пельвеция (Pelvetia) слоевище ветвится правильно или неправильно дихотомически. Ветви плоские или цилиндрические, без ребра. Воздушные пузыри одиночные. Криптостомы и цекостомы отсутствуют, изредка встречаются ямки без волосков. Рецептакулы расположены на вершинах ветвей. Оогонии содержат по две яйцеклетки.
Пельвеция желобчатая (P. canaliculata) имеет слоевище в виде кустов высотой до 15 см с дугообразными ветвями, согнутыми по продольной оси в виде желоба. Гаметангии созревают с конца июня по сентябрь. Растет пельвеция желобчатая над поясом фукусов в среднем и верхнем горизонтах литорали. Распространена она от Белого и Баренцева морей до севера Португалии.
Пельвеция Райта (P. wrightii) имеет слоевища длиной 10-90 см с прямыми ровными ветвями, несущими иногда одиночные пузыри. Выход гамет происходит в июле - сентябре. Произрастает в среднем и нижнем горизонтах литорали и в сублиторали до глубины 1 м в северной половине Японского моря, включая Хоккайдо и северную часть Хонсю, а также в заливе Анива и у Курильских островов до острова Уруп.
У представителей рода фукус (Fucus) слоевище дихотомически-разветвленное с плоскими ветвями, имеющими продольное ребро, а также криптостомы и цекостомы. Рецептакулы располагаются на вершинах ветвей. В оогониях образуется по 8 яйцеклеток. Виды рода фукус распространены в холодных и умеренных морях северного полушария. Часто они образуют большие заросли на литорали, это облегчает их сбор и использование. Виды рода фукус применяют в качестве удобрений, как корм для скота, для производства кормовой муки, альгинатов и других химических веществ. В морях СССР встречается 5 видов этого рода (табл. 18, 19).

Таблица 18. Бурые водоросли: вверху слева - фукус пузырчатый (Fucus vesiculosus); вверху справа - фукус двусторонний (F. distichus); внизу слева - бурая нитчатая водоросль хордария плетевидная (Chordaria flagelliformis) на фукусе пузырчатом; внизу справа - бурая нитчатая водоросль пилайелла литоральная (Pylaiella litoralis) на фукусе пузырчатом. Фото Ю. Астафьева

Таблица 19. Заросли бурых водорослей в начале отлива на литорали Белого моря. Фото В. Кузина
Фукус зубчатый (F. serratus) выделяется среди остальных видов ветвями с зубчатыми краями. Слоевища его раздельнополые, в длину достигают 1 м. В условиях Баренцева моря гаметы созревают в августе - сентябре, единичные рецептакулы со зрелыми гаметангиями встречаются на отдельных растениях почти круглый год, крайне редко их можно найти весной. Фукус зубчатый растет в нижнем и иногда в среднем горизонте литорали в верхней сублиторали. Встречается у берегов Европы от Новой Земли до севера Португалии и у берегов Северной Америки в районе залива Святого Лаврентия.
Слоевища фукуса пузырчатого (F. vesiculosa) часто имеют воздушные пузыри, обычно располагающиеся попарно по бокам ребра. В прибойных местах пузыри не развиваются. Ребро у ветвей резко выражено и прослеживается до вершины. Растения раздельнополые. Мужские экземпляры по сравнению с рядом растущими женскими растениями нередко имеют слоевища меньшего размера, с более узкими ветвями и более мелкими рецептакулами. Проспоры и гаметофиты появляются на спорофитах с сентября до конца января или до конца февраля, в сильно опресненных местах их закладка прекращается в ноябре. В январе на гаметофитах появляются первые гаметангии. В начале июня в гаметангиях образуются гаметы; выход их в воду продолжается до конца августа.
Фукус пузырчатый растет в разнообразных условиях - от сильно открытых до защищенных мест в среднем и нижнем горизонтах литорали и в верхней сублиторали, последнее чаще всего наблюдается в опресненных морях (Белое и Балтийское). Дерновинки мелких слоевищ, никогда не образующих проспор и гаметофитов, встречаются на илистых грунтах в верхнем горизонте литорали и в супралиторали вместе с моховидными и другими наземными растениями. Неприкрепленные слоевища фукуса пузырчатого встречаются в Белом море до глубины 22 м. Наиболее пышные заросли фукус пузырчатый образует на литорали в полузащищенных местах над поясом аскофиллума, но возможно и обратное их расположение. Фукус пузырчатый распространен у побережья от Карского моря до северо-запада Испании и у берегов Северной Америки от острова Элсмир до штата Северная Каролина (США). В Балтийском море он произрастает до восточного берега Выборгского залива.
Слоевища фукуса спирального (F. spiralis) внешне сходны со слоевищами фукуса пузырчатого, лишенными пузырей. Отличается этот вид тем, что у него слоевища обоеполые и рецептакулы имеют узкую стерильную кайму по краям. Растет фукус спиральный на литорали выше пояса фукуса пузырчатого. Распространен он у берегов Европы от Кольского полуострова до Канарских островов и у берегов Северной Америки от острова Ньюфаундленд до штата Нью-Йорк (США).
Два других вида - фукус двусторонний (F. distichus) и фукус исчезающий (F. evanescens) - отличаются от вышерассмотренных видов наличием цекостомов и менее четко выраженным ребром. Слоевища у них обоеполые. Фукус двусторонний распространен у берегов Азии и Европы от Карского моря до севера Великобритании и у берегов Америки от острова Элсмир до Нью-Йорка. Растет фукус двусторонний в местах с более сильным движением воды и глубже, чем фукус пузырчатый. Развитие органов размножения и созревание гамет происходят у фукуса двустороннего на две-три недели раньше.
Фукус исчезающий произрастает в Тихом океане от Приморского края, северной части Хонсю и севера Калифорнии до Чукотского и Восточно-Сибирского морей. Растет он в нижнем и среднем горизонтах литорали, в Арктике - в верхней сублиторали. Гаметы созревают летом.
Род аскофиллум (Ascophyllum) включает один вид - аскофиллум узловатый (A. nodosum). Слоевище его, достигающее 1-1,5 м в длину, имеет длинные редкодихотомическиветвящиеся ветви, на которых попеременно или супротивно расположены более короткие ветви. Рецептакулы развиваются как короткие булавовидные веточки по бокам крупных; при созревании гамет они раздуваются на вершине и становятся похожими на крупные ягоды. Слоевища у аскофиллума раздельнополые. Мужские экземпляры более темной окраски, с менее округлыми ветвями и более мелкими рецептакулами. Веточки-рецептакулы появляются на слоевищах с середины июля до конца октября, тогда же в них развиваются проспоры и гаметофиты. Первые гаметангии возникают в конце декабря, гаметы созревают в июне, и в течение месяца происходит их выход. Аскофиллум растет в нижнем и отчасти в среднем горизонте литорали и в верхней сублиторали (в Белом море). Распространен аскофиллум у берегов Азии и Европы от Карского моря до Португалии и у берегов Америки от Баффиновой Земли до штата Нью-Джерси (США).
Семейство цистозейровые (Cystoseiraceae)
Слоевище цистозейровых кустистое, прикрепляется к грунту диском или реже ризоидами. Рост ветвей идет посредством верхушечной клетки, имеющей вид трехгранной усеченной пирамиды. Сердцевина образована плотно сомкнутыми рядами клеток. В оогониях развивается по одной яйцеклетке. Ветви, в отличие от саргассовых, никогда не развиваются из пазух "листьев". "Листья" на ветвях имеются или отсутствуют.
У водорослей из рода цистозейра (Cyctoseira) воздушные пузыри однокамерные, без поперечных перегородок. Ветви цилиндрические или многогранные, с "листьями" или без них. В морях СССР виды рода цистозейра произрастают в Черном, Японском и Охотском морях и у Курильских островов. В Черном море растут цистозейра косматая (С. crinita, рис. 143, 7) и цистозейра бородатая (С. barbata, рис. 143, 6). Цистозейра косматая выделяется среди цистозейр, растущих в морях СССР, тем, что от диска, которым слоевище прикрепляется к грунту, отходит несколько вертикальных стволов (до 20). Длина стволов составляет 2/3 длины слоевища, достигающей 0,5-1,2 м. Пузыри одиночные, некоторые из них вильчато-рассеченные или с боковыми выростами. Рецептакулы обоеполые с выступающими бородавчатыми концептакулами и короткими шипиками по бокам. Развиваются рецептакулы на слоевище дважды в год - весной и осенью. Ветви, несущие рецептакулы, после выхода гамет разрушаются. Растет цистозейра косматая в открытых и полузащищенных местах на глубине до 10-15 м. Она распространена в Средиземном и Черном морях, кроме мелководной северо-западной части последнего.
У цистозейры бородатой от диска отходит, как правило, один высокий ствол, который у старых экземпляров становится шероховатым. Слоевище достигает в длину 1,7 м. Воздушные пузыри одиночные или собраны цепочками, располагаются они на конечных веточках или на верхних междоузлиях. Растения обоеполые; рецептакулы у них без шипиков, с гладкой, слегка волнистой поверхностью, развиваются дважды в год - весной и осенью. После выхода гамет ветви с рецептакулами разрушаются. Цистозейра бородатая растет в Атлантическом океане у южного берега Пиренейского полуострова, в Средиземном и Черном морях, иногда встречается в Азовском море у южного берега.
Цистозейра толстоногая (С. crassipes), произрастающая в дальневосточных морях СССР, имеет слоевища иного облика, чем предыдущие виды. Ствол у нее короткий и несет на верхнем конце короткие и толстые, иногда булавовидные, многолетние ветви, на которых вырастают длинные ветви неограниченного роста с короткими боковыми веточками. Слоевище в длину достигает 2-6 м. В нижней части слоевища бывают узкие "листья" без ребер. Воздушные пузыри одиночные или цепочками располагаются на конечных ветвях. Слоевища цистозейры толстоногой раздельнополые. Гаметангии созревают на юге ареала в июне, на севере - в августе.
Цистозейра толстоногая растет на камнях и скалах в нижнем горизонте литорали и в сублиторали на глубине до 25 м в открытых и полу-защищенных местах. Распространена она у берегов Азии от северо-восточного побережья Корейского полуострова до материкового побережья Охотского моря и от северо-восточцой части острова Хонсю до острова Уруп (Курильские острова). У побережья Северной Америки цистозейра толстоногая произрастает от южной части Берингова моря до штата Вашингтон (США).
Род хормофиза (Hormophysa) включает единственный вид хормофиза трехгранная (Н. triquetral широко распространенный в тропических морях. Его нередко относят к роду цистозейра. Отличие заключается в том, что у хормофизы ветви с широкими пластинчатыми краями, расходящимися по радиусам на три стороны.
Род халидрис (Halidrys) отличается от рода цистозейра стручковатыми воздушными пузырями, поделенными внутри поперечными перегородками на камеры, а также слегка уплощенными ветвями, расположенными большей частью в одной плоскости. У атлантических берегов Европы от севера Норвегии до Португалии распространен халидрис стручковатый (Н. siliquosa).
Семейство саргассовые (Sargassaceae)
У этих водорослей слоевище кустистое, прикрепляется к субстрату диском, реже ризоидами. Все выросты - ветви, воздушные пузыри и рецептакулы - растут из пазух "листьев", которые могут быть широкими с ребром или без него или шиловидными. Когда "листья" разрушаются, то в основании ветвей или других выростов остается короткий шип. Воздушные пузыри всегда одиночные и располагаются на конечных веточках. Сердцевина ветвей образована плотно сомкнутыми рядами клеток, только в старых побегах она разрушается и образуется полость с рыхлопереплетенными нитями. Ветви растут посредством верхушечной клетки, имеющей вид трехгранной усеченной пирамиды. Оогонии содержат по одной яйцеклетке. При созревании они выходят из концептакула, но остаются прикрепленными к оогонию слизистым стебельком. В таком положении происходит оплодотворение, дробление зиготы и развитие проростка до начала образования ризоидов, после чего они отделяются.
У водорослей из рода саргассум (Sargassum) "листья" пластинчатые с продольным ребром или без него или шиловидные (рис. 143, 8). Ствол короткий, от верхней его части отходят длинные ветви, несущие рецептакулы, эти ветви ежегодно отмирают. В тропических морях есть виды, у которых все слоевище однолетнее. Представители рода саргассум распространены в основном в тропических и субтропических морях, отдельные виды заходят в теплые воды морей умеренных поясов. Саргассум используют для получения альгинатов, отдельные виды, имеющие нежесткие ветви, употребляют в пищу. В СССР виды рода саргассум растут в дальневосточных морях. Наиболее часто встречаются несколько видов.
Саргассум бледный (S. pallidum) прикрепляется к грунту конусовидным диском, ствол у него грубый, с широко расставленными ветвями. Широкие "листья" с ребром; в основании слоевища они бывают овальными или удлиненными (длиной до 15 см). Выше по слоевищу "листья" становятся мельче и уже, вплоть до шиловидных. В условиях сильного движения воды крупные листья не развиваются. Воздушные пузыри шаровидные. Растения у этого вида раздельнополые. Выход гамет в заливе Петра Великого происходит с конца июня до начала августа, на севере Татарского пролива он продолжается до сентября. Длина слоевищ достигает 2-4,5 м. Этот вид растет в сублиторали на глубине до 20 м, иногда встречается в нижнем горизонте литорали.
Саргассум бледный распространен в Желтом и Японском морях, у Тихоокеанского побережья Японии, вдоль юго-восточного берега Сахалина и у южных Курильских островов до острова Итуруп.
Слоевище саргассума Миябе (S. miyabei) прикрепляется к грунту слегка расширенным основанием ствола и ризоидами, которые по всей своей длине прилегают к грунту. При неаккуратном сборе растений ризоиды легко обламываются и остаются на грунте; при этом кажется, что слоевища прикрепляются диском, как у других видов. Саргассум Миябе отличается еще коротким стволом и мелкими (длиной до 13 мм) клиновидными "листочками", густо покрывающими молодые побеги. Воздушные пузыри у саргассума Миябе эллипсоидные, с острым кончиком. Растения раздельнополые, рецептакулы мужских растений длиннее женских. Выход гамет у саргассума Миябе, растущего в заливе Петра Великого, происходит в июне - июле, на севере Татарского пролива он продолжается до октября. Растет саргассум Миябе в открытых и полузащищенных местах в нижнем горизонте литорали и в сублиторали. Этот вид встречается в северной половине Японского моря, включая остров Хоккайдо, у юго-восточного побережья Сахалина и южных Курильских островов до острова Итуруп.
Своеобразные виды рода саргассум живут в Саргассовом море. В основном там преобладают саргассум плавающий (S. fluitans) и саргассум погруженный (S. natans). Для обоих видов характерно отсутствие органов прикрепления, рецептакулов и какой-либо главной оси. Слоевища этих и других видов рода саргассум образуют спутанную массу, плавающую у поверхности.
Род турбинария (Turbinaria, рис. 143, 9) довольно обычен в тропических морях. Для его представителей характерны мясистые листья, расширяющиеся кнаружи в виде конуса. К грунту слоевище турбинарии прикрепляется маленьким диском и жесткими слабо разветвленными ризоидами. В прибойных местах молодые слоевища представлены слабо разветвленными цилиндрическими побегами и внешне не имеют ничего общего с более крупными слоевищами.
У берегов Японии и южной половины Японского моря, включая залив Петра Великого, у берега на малой глубине часто встречается представитель рода коккофора (Coccophora) - коккофора Лангсдорфа (С. langsdorfii). Она выделяется среди других водорослей прутовидными побегами высотой до 0,5 м, отходящими от короткого сильно разветвленного ствола с широким диском при основании. Весной и в начале лета побеги коккофоры покрыты крохотными чешуевидными треугольными "листьями".
На вершинах ветвей на коротких веточках расположены грушевидные пузыри, которые одновременно служат рецептакулами. Растения раздельнополые. Гаметы выходят в конце мая и в июне, после этого рецептакулы и несущие их веточки разрушаются, и слоевище преобразуется. Ребра крохотных чешуевидных "листьев" вырастают в нитевидные слабодихотомически-ветвящиеся веточки длиной до 4-15 см, отходящие перпендикулярно к ветвям. В связи с этим в конце лета и зимой длинные побеги коккофоры выглядят пушистыми.
По мере развития новых рецептакулов нитевидные веточки опадают. Яйцеклетки коккофоры используют в научных исследованиях для выяснения механизмов, определяющих развитие растений.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'