Порядок ламинариевые (Laminariales)
Ламинариевые имеют гетероморфный цикл развития с обязательным чередованием спорофитов и гаметофитов.
Зрелые спорофиты - крупные растения высотой 0,5-6 м, для отдельных видов обычны размеры в 20 м. Самые крупные ламинариевые достигают 50 м. Слоевище ламинариевых имеет одну или несколько пластин, расположенных на простом или разветвленном стволе. Слоевище прикрепляется к грунту диском или разветвленными ризоидами. Внутри пластины и ствола можно различить меристодерму и кору, состоящие из окрашенных клеток, промежуточный слой из крупных бесцветных клеток и сердцевину. У одних ламинариевых (подпорядок филлариевые) сердцевина состоит из плотно сомкнутых рядов цилиндрических нешироких клеток с оболочками обычной толщины и рядов широких клеток с толстыми боковыми стенками. Сердцевина других ламинариевых (подпорядок ламинариевые) образована неплотно расположенными клеточными нитями: одни из них идут поперек слоевища от клеток промежуточного слоя; другие ориентированы по длинной оси слоевища, причем клетки последних очень длинные и узкие и только у поперечных перегородок они воронкообразно расширяются (рис. 139, 1, 2). Эти нити называют трубчатыми нитями, а их поперечные перегородки - ситовидными пластинами, так как в них находятся поры. В краевой части сердцевины крупных ламинариевых, таких, как макроцистис и нереоцистис, среди тонких нитей располагаются ситовидные трубки. Они выделяются клетками большого диаметра по всей длине, их поперечными перегородками служат ситовидные пластины. В коре или промежуточном слое многих ламинариевых расположены слизистые каналы, образующие в слоевище сеть, петли которой вытянуты по его продольной оси. В некоторых участках слизистых каналов, особенно в местах их ветвления, располагаются группы секреторных клеток (рис. 140), содержащих большое количество диктиосом. Роль секреторных клеток и слизистых каналов ламинариевых пока не ясна; видимо, они выполняют какую-то физиологическую функцию. Количество их в слоевищах одного вида изменяется в зависимости от условий произрастания, и иногда они могут совершенно не развиваться.
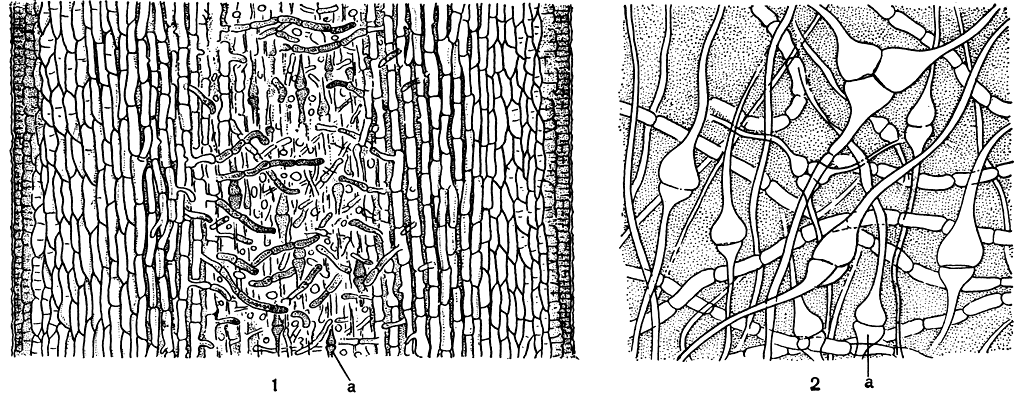
Рис. 139. Продольный разрез молодого слоевища ламинарии (Laminaria): 1 - при малом увеличении; 2 - при большом увеличении: а - трубчатые нити
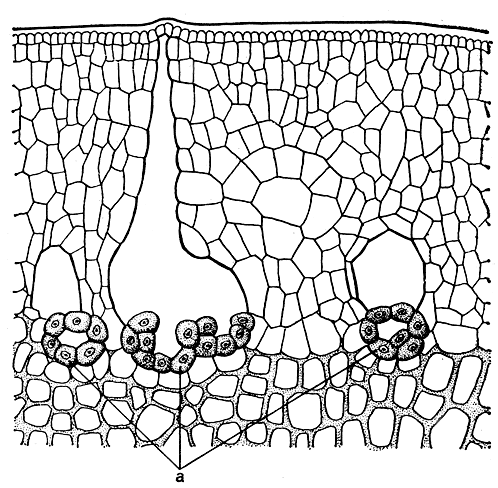
Рис. 140. Слизистые каналы ламинарии северной (Laminaria hyperborea): а - секреторные клетки
Для представителей семейства алариевых характерно присутствие в поверхностном слое пластин крупных клеток, называемых гландулярными; они наполнены фукозаном. В молодых частях слоевища эти клетки желтые, в старых частях они темно-бурые или черные.
Рост слоевищ спорофитов осуществляется посредством интеркалярной зоны роста. Она занимает основание пластины и вершину ствола. Если ствол разветвленный и имеется несколько пластинок, то интеркалярная зона роста находится в основании каждой пластины. Интеркалярная зона роста не отграничена резко от остальной части слоевища. Деление клеток и соответственно рост наблюдается и за ее пределами.
Наряду с процессами роста в пластинах спорофита ламинариевых происходят процессы разрушения верхнего конца. Нанося на пластину через определенные расстояния отверстия и измеряя, как эти расстояния увеличиваются со временем, установили, что за время роста пластина обновляется по длине 1-5 раз, а по массе до 20 раз. Слоевища ламинариевых обладают наибольшей скоростью роста, достигающей более 10 см в сутки. У нереоцистиса Лютке наблюдалось удлинение ствола на 13,5 см в сутки, не считая роста пластин. Особенно быстро слоевища ламинариевых растут в конце зимы и весной, когда температура воды нередко держится около 0°С. В период бурного роста пластина содержит много воды и поэтому, если смять ее рукой, легко трескается. С повышением температуры воды скорость роста замедляется и усиливаются процессы синтеза и накопления органических веществ. Пластины становятся толще, темнее и перестают трескаться при сминании. Наиболее интенсивно накопление органических веществ происходит в основании пластины. В то же время наибольшая скорость фотосинтеза отмечается в верхней ее части. Наличие сердцевины с хорошо развитыми проводящими элементами обеспечивает транспортировку веществ в пределах слоевища.
Большинство ламинариевых - многолетние растения. Ежегодно после завершения выхода спор пластинчатая часть слоевища целиком разрушается. Этот процесс начинается с вершины. Тогда же или несколько позднее в интеркалярной зоне роста начинается интенсивное деление клеток и происходит закладка новой пластины. Она появляется между стволом и старой пластиной. У одних ламинариевых между двумя пластинами хорошо выражен пережим, у других он отсутствует. Обычно новая пластина шире старой. По мере роста новой пластины старая отодвигается все дальше вверх, иногда ее остатки могут долго сохраняться на вершине. При низкой температуре воды разрушение старой пластины происходит медленно. В Арктике у ламинарии копытной (Laminaria solidungula) на вершине новой пластины можно наблюдать цепочку из трех-четырех старых пластин. Как свидетельство многолетнего роста некоторых ламинариевых, в их стволах заметны кольца. Лучше всего они выражены в самом низу - в наиболее старой части ствола.
Спорофиты всех ламинариевых образуют органы бесполого размножения одного типа - одноклеточные одногнездные спорангии, которые развиваются на поверхности слоевища среди одноклеточных булавовидных парафиз, содержащих в верхнем, расширенном конце хлоропласты и физоды (рис. 141). Лишь когда парафизы достигнут почти полного развития, начинают расти спорангии. У всех ламинариевых они имеют сходную форму - эллиптическую или цилиндрическую. Число зооспор в спорангиях колеблется в зависимости от вида и отчасти от внешних условий от 16 до 128. Образование зооспор сопровождается мейозом. При развитии спорангиев на пластинчатых частях слоевища появляются большие по площади пятна (или одно пятно), слегка (на доли миллиметра) поднимающиеся над поверхностью и незначительно выделяющиеся окраской. После созревания и выхода зооспор происходит разрушение спороносных участков пластин, на их месте образуются отверстия, а вскоре разрушается и вся пластина. Массовое развитие спорангиев у ламинариевых начинается в конце лета - начале осени. В холодных морях они сохраняются на слоевищах до начала следующего лета. На единичных экземплярах спорангии могут появляться в начале лета, а также в иные времена года.
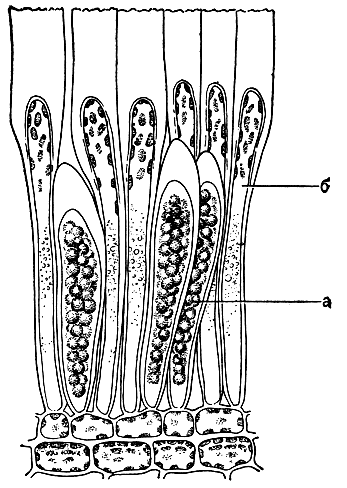
Рис. 141. Спорангии (а) и парафизы (б) ламинариевых водорослей
Зооспоры ламинариевых имеют типичное для подвижных клеток бурых водорослей строение. Они грушевидные, длиной 5-10 мкм, с двумя жгутиками, отходящими сбоку. Передний жгутик перистый, задний - бичевидный. Отчетливо видимая оболочка у зооспор отсутствует. Внутри, как правило, содержится по одному хлоропласту. Относительно существования глазка у зооспор ламинариевых имеются противоречивые сведения. При наиболее подходящих температурных условиях зооспоры способны к активному движению в течение двух суток. При высокой температуре (для ламинариевых морей СССР это +20°С и выше) они проявляют активность всего несколько минут или вообще не способны двигаться. Если за время активного движения зооспора не найдет место для прикрепления, она погибает. Направление движения зооспор определяется интенсивностью света. При высокой освещенности зооспоры проявляют отрицательный фототаксис. Встретив твердую поверхность, зооспора прикрепляется передним жгутиком, который сокращается и подтягивает тело зооспоры к субстрату. После этого зооспора теряет жгутики и округляется, у нее появляется ясно видимая оболочка, и зооспора превращается в эмбриоспору, которая прорастает без периода покоя.
В течение 1-2 суток эмбриоспора дает вырост - проростковую трубку, в которую переходит все содержимое. Направление роста проростковой трубки определяется светом и расстоянием от других эмбриоспор. При небольшом расстоянии между эмбриоспорами их проростковые трубки направлены в разные стороны. Протоплазма, переместившаяся в проростковую трубку, отделяется от опустевшей эмбриоспоры поперечной перегородкой (рис. 142). Так образуется первая клетка гаметофита. Оболочка эмбриоспоры долго сохраняется у гаметофита, часто даже тогда, когда на нем появляется спорофит. Гаметофиты большинства ламинариевых, за редким исключением, раздельнополые и различаются морфологически. Различия между мужскими и женскими гаметофитами заметны уже на стадии первой клетки. У женского гаметофита она короче и шире. В каждом спорангии ламинариевых образуется равное количество спор, дающих мужские и женские гаметофиты. Повышенная температура воды в первую очередь вызывает гибель женских спор.
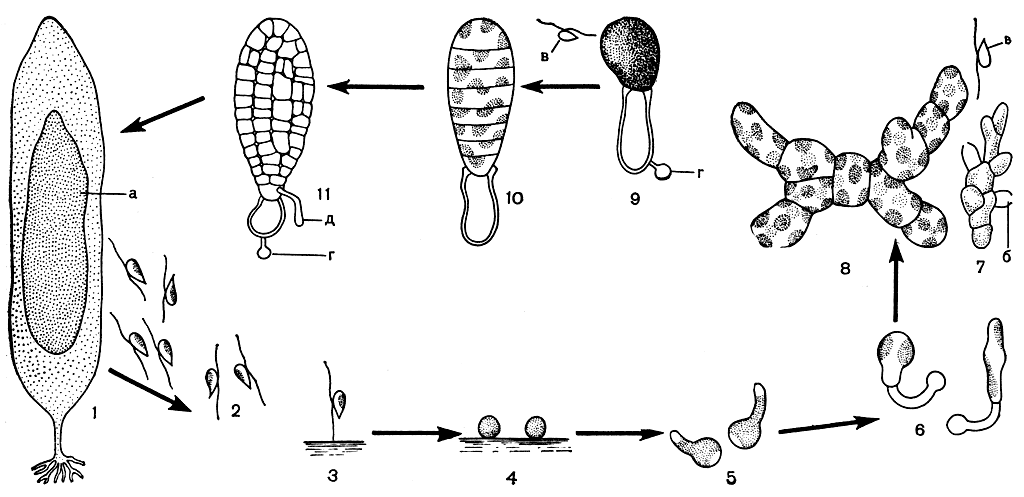
Рис. 142. Цикл развития ламинариевых водорослей: 1 - слоевище со спорами: а - пятно спорангиев; 2 - плавающие зооспоры; 3 - зооспора прикрепилась передним жгутиком к грунту; 4 - эмбриоспоры; 5 - эмбриоспоры с проростковой трубкой; 6 - образование первых клеток гаметофитов (слева - женский, справа - мужской); 7 - мужской гаметофит: б - антеридий, в - антерозоиды; 8 - многоклеточный женский гаметофит; 9 - одноклеточный женский гаметофит после выхода яйцеклетки из оогония: г - оболочка эмбриоспоры; 10-11 - начало развития слоевища спорофита: д - первичный ризоид
Гаметофиты ламинариевых многоклеточные, в виде однорядных разветвленных стелющихся нитей; женские гаметофиты могут быть одноклеточными. Мужские гаметофиты сильнее разветвлены и мельче женских, с более мелкими бледно окрашенными клетками. Обычно гаметофиты существуют не более одного - четырех месяцев до созревания гаметангиев. Женские многоклеточные гаметофиты не превышают в диаметре 300 мкм. При температуре, слишком высокой для образования гаметангиев, гаметофиты могут жить больше года и достигать в диаметре 1 см и более. В экспериментальных условиях гаметофиты ламинариевых способны к вегетативному размножению. Если разрезать многоклеточный гаметофит на части, из них вырастут новые гаметофиты. Это используется при всякого рода исследованиях, когда необходим генетически однородный материал. В природных условиях такое размножение едва ли возможно.
Половой процесс у ламинариевых - оогамия. Антеридии имеют вид мелких бесцветных клеток, располагающихся группами или по одной на поверхности вегетативных клеток. В антеридиях образуется по одному антерозоиду. После выхода антерозоидов мужские гаметофиты погибают. У многоклеточных женских гаметофитов в оогонии превращаются большей частью конечные клетки ветвей, иногда и предыдущая клетка. Таким образом, один гаметофит несет несколько оогониев. В случае одноклеточных женских гаметофитов единственная клетка со временем превращается в единственный оогоний. Образование оогониев сопровождается увеличением размеров клеток, содержимое их окрашивается в темно-бурый цвет. В каждом оогонии формируется по одной яйцеклетке. При созревании она выходит через отверстие на вершине оогония, но не отделяется, а остается прикрепленной к краям отверстия. В таком положении происходит ее оплодотворение и рост спорофита. После выхода всех яйцеклеток гаметофит отмирает. Однако оболочки его клеток еще долго сохраняются в основании спорофитов. На месте одного многоклеточного женского гаметофита развивается несколько спорофитов, которые располагаются по его периферии. Женские гаметофиты не предоставляют никаких питательных веществ развивающимся спорофитам, они обеспечивают им только место для прикрепления на грунте. Это имеет важное значение для существования водорослей, поскольку твердые грунты занимают в море небольшую площадь, и в довершении всего их поверхность в значительной мере занята другими водорослями, а также прикрепленными животными. Основная масса зачатков морских донных водорослей (спор и гамет) погибает из-за того, что они не попадают на подходящую поверхность.
Оплодотворенная яйцеклетка, оставаясь прикрепленной к гаметофиту, вырабатывает оболочку и без периода покоя трогается в рост. Вначале образуется однорядная нить примерно из семи клеток, затем наряду с поперечными перегородками появляются продольные и образуется однослойная пластинка. Из нижних клеток пластинки вырастают тонкие ризоиды. По мере роста пластинки в ней увеличивается число слоев клеток, происходит их специализация, появляются ствол и многорядные ризоиды. Если яйцеклетка почему-либо оказывается не прикрепленной к оболочке оогония, то оплодотворение происходит, но развитие спорофита идет ненормально. Образуется шарообразное скопление клеток, которое существует около месяца и погибает.
В лабораторных условиях удавалось выращивать гаметофиты апоспорически, минуя зооспоры. При содержании стерильных спорофитов в аквариуме в течение трех-четырех месяцев у них отмирали клетки. Несколько клеток оставалось живыми, и они развивались в однорядные разветвленные нити, на которых образовывались оогонии и антеридии. Поскольку клетки спорофитов диплоидные, то и гаметофиты получались диплоидными. Яйцеклетки таких гаметофитов после оплодотворения развивались в тетраплоидные спорофиты. Таким же путем удавалось выращивать гаметофиты из клеток гаплоидных спорофитов, полученных посредством партеногенеза (из неоплодотворенных яйцеклеток) и апогамии (из вегетативных клеток гаметофитов). Партеногенетическому развитию спорофитов предшествует массовая дегенерация яйцеклеток после выхода их из оогониев в условиях отсутствия мужских гаметофитов. В нормальные спорофиты при этом партеногенетически развивались немногие яйцеклетки. Апогамные спорофиты вырастают из конечных клеток зрелых ветвей мужских гаметофитов.
Ламинариевые обычны для холодных вод северного и южного полушарий, где даже в самые теплые месяцы температура воды держится выше +20°C всего лишь в течение двух-трех недель. Причем в северном полушарии растет большее число родов и видов. В последнее время ламинариевые (виды рода ламинария) обнаружены в тропических водах около Бразилии и в Филиппинском море, но растут они там на глубине 70-85 м, где температура воды ниже, чем у поверхности.
Ламинариевые встречаются по вертикали от поверхности моря до глубины 200 м (Laminaria rodriguezii в Средиземном море). В литоральной зоне они растут в основном в нижнем горизонте литорали в тех местах, где во время отлива подвергаются действию брызг, или там, где постоянно преобладают пасмурная погода и высокая влажность воздуха. Так, ламинариевые занимают большие площади на литорали Командорских островов, отличающихся как раз такими погодными условиями.
Таким крупным водорослям, как ламинариевые, нужны условия, при которых обеспечивалось бы интенсивное поступление питательных веществ к слоевищам. Этому соответствуют места с постоянным сильным течением или прибоем. Там, где нет достаточных по силе течений или прибоя, ламинариевые растут в водах, обогащенных питательными веществами (но не до степени загрязнения) за счет разного рода стоков, содержащих много органики, или за счет неглубокого залегания илистых грунтов. Поэтому заросли ламинариевых встречаются у открытых берегов, особенно у мысов, далеко вдающихся в море. Заросли же видов, способных расти в спокойных местах, располагаются по соседству с илистыми грунтами. Это приводит к тому, что ламинариевые можно найти на глубине, в условиях пониженной освещенности, но при наличии ила среди камней, а в том же месте у берега, на меньшей глубине, их нет,- если там отсутствует постоянный прибой, а дно имеет большой уклон, из-за чего органические остатки опускаются глубже. Точно так же заросли ламинариевых легче встретить в местах с мутной водой, подверженной интенсивному движению, чем в местах с прозрачной водой, находящейся в менее активном движении.
Подпорядок филлариевые (Phyllarineae)
Сердцевина у филлариевых плотного строения; она состоит из нитей большого диаметра с толстостенными клетками и из нитей меньшего диаметра с тонкостенными клетками; трубчатые нити, ситовидные трубки и гифы у них отсутствуют. Слизистые каналы неизвестны. К подпорядку относится одно семейство филлариевых (Phyllariaceae) с двумя родами: филлария (Phyllaria) и саккориза (Saccorhiza), распространенными в северной части Атлантического океана. Слоевище филлариевых имеет одну целую или рассеченную на вертикальные лопасти пластину, переходящую в ствол с органами прикрепления при основании. У филларии органами прикрепления служат ризоиды. Молодые слоевища саккоризы прикрепляются к субстрату диском; позднее в основании ствола на некоторой высоте над грунтом образуется диск (саккориза кожистая - S. dermatodea) или луковицеобразное расширение (саккориза многополосная - S. polyschides), с нижней стороны которых вырастают неразветвленные ризоиды.
В морях СССР растет один вид - саккориза кожистая. Внешне она напоминает ламинарию. Поверхность пластины выглядит бархатистой, так как она покрыта волосками. Слоевище у саккоризы кожистой однолетнее, достигает в длину 1-2 м. Спорангии развиваются в октябре-декабре в основании пластины и на стволе. Растет этот вид в верхней сублиторали и изредка в нижней литорали в открытых местах с пологим уклоном дна, а также в полузащищенных местах. Саккориза кожистая встречается у берегов Северной Атлантики от южной части Карского моря до севера Норвегии и у берегов Америки от острова Элсмир до севера штата Массачусетс (США).
Саккориза многополосная растет у берегов Западной Европы. Она интересна тем, что перед развитием спорангиев у нее по бокам ствола вырастает складчатая бахрома, на которой образуются спорангии. Кроме этого, спорангии могут располагаться на луковицеобразном расширении ствола и в нижней части пластины.
Подпорядок ламинариевые (Laminarineae)
Сердцевина неплотного строения с гифами, трубчатыми нитями или с ситовидными трубками. Слизистые каналы имеются или отсутствуют.
Семейство хордовые (Chordaceae)
Слоевище у хордовых однолетнее, шнуровидное, неразветвленное, с полостью, слизистые каналы отсутствуют. К субстрату прикрепляется диском. Зона роста расположена под вершиной. Семейство включает один род хорда (Chorda), содержащий всего 2 вида.
Хорда нитевидная (Chorda filum) имеет оливково-желтое слоевище, слабо покрытое волосками, часто бесцветными. Нижняя клетка волосков почти в два раза уже нижележащей клетки слоевища. Растет в лужах нижнего горизонта литорали и в сублиторали в полуоткрытых и защищенных местах на скалах и заиленном песке на глубине до 15-20 м. Хорда нитевидная встречается в северной половине Атлантического и Тихого океанов, южная граница ее распространения совпадает с зимней изотермой воды +12 °С.
Слоевище хорды пушистой (С. tomentosa) обычно оливково-зеленого цвета, густо покрыто интенсивно окрашенными волосками. Нижняя клетка волосков слабо отличается по ширине от нижележащей клетки слоевища. Хорда пушистая растет в лужах нижнего горизонта литорали и в верхней сублиторали до глубины 5 м в полуоткрытых и защищенных местах. Это растение весенней флоры, южная граница его распространения проходит значительно севернее, чем у хорды нитевидной.
Семейство ламинариевые (Laminariaceae)
У представителей этого семейства на вершине ствола расположена одна пластина, реже несколько, на боковых его поверхностях особые пластинки отсутствуют. У многолетних видов пластинка следующего года возникает интеркалярно. Продольные разрывы пластин никогда не возникают интеркалярно на стыке пластины и ствола, разрывы идут от верхнего края вниз.
Род ламинария (Laminaria) насчитывает 30 видов, причем только 3 из них растут в южном полушарии: 2 вида у берегов Бразилии на большой глубине (70-80 м), 1 вид у южной оконечности Африки. Остальные виды распространены в северном полушарии; больше всего видов ламинарий в Тихом океане. Пластина у этих водорослей ровная или морщинистая, без волосков, целая или рассеченная, без отверстий, иногда с двумя продольными рядами вмятин и выпуклостей или с двумя продольными складками. Органами прикрепления служат диск, ризоиды или ризом (стелющийся побег с ризоидами). Наибольшее промысловое значение имеют ламинария сахаристая, ламинария пальчатая, ламинария северная, ламинария японская, ламинария узкая.
У ламинарии сахаристой (L. saccharina) нерассеченная удлиненная пластина, ровная или морщинистая, с темной продольной полосой, иногда с двумя продольными рядами вмятин и выпуклостей. Слоевище прикрепляется к субстрату ризоидами. Слизистые каналы у слоевищ, растущих в морях СССР, известны только в пластине, а у растений атлантического прибрежья Северной Америки они бывают и в пластине и в стволе. Спорангии начинают развиваться одновременно на обеих поверхностях пластины, при этом очертания пятен спорангиев в точности совпадают. Зрелые спорангии наблюдаются с конца августа по январь, а иногда до июня. Слоевище у этой водоросли многолетнее, максимальная его длина достигает 7 м.
Ламинария сахаристая растет в сублиторали на глубине до 20 м, изредка встречается в нижнем горизонте литорали, особенно в литоральных "ваннах". Это один из наиболее массовых видов рода ламинария в северной части Атлантического океана. Ламинария сахаристая распространена от западной части моря Лаптевых и Баффиновой Земли на севере до севера Португалии и штата Массачусетс (США) на юге.
В восточном секторе Арктики и в северной части Тихого океана (до северных островов Японии на юге) широко распространена ламинария Гурьяновой (L. gurjanovae), которую прежде тоже считали ламинарией сахаристой. В дальневосточных морях ламинарией сахаристой по ошибке еще называют растения ламинарии курчавой (L. cichorioides), слоевища которой в зависимости от скорости движения воды могут иметь различный облик. В спокойных местах пластина с сильно курчавыми краями и двумя рядами крупных выпуклостей и вмятин. В местах же с постоянным движением воды пластины гладкие и с ровными краями. Такие слоевища и считают ошибочно ламинарией сахаристой. Ламинария курчавая отличается от ламинарии сахаристой и ламинарии Гурьяновой тем, что спорангии у нее начинают развиваться сперва на одной поверхности пластины и лишь с большим запозданием на второй. Поэтому очертания пятен спорангиев на них не совпадают. Имеются и другие отличительные признаки. Распространена ламинария курчавая в северной половине Японского моря, вокруг острова Хоккайдо, около острова Кунашир и островов Малой Курильской гряды, вдоль южного и восточного берегов острова Сахалин до залива Терпения, крайне редко она встречается у острова Итуруп и в северо-западной части Охотского моря.
У ламинарии японской (L. japonica) пластина нерассеченная, удлиненная (длина ее 2-6 м, иногда до 12 м, а ширина 10-35 см), расположенная на стволе несколько косо. Орган прикрепления - ризоиды (табл. 39,1). По продольной оси пластины проходит широкая и толстая срединная полоса, занимающая 1/5-1/2 ее ширины и ограниченная по краям двумя продольными складками. Молодые слоевища часто имеют два ряда выпуклостей и вмятин на месте продольных складок. Слизистые каналы в пластине расположены между сердцевиной и корой, часто в два ряда, особенно в срединной полосе. Сердцевина в пластине уже, чем у других видов. У молодых слоевищ ствол цилиндрический, у немолодых - уплощенный или желобчатый. Пластина плавно переходит в ствол. Слизистые каналы в стволе всегда имеются.
Слоевища ламинарии японской растут два года. На южной границе распространения, где температура воды в море летом бывает выше + 20 °С, они однолетние. Спорангии у этого вида образуются на одной или двух поверхностях пластины; в последнем случае они сначала развиваются на нижней, слегка вогнутой поверхности, и очертания групп спорангиев на двух сторонах пластины не совпадают. В условиях естественных зарослей спорангии начинают появляться с конца июля, крайне редко спороносящие экземпляры встречаются в июне.
Ламинария японская считается самым ценным промысловым видом среди бурых водорослей. Она обладает наилучшими вкусовыми качествами. Ее пластина толще и менее жесткая, чем у других видов рода ламинария. Наибольшую пищевую ценность имеет утолщенная срединная полоса слоевищ второго года. Первогодние слоевища считаются непромысловыми, хотя они и достигают крупных размеров: пластина у них узкая, тонкая и содержит меньше питательных веществ. Ламинарию японскую используют для приготовления первых, вторых и третьих блюд, а также разнообразных консервов и кондитерских изделий. Слоевища или их части, непригодные для пищевого использования, идут в химическую переработку для получения альгинатов, маннита, йода и т. д.
Ламинария японская растет в местах с постоянным движением воды, она встречается от поверхности до глубины 25 м, реже 35 м. Промысловые заросли находятся в основном на глубине 4-10 м. Этот вид распространен в северной половине Японского моря - от побережья КНДР и северной части острова Хонсю до 50 ° с. ш., а также у южного и юго-восточного берега Сахалина и у южных Курильских островов. Заросли у некоторых островов Малой Курильской гряды дают наибольшую биомассу - 140 кг/м2.
В 20-х годах нашего столетия зачатки ламинарии японской были случайно завезены в Желтое море к побережьям Ляодунского и Шаньдунского полуостровов. В КНР были разработаны методы ее искусственного разведения, и сейчас ее культивируют вдоль побережья КНР вплоть до Кантона.
В естественных условиях запасы ламинарии японской подвержены сильным колебаниям. Это отражается на объеме заготовок. Большой урон зарослям наносят сильные штормы и торошение льда у берега в холодные зимы; освобождаемые во время промысла участки дна нередко заселяются непромысловыми водорослями. Поэтому люди издавна пытались искусственно увеличить площади под ламинарией японской. Было найдено несколько способов, с успехом применяемых в Японии, КНР и КНДР. Эти способы используются и для выращивания других видов рода ламинария и прочих водорослей. Самый простой из них - это чистка твердого грунта от других водорослей и животных в период, когда ожидается появление в воде спор ламинарии японской, и последующий уход за зарослями, т. е. удаление сорных растений и растительноядных животных. Для расширения площади под зарослями в море с лодок сбрасывают камни или там, где возможно, взрывают скалы на берегу, обрушивая их в воду. На подготовленный таким образом новый грунт или на очищенный старый для ускорения появления зарослей сбрасывают зрелые слоевища, собранные в других местах. Остальные способы выращивания ламинарии японской связаны с погружением в море субстратов, искусственно засеянных спорами. В качестве таких субстратов используют камни и бетонные блоки, сбрасываемые на дно, или веревки, которые подвешивают к канатам, поддерживаемым у поверхности поплавками и закрепленным на месте якорями.
Посев спор ламинарии японской и других водорослей на субстраты делают следующим образом. Чтобы выход спор был дружным, предварительно производят стимулирование слоевищ. Для этого их выкладывают на воздух в затененном месте на несколько часов или на ночь. Для контроля за ходом стимулирования периодически на поверхность слоевища наносят каплю морской воды, через несколько минут эту каплю переносят на предметное стекло под микроскоп. Когда в поле зрения (при увеличении объектива 8х, окуляра - 10х) будет 8-10 спор, слоевище считается стимулированным. Его кладут в бассейн, куда предварительно поместили субстраты для последующего культивирования ламинарии. Засеянные спорами камни сбрасывают в море в подходящих местах. При культивировании ламинарии на веревках иногда предварительно выращивают рассаду, которую позднее рассаживают с нужными промежутками на другие веревки. Выращивание ламинарии на веревках дает наибольший урожай с единицы площади, и сейчас оно стало наиболее распространенным на плантациях. Для получения максимальных урожаев плантации ламинарии японской размещают вблизи городских стоков или вносят в воду минеральные удобрения. Урожай ламинарии японской при таком культивировании достигает 50-65 т сухой массы с гектара.
Ламинария узкая (L. angustata) имеет нерассеченную длинную узкую пластину, по продольной оси которой проходит узкая темная полоса, прогнутая в виде мелкого желоба. Вся пластина, кроме основания, тоже имеет вид желоба. Орган прикрепления - ризоиды. Слоевища ла-минарии узкой достигают наибольшей длины по сравнению с другими видами рода ламинария, известны экземпляры длиной до 22 м. Продолжительность жизни этой водоросли 3-4 года. Спорангии появляются в конце июля и располагаются на одной или двух поверхностях пластины, причем очертания групп спорангиев на них не совпадают. Ламинария узкая растет в нижнем горизонте литорали и в сублиторали до глубины 8-12 м в местах с постоянным прибоем или течением. Распространение этого вида ограничено островом Кунашир, островами Малой Курильской гряды, островом Хоккайдо и северо-восточной частью Хонсю. Небольшие группы мелких слоевищ встречаются на мысах в Приморском и Хабаровском краях. Ламинария узкая наряду с ламинарией японской считается ценным съедобным видом.
У ряда видов ламинарий пластина молодых слоевищ целая, но с увеличением размеров под действием движения воды она рвется на продольные лопасти. Такова, например, ламинария палъчаторассеченная (L. digitata). Она имеет широкоовальную или клиновидную пластину, рассеченную, а в спокойных местах целую - в таком случае она бывает плоской или полусферически-изогнутой. Орган прикрепления - ризоиды. Слизистые каналы имеются только в пластине. Спорангии наблюдаются зимой и весной в виде округлых или овальных пятен, очертания которых на обеих поверхностях пластины не совпадают. Слоевище у ламинарии пальчаторассеченной многолетнее, длина его достигает в отдельных случаях 5 м, а ширина пластины - 1 м. Растет в сублиторали, иногда в прибойных местах встречается в нижнем горизонте литорали. Этот вид распространен в Северной Атлантике от Карского моря и Гудзонова залива на севере до полуострова Бретань и Нью-Йорка на юге.
У берегов Европы растет ламинария северная (L. hyperborea). Она напоминает ламинарию пальчаторассеченную, но отличается от нее наличием слизистых каналов в стволе и сильным сужением между старой и новой пластинами, а также другими признаками. Она встречается от Шпицбергена и Кольского полуострова до Средиземного моря (Балеарские острова и Миссинский пролив). У берегов Северной и Средней Европы она растет от поверхности до глубины 15-25 м, а в Миссинском проливе ее заросли находятся на глубине 50-100 м в местах с постоянным течением. Ламинария северная служит одним из основных источников получения альгинатов в странах Западной Европы и является поэтому объектом всестороннего изучения. На ней выясняли, как растительноядные водные животные влияют на вертикальное распределение водорослей. Оказалось, что если регулярно уничтожать морских ежей глубже нижней границы ее произрастания, она начинает расти еще глубже; в Миссинском проливе ламинария северная не может расти выше 50 м из-за растительноядных животных.
По берегам северной части Охотского моря в выбросах часто встречаются слоевища двух видов ламинарий с рассеченной пластиной и стволом, лишенным слизистых каналов. Один вид - ламинария прижаторизоидная (L. арpressirhiza) - характеризуется образованием спорангиев на одной поверхности пластины; при их созревании лопасти свертываются в трубки. У другого вида - ламинарии наклонноризоидной (L. inclinatorhiza) - имеются длинные слабоветвящиеся ризоиды, образующие высокий и широкий конус, что позволяет водоросли расти на грунте, занесенном илом. У нее спорангии развиваются одновременно на обеих поверхностях пластины, очертания пятен спорангиев на них совпадают.
Из видов, похожих на ламинарию пальчаторассеченную, в северной части Тихого океана широко распространена ламинария Бонгарда (L. bongardiana). Слизистые каналы у нее развиты и в пластине и в стволе. В отличие от других видов рода ламинария, имеющих рассеченную пластину (кроме иногда ламинарии прижаторизоидной), щель каждого разрыва у ламинарии Бонгарда наклонена к поверхности пластины, и концы щели на разных поверхностях смещены друг относительно друга. Спорангии образуются на одной или двух поверхностях пластины, причем очертания групп спорангиев на них не совпадают. Облик слоевищ ламинарии Бонгарда крайне изменчив в зависимости от условий произрастания. В нижнем горизонте литорали с пологим уклоном дна и ровной поверхностью грунта, а также в сублиторали при сильных течениях растут слоевища с коротким стволом и узкой длинной пластиной, часто нерассеченной. Если поверхность в нижнем горизонте литорали неровная (нагромождения камней, выступы скал) или в сублиторали отсутствуют сильные течения, то слоевища имеют широкую и короткую пластину и длинный ствол. В мелких литоральных лужах произрастают некрупные слоевища с широкой пластиной и коротким (1-3 см) стволом. Нередко у слоевищ с широкой пластиной имеется один разрыв, который проходит через всю пластину и затрагивает верхнюю часть ствола. Течением каждая из половинок пластины закручивается вокруг своей части ствола. Получается слоевище с вильчато-разветвленным стволом и двумя пластинами. Заросли ламинарии Бонгарда встречаются в СССР от острова Итуруп (Курильские острова), вдоль восточного берега Камчатки до Берингова пролива. Вдоль берегов Северной Америки она распространена до 42 ° с. ш.
Ламинария копытная (L. solidungula) - вид, широко распространенный в Арктике, в том числе там, где не встречаются другие ламинариевые. Пластина ламинарии копытной удлиненно-яйцевидной формы, ствол расширяется снизу к верхней части и снова сужается при переходе к пластине. Слоевище прикрепляется к грунту диском. В суровых районах Арктики слоевища мелкие, с пластиной длиной 10-20 см. Благодаря тому что сохраняются пластины прошлых лет (иногда до трех-четырех пластин), общая длина слоевища достигает 60-70 см. У самой южной границы распространения, около полуострова Лабрадор, ламинария копытная вырастает до 2,5 м. Такое хорошее развитие вида именно у южной границы распространения позволяет предполагать, что он не встречается южнее не из-за неблагоприятных физико-химических факторов, а из-за конкуренции со стороны других видов, в том числе ламинариевых.
Ламинария йезоензис (L. yezoensis) растет у дальневосточных берегов СССР и у Северной Америки. У нее рассеченная на лопасти пластина, прикрепляющаяся к грунту диском; слизистые каналы в стволе отсутствуют, поэтому ее прежде путали с ламинарией пальчаторассеченной.
Наконец, у трех видов ламинарий слоевище имеет стелющийся по грунту побег с ризоидами (ризом) и вертикальными стволами, несущими по одной пластине. Из этих видов наиболее широко распространена ламинария длинноствольная (L. longipes). Ее заросли в зоне разрушения волн встречаются вдоль берега Тихого океана от Курильских до Алеутских островов и до 55 ° с. ш. у берегов Северной Америки.
При изучении в культуре у ламинарии длинноствольной обнаружены гаметофиты одного типа, без разделения на мужские и женские, которые были способны к длительному вегетативному росту и очень редко производили спорофиты. Ризом и ствол у спорофита многолетние.
Род филлариелла (Phyllariella) включает всего один вид - филлариеллу охотоморскую (Ph. ochotensis), встречающуюся в северной половине Охотского моря. Пластина у нее гладкая, с пучками волосков, слизистые каналы отсутствуют, орган прикрепления - диск.
Род челлманиелла (Kjellmaniella) характеризуется удлиненной, нерассеченной пластиной с вмятинами и выпуклостями в виде сложного орнамента, который расположен поперечными рядами, отходящими от срединной гладкой полосы. Орган прикрепления - ризоиды. К этому роду относятся 2 вида.
Челлманиелла тонколистная (К. gyrata) обладает тонкой пластиной с узкой неутолщенной срединной полосой. Распространена у южных Курильских островов и у острова Хоккайдо.
Челлманиелла толстолистная (К. crassifolia) имеет толстую пластину с широкой утолщенной срединной полосой. Встречается в Японском море по западному берегу Татарского пролива, у южной оконечности Сахалина и у острова Хоккайдо.
Род циматэре (Cymathere) включает 3 вида. У них удлиненная пластина, целая, с двумя - четырьмя продольными складками, простыми или утолщенными (складки-ребра).
Циматэре японская (С. japonica), растущая у южных Курильских островов, имеет 4 продольные складки. Иногда краевые складки выражены слабо, и тогда этот вид легко спутать с ламинарией японской. Различия между ними заключаются в расположении спорангиев. У циматэре японской спорангии развиваются только на одной стороне пластины в ее основании, оставляя свободной срединную полосу, ограниченную складками.
Две складки-ребра имеет циматэре двускладчатая (С. fibrosa), которая встречается у средних и северных Курильских островов. Циматэре трехскладчатая (С. triplicate) отличается от остальных видов рода тем, что ее слоевище прикрепляется к субстрату диском и на одной поверхности пластины проходят 3 складки-ребра, а на другой поверхности - 4. Циматэре трехскладчатая растет вдоль Тихоокеанского побережья от острова Уруп (Курильские острова) до Командорских и Алеутских островов и у берегов Северной Америки на юг до 48 ° с. ш.
Род костария (Costaria) включает всего один вид - костарию ребристую (С. costata), пластина которой отличается пятью грубыми продольными ребрами, из них 3 находятся на одной поверхности пластины, а 2 расположены между ними на другой поверхности (табл. 17). Кроме ребер, пластина имеет отверстия и пучки волосков, поверхность ее изборождена выпуклостями и вмятинами, число которых, так же как и число отверстий, возрастает в местах с ослабленным движением воды. На поверхности ствола заметны мелкие ребрышки. В основании ствола развиты ризоиды. Спорангии начинают появляться с июня в виде многочисленных пятен, расположенных в углублениях пластины; со временем пятна спорангиев разрастаются и сливаются в более крупные, выходя за пределы углублений. Слоевища однолетние, они появляются во второй половине зимы. В заливе Петра Великого их разрушение завершается к концу июля - началу августа; на севере Татарского пролива и у Курильских островов слоевища сохраняются до сентября - октября.
Костария ребристая растет в большом количестве как в защищенных, так и в прибойных местах, в последнем случае она встречается вместе с другими ламинариевыми, лучше противостоящими сильному движению воды. Костарию можно встретить в нижнем горизонте литорали и в сублиторали до глубины 22 м. Распространена она в Японском море от Татарского пролива на севере до Корейского полуострова и северной части Хонсю на юге, в юго-западной части Охотского моря, в прибрежье Северной Америки от островов Шумагина до Сан-Педро (Калифорния).
У рода агарум (Agarum) пластина с одним продольным ребром и многочисленными отверстиями (табл. 16, 5), пучки волосков и слизистые каналы отсутствуют. Прикрепляется слоевище ризоидами. Наиболее широко известен агарум продырявленный (A. cribrosum). Отверстия в его пластине бывают от мелких до крупных, диаметром 2 см, изменчива и ширина ребра. В спокойных местах растут слоевища с более мелкими и многочисленными отверстиями и с более узким ребром. Агарум продырявленный встречается на глубине 5-36 м на скалистых и илистых грунтах в открытых и полузащищенных местах, за исключением зоны разрушения волн. Заросли его наблюдаются в нижней сублиторали, тогда как большинство других ламинариевых обычно преобладает в верхней сублиторали. Такое произрастание агарума продырявленного по вертикали связано с тем, что в верхней сублиторали он не выдерживает конкуренции со стороны других ламинариевых, а глубже морские ежи и другие растительноядные животные предпочитают выедать другие ламинариевые, не трогая агарума, обладающего жестким слоевищем. Агарум продырявленный распространен в северной части Тихого океана от Берингова пролива до севера Японии, у побережья КНДР, омываемого Японским морем, и до штата Вашингтон (США). Он встречается также у атлантического берега Северной Америки от острова Элсмир до штата Массачусетс (США).
Род талассиофиллум (Thalassiophyllum) включает один вид - талассиофиллум решетчатый (Т. clathrus), имеющий на стволе одну или несколько дырчатых пластин, свернутых в виде конуса (табл. 16, 4). Отверстия у пластины фестончатые, вывернутые на внутреннюю сторону. Молодое слоевище имеет плоскую широкую целую пластину с отверстиями. Позднее по боковым краям пластины образуются две зоны роста, при этом каждый край сворачивается в трубку. Одновременно пластина разрывается вдоль на две половины, каждая из которых растет в дальнейшем за счет своей зоны роста, находящейся внутри конуса. По мере роста каждой пластины увеличивается число витков в конусах. Нижний конец конуса, уплотняясь, превращается в ветвь. У старых слоевищ появляются дополнительные пластины, которые возникают как выросты в основании старой пластины. Поэтому старые слоевища приобретают кустистый вид.
Талассиофиллум решетчатый растет в открытых местах в сублиторали и в лужах нижнего горизонта литорали у средних и северных Курильских островов, вдоль Тихоокеанского побережья Камчатки, у Командорских и Алеутских островов и у южного побережья Аляски.
Семейство артротамновые (Arthrothamnaceae)
У этих водорослей многолетнее слоевище с разветвленным стволом и несколькими лентовидными пластинами. Каждая пластина с двумя ушками (полукруглыми выростами) по углам основания. Новые пластины появляются на смену старым как выросты по одной из центра ушек. Таким образом, одна старая пластина дает две новые. Старая пластина разрушается, за исключением узких участков, которые идут от ствола к новым пластинам: они утолщаются и превращаются в новые ветви ствола. Семейство артротамновых включает один род с двумя видами.
Артротамнус курильский (Arthrothamnus kurilensis) обладает прямостоячими стволом и ветвями. Растет в открытых местах в нижнем горизонте литорали и в сублиторали до глубины 30 м у юго-западного побережья Сахалина и у Курильских островов (Итуруп, Уруп, Симушир). У артротамнуса раздвоенного (А. bifidus) ствол стелется по грунту и состоит из толстых треугольных пластин с ризоидами по бокам, от двух наружных углов каждой пластины отходят по одной такой же пластине или по узкой лентовидной пластине. Этот вид растет у открытых берегов в нижнем горизонте литорали и в сублиторали до глубины 25 м в районе, включающем остров Хоккайдо, Курильские острова, восточный берег Камчатки до Кроноцкого залива.
Семейство лессониевые (Lessoniaceae)
У лессониевых ствол разветвленный, на концах ветвей по одной пластине. Ветвление ствола и увеличение числа пластин происходит за счет продольного разрыва, который возникает интеркалярно на стыке основания пластины и вершины ствола; удлиняясь, разрыв достигает верхнего края пластины. Спорангии образуются на поверхности пластин (рис. 143, 1).

Рис. 143. Бурые водоросли: 1 - лессония (Lessonia); 2 - егрегия (Egregia); 3 - вершина ветви хормозиры Банкса (Hormosira banksii); 4,5 - химанталия удлиненная (Himanthalia elongata): 4 - молодое растение, 5 - растение с рецептакулами (а); 6 - вершина ветви цистозейры бородатой (Cystoseira barbata); 7 - вершина ветви цистозейры косматой (Cystoseira crinita); 8 - вершина ветви саргассума (Sargassum); 9 - вершина ветви турбинарии (Turbinaria)
Род лессония (Lessonia) имеет ветви с длинными междоузлиями, воздушные пузыри отсутствуют. Пластины делятся разрывом на две равные части. Известно 5 видов этого рода. Из них 4 вида растут в южном полушарии, где часто образуют настоящие подводные леса. Наиболее крупные слоевища имеет лессония чернеющая (L. nigrescens), растущая у берегов Перу, Чили и Фолклендских островов. Ее стволы достигают в толщину 10 см, а в высоту 4 м. Один вид - лессония ламинариевидная (L. laminariaeoides) встречается в северном полушарии, в северной половине Охотского моря, образуя вместе с ламинарией Гурьяновой заросли в нижнем горизонте литорали и верх-ней сублиторали. Слоевище ее достигает в длину 1-2 м; первое ветвление ствола расположено у самых ризоидов, выше междоузлия достигают в длину 30 см, пластины вытянутые, длиной 20-90 см. Спорангии встречаются с сентября до весны следующего года, они располагаются на обеих поверхностях пластин сплошным пятном.
Род макроцистис (Macrocystis) имеет ствол многократно разветвленный. Первые 2-3 ветвления снизу дихотомические, остальные односторонние. Верхушечная пластинка отчленяет новые пластинки с одной своей стороны посредством косых разрывов. Перед каждой пластинкой ветвь обычно расширена в грушевидно-веретеновидный воздушный пузырь. Благодаря воздушным пузырям и длинным ветвям основная часть слоевища макроцистиса плавает у поверхности моря. Это позволяет механизировать его добычу. Виды рода макроцистис перерабатывают для получения альгинатов и других химических продуктов.
Макроцистис грушевидный (М. pyrifera, табл. 16, 2) прикрепляется к субстрату ризоидами, слоевище его достигает размеров 30-50 м, длина пластин 1-1,5 м. Это многолетнее растение, отдельные экземпляры живут до 8-10 лет. Впервые спорангии появляются на шестимесячных экземплярах. Спорангии образуются на нижних пластинах, сидящих на вильчато-разветвленных коротких побегах в основании слоевища. Эти пластины иногда называют спорофиллами. Спорангии могут появляться также на верхних пластинах, плавающих у поверхности. Созревание спорангиев происходит сначала на верхней, более старой, части пластины. Развитие отдельного спорангия завершается за две недели.
Макроцистис грушевидный встречается у открытых берегов на глубине 6-40 м, иногда его обнаруживали на глубине до 60 м. Распространен в южном полушарии у Южной Америки от мыса Горн до Перу вдоль западного берега и до 50° ю. ш. у восточного берега, около Тасмании и Новой Зеландии, у южной оконечности Африки; в северном полушарии растет у Тихоокеанского побережья Северной Америки от Ситки до полуострова Калифорния. В середине прошлого века макроцистис грушевидный находили в Охотском море, у берегов Камчатки и острова Уналашка, сейчас он здесь не встречается.
Макроцистис грушевидный имеет большое практическое значение, поэтому в настоящее время его экологию интенсивно изучают, особенно у берегов Калифорнии, где находятся наиболее крупные предприятия по его переработке. Обнаружено, что большой урон макроцистису могут причинять морские ежи. Они способны полностью уничтожать целые заросли этой водоросли. Массовая гибель крупных слоевищ происходит в годы с необычно сильным летним прогревом воды. При этом гибнут побеги, поднимающиеся до перегретого слоя воды. На молодых слоевищах, находящихся глубже, прогрев поверхностных слоев воды не отражается. Эффективный радиус рассеивания спор, при котором они дают новые растения, составляет у отдельно растущих слоевищ макроцистиса грушевидного 5 м. В зарослях возобновление слоевищ происходит на удалении до 90 м. Расселение на большие расстояния осуществляется посредством оторванных пластин со спорангиями.
Макроцистис целънолистный (М. integrifolia) имеет стелющееся основание с ризоидами (ризом). Слоевище многолетнее, высотой до 5-8 м, длина пластин 25-60 см. Растет до глубины 7 м у Тихоокеанского побережья Америки от острова Ванкувер до залива Камель в Калифорнии, у побережья Перу и северной части Чили. В Ботаническом институте АН СССР в Ленинграде хранятся целые экземпляры макроцистиса цельнолистного длиной до 0,5 м, собранные в 1870-1871 гг. у Сахалина вблизи Александровска-Сахалинского и в заливе Анива. Позднее макроцистис у берегов Азии не встречался.
Род нереоцистис (Nereocystis) включает один вид - нереоцистис Лютке (N. luetkeana, табл. 19, 2). У него длинный ствол, достигающий 15-25 м, который постепенно расширяется кверху и несет на конце пузырь диаметром 12-20 см. На верхней части пузыря растут две короткие дихотомически-ветвящиеся веточки длиной в несколько сантиметров с длинными узкими пластинами на концах. Длина пластин достигает 3-9 м. Всего таких пластин на слоевище бывает 24-40. Спорангии образуются на пластинах. Слоевище прикрепляется к грунту ризоидами, которые образуют конус диаметром до 30 см. Несмотря на гигантские размеры, слоевища у нереоцистиса однолетние. Они появляются в феврале - марте, спороносят в ноябре и исчезают (отрываются от грунта) в декабре - январе; крайне редко единичные спороносящие экземпляры встречаются до апреля. Растет нереоцистис на глубине до 20 м, при этом верхняя часть ствола с пузырем плавает у поверхности. Часто он образует настолько густые заросли, что через них невозможно пробраться на лодке. Нереоцистис распространен у Тихоокеанского побережья Северной Америки от острова Уналашка до Калифорнии. Свободноплавающие слоевища встречаются у Алеутских островов. В СССР эту водоросль можно найти в выбросах на берегах Командорских островов, обычно это стволы с пузырем, но без пластин. Крайне редко плавающие стволы встречаются в Беринговом проливе.
Род пелагофикус (Pelagophycus) включает один вид - пелагофикус пора (Р. роrrа). В строении его слоевища много общего с нереоцистисом. Отличие заключается в длинной ветви (до 1,6 ж), отходящей от пузыря. Первое ее ветвление дихотомическое, следующие - односторонние, как у макроцистиса. Слоевище несет около 10 пластин длиной по 4-5,5 м и шириной 29-45 см. Общая длина слоевища достигает 50 м. Растет пелагофикус вместе с макроцистисом у берегов Южной Калифорнии.
Семейство алариевые (Alariaceae)
У этих водорослей на вершине ствола расположена пластина с продольным ребром или лишенная его. Края пластины ровные или с длинными боковыми лопастями. По бокам сильно или слабо уплощенного ствола у многих представителей имеются пластинки на тонких стебельках (спорофиллы) или развивается сплошная складчатая кайма. Спорангии образуются на спорофиллах, кайме или на лопастных выростах центральной пластины. В поверхностном слое клеток пластинчатых образований часто наблюдаются гландулярные клетки, наполненные физодами. Они желтые в молодых частях слоевища и темно-бурые, почти черные в старых.
Род алария (Alaria) характеризуется пластиной с продольным ребром, идущим от ствола до вершины. Спорофиллы расположены на тонких стебельках по бокам ствола (рис. 144). Все виды рода алария распространены в северном полушарии и предпочитают места с постоянным движением воды.
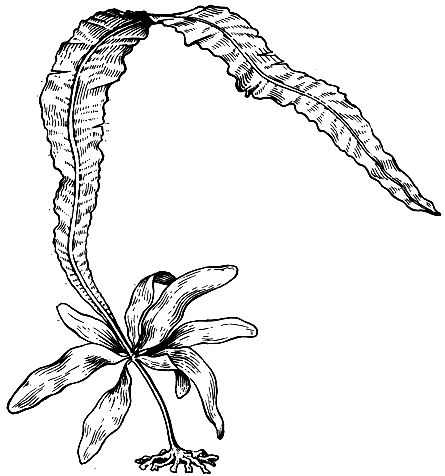
Рис. 144. Алария (Alaria)
В Атлантическом океане наиболее широко распространена алария съедобная (A. esculenta). Ее слоевища достигают в длину 2-3 м. Столь же широко распространена в Тихом океане алария окаймленная (A. marginata), имеющая такие же размеры.
Наиболее крупное слоевище у аларии полой (A. fistulosa). Для нее характерно наличие полого ребра в пластине. Местами полость в ребре пересекается поперечными перегородками, в результате получаются как бы длинные камеры, наполненные газом. Благодаря этому слоевище имеет положительную плавучесть, и она сохраняется даже при повреждениях ребра, например при разрушении вершины. Плавающие обрывки пластин аларии полой встречаются далеко за пределами границ ее произрастания. Обычно алария полая растет так, что вершины ее пластин достигают поверхности воды и стелются под ней. Максимальная глубина, на которой наблюдали этот вид, составляет 35 м. По некоторым данным, у средних Курильских островов встречаются заросли аларии полой, состоящие из растений длиной 41 м.
Алария полая распространена от острова Итуруп (Курильские острова) до южной части Берингова моря, а также у Командорских и Алеутских островов и вдоль Тихоокеанского побережья Аляски. Небольшие заросли ее встречаются у Камня Опасности в проливе Лаперуза (между островами Сахалин и Хоккайдо). Народы, живущие по берегам тех мест, где она растет, называют аларию полую бобровой капустой. Это связано с тем, что часто вблизи мест ее произрастания водится калан (его еще называют камчатским бобром). Виды рода алария в холодных водах северного полушария столь же обычны, как виды рода ламинария. Их и используют сходным образом. Некоторые народы, например населяющие Алеутские и Командорские острова и побережье Чукотского полуострова, употребляют виды рода алария в пищу. Особо ценятся спорофиллы аларии узкой (A. angusta), отличающиеся большой толщиной и мягкостью. На Чукотке пластинки аларии мочат в пресной воде до тех пор, пока ребро не станет мягким.
Представители рода ундария (Undaria) растут в тепловодных местах Японского моря, в Желтом море и в северной части Восточно-Китайского моря. Особенно широко распространена ундария перистая (U. pinnatifida). Она имеет цельнокрайнюю или лопастную пластину с пучками волосков. Для прикрепления к грунту у нее служат ризоиды. Слоевища всегда однолетние. В начале лета по краям ствола начинает расти складчатая кайма, на которой образуются спорангии. В августе спорангии созревают, и слоевища начинают разрушаться. Ундария перистая обладает нежной пластиной, ее используют в пищу. В Японии ее культивируют на специальных морских плантациях. В СССР ундария перистая растет в заливе Петра Великого.
Род эклония (Ecklonia) встречается у берегов Японии, Австралии, Новой Зеландии и южной оконечности Африки. Для эклонии характерно развитие спорангиев на лопастных выростах по бокам центральной пластинки. У самого крупного вида - эклонии трубовидной (Е. buccinalis), растущей у южной оконечности Африки, лопасти во много раз крупнее центральной пластинки. Слоевища эклонии трубовидной достигают в длину 8 м. По внешнему облику они напоминают нереоцистис. У них длинный (до 5 м) ствол, расширяющийся кверху и полый.
Для представителей рода егрегия (Egregia) характерно наибольшее расчленение слоевища среди алариевых и других ламинариевых. Эти водоросли растут у Тихоокеанского побережья Северной Америки (рис. 143, 2).
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'