Сейсмонастии
Сейсмонастические ответные реакции - самые быстрые и наиболее заметные в мире растений. Как и тигмонастические движения, о которых еще пойдет речь, они вызываются механическими причинами. Особенно раздражающе действуют сотрясения и удары. Об этом свидетельствует и их название, поскольку "сейсмос" по-гречески означает "сотрясение" или "землетрясение". Уже падающая дождевая капля или порыв ветра может вызвать раздражение. Как и при проявлении других настий, направление движения органа определяется его строением. Поскольку сейсмонастически реагирующие растения оказываются особенно интенсивно исследуемыми объектами, мы расскажем о сейсмонастиях несколько подробнее, чем о до сих пор рассмотренных настиях.
Наиболее известные сейсмонастические изменения положения обнаруживают листья мимоз. Мimosa pudica, которую называют чувствующим растением, займет центральное место в нашем рассказе. Отклоняясь от до сих пор использовавшейся нами схемы изложения материала, мы рассмотрим процессы, протекающие между раздражением и ответной реакцией, сперва у этого классического объекта, а уж затем обратимся к другим примерам. Конечно, вам известно это растение, разводимое в оранжереях ботанических садов. Каждый из его листьев состоит из черешка, обычно четырех лучеобразно расходящихся от его вершины черешочков с попарно (перисто) расположенными листочками. У оснований черешка, черешочков и листочков находятся подушечки сочленений (рис, 34, а), состоящие главным образом из паренхимы, образованной тонкостенными клетками. Поскольку эта паренхима участвует в движениях сочленении, ее называют моторной тканью. В.первичном сочленении, где черешок отгибается вниз, она расположена у нижней его стороны. Моторная ткань вторичных сочленений, с помощью которых черешочки сближаются друг с другом, находится внутри, а такая же ткань у оснований отгибающихся кверху листочков - на верхней стороне. Своим названием, которое мимозы получили еще в XVII в., они обязаны своей способности резко изменять свой внешний вид (слово "мимос" греческое и может быть переведено как актер или подражатель).
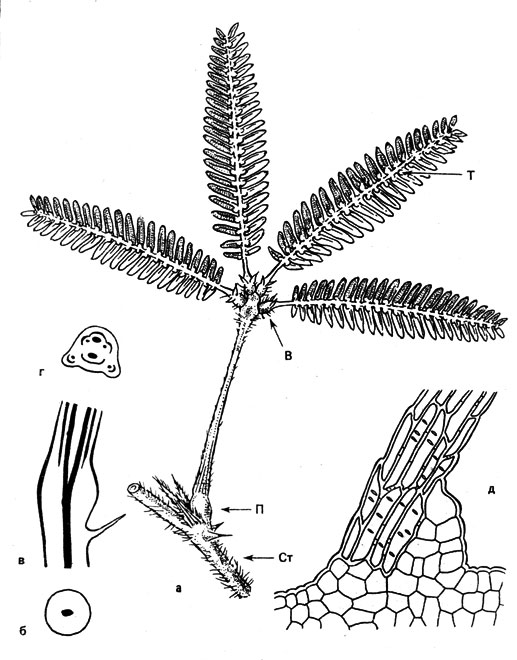
Рис. 34. Морфология, и анатомия листа Mimosa pudica: а - схематическое изображение строения листа; П, В и Т - первичное, вторичное и третичное сочленения; Ст - стебель; б - сочленение черешка (первичное сочленение) на поперечном срезе; проводящие и механические ткани показаны (как и на в и г) черным цветом; в - переход сочленения в собственно черешок (продольный срез); г - черешок выше сочленения (поперечный срез); д - нижняя часть раздражимой щетинки подушечки первичного сочленения (клеточное строение)
Если в черешке относительно жесткие проводящие пучки расположены по его периферии, то в сочленении мы видим их объединенными в один центральный тяж (рис. 34). Благодаря этому возрастает способность к движению.
В зависимости от того, воздействует ли раздражитель непосредственно или в результате проведения возбуждения, различают прямое и непрямое раздражения. Раздражим весь побег, и сотрясение всего растения тоже приводит к проявлению соответствующей ответной реакции. К прикосновению особенно чувствительны нижние стороны первичных сочленений, поскольку здесь находятся жесткие раздражимые щетинки, передающие давление подушечкам сочленений (рис. 34, д). Речь идет о трихомах (волосках), имеющих длину от 1 до 2,5 мм. Они состоят из продольно вытянутых клеток с плотными одревесневшими оболочками. Такого рода стимулятор расположен на паренхимном основании, которое при прикосновении к щетинке растягивается и придавливается.
Растение реагирует не только на сотрясения, но также и на другие воздействия, о чем уже говорилось при обсуждении движений, обусловленных фото- и термонастиями. Например, в темноте положение черешков, черешочков и листочков подобно тому, какое бывает после сейсмонастического раздражения. Описаны также электро- и травматонастические реакции. Наконец, следует еще отметить возможность изменений положения, обусловленных тропизмами. Так, Mimosa pudica способна, варьируя положения листочков, обеспечивать оптимальное использование света. Например, во второй половине дня солнечные лучи падают сбоку, и листочки располагаются соответствующим образом. В основе сейсмонастических изменений органами своего положения лежат колебания тургорного давления. Если порог раздражения превзойден, ответная реакция проявляется в полной мере. Следовательно, речь идет о процессах, протекающих в основном в соответствии с законом "все или ничего". Латентный (скрытый) период сравнительно быстрых движений листа составляет около 0,08 с, а время осуществления ответной реакции здесь по меньшей мере может быть приравнено к продолжительности латентного периода. Вопросы, связанные с рефрактерной стадией (стадией невосприимчивости), мы еще рассмотрим подробнее. Особо следует учитывать сильную зависимость раздражимости от температуры и влажности воздуха. Способность к осуществлению ответной реакции бывает максимальной при 40° С, тогда как при температурах ниже 12,5° и выше 60° С она утрачивается.
Для сейсмонастических движений характерно быстрое и бросающееся в глаза проведение возбуждения. Оно относительно просто может быть продемонстрировано на следующих друг за другом движениях листочков. Так, например, если подвергнуть раздражению листочки, находящиеся на верхушке черешочка, то листочки попарно друг за другом сблизятся, отклонившись косо вверх, черешочки сблизятся, а затем отогнется вниз и черешок. При сильном раздражении отдельного листа или даже листочка возбуждение, распространяясь вверх и вниз по стеблю (примерно на 50 см), может дойти до других листьев. При этом ответные реакции проявляются сначала в первичных, затем во вторичных и, наконец, в третичных сочленениях. Скорость проведения возбуждения достигает - в зависимости от температуры - от 0,4 до 3 см/с. Если же растение ранено, максимальная скорость проведения возбуждения может доходить даже до 10 см/с. В общем, у Мimоsа различают два разных механизма проведения возбуждения.
При химическом проведении возбуждения раздраженными клетками выделяется возбуждающее вещество, которое может перемещаться в растении по проводящей и основной тканям. Оно проходит не только через мертвые участки тканей, но и через заполненную водой стеклянную трубочку, соединяющую отрезанный лист с местом, от которого он был отделен. Если это вещество (предположительно речь идет о содержащей азот аминокислоте) дойдет до клеток сочленения, то есть до ткани, способной осуществлять движение, то последует ответная реакция. Выделенное в почти чистом виде возбуждающее вещество, поднимающееся в срезанных побегах мимозы по их проводящим тканям, оказывает свое действие уже при концентрации 10-8 г/мл. Раздражающе могут действовать также аминокислоты и другие соединения. Например, как показали исследования Шилдкнехта с сотрудниками (1978), смесь, состоящая из 20% L-глютаминовой кислоты и 80% β-аланина, проявляет особенно сильное раздражающее действие (табл. 3). При проведении этих опытов двух-, четырехнедельные листья Mimosa рudiса, срезанные лезвием безопасной бритвы ниже первичного листового сочленения (рис. 34, а), помещали в водные растворы аминокислот; при этом для проведения опытов наиболее благоприятными оказались температуры от 20 до 30° С. Скорость химического проведения раздражения достигает примерно от 0,15 до 2 см/с. Упомянем и второй способ проведения возбуждения - электрический. При нем потенциал действия распространяется со скоростью от 2 до 5 см/с. Путями его проведения, по-видимому, служат продольно вытянутые клетки паренхимы (основной ткани), находящиеся в проводящих пучках. Мертвые или переохлажденные сочленения или ткани вряд ли участвуют в проведении возбуждения. Но возбуждающее вещество, продуцируемое клетками, может обходить такого рода участки и вызывать возбуждения на другой стороне.
Таблица 3. Активность аминокислот и их смесей как раздражителей. Приведенная продолжительность осуществления ответной реакции типична для соответствующей кислоты или смеси. Если графа "примечания" оставлена незаполненной, то это значит, что листочки Mimosa pudica после осуществления ответной реакции остаются сблизившимися
| Аминокислоты или их смеси | Время осуществления реакции, средние величины |
Примечания |
| L-глютаминовая кислота (0,1 м) | 44″-1′10″ | Периодическое сближение и расхождение листочков |
| α -аланин (0,1 м) | 33″-1′39″ | Листочки после сближения расходятся |
| L-глютаминовая кислота: β -аланин (1:1) | 9″-45″ | * |
| L-глютаминовая кислота: D,L-серин (1:1) | 20″-1′ | * |
| α -аланин: D,L-серин (1:1) | 17″-1′30″ | * |
| L-глютаминовая кислота: α -аланин: D,L-серин (1:1:1) | 20″-1′ | * |
Что мы знаем о процессах, проходящих в сочленениях? Как уже было упомянуто при обсуждении основных положений физиологии раздражений, возбуждение характеризуется проявлением потенциала действия. У Mimosa pudica он достигает примерно 140 мВ; в это время потенциал покоя, как правило, равен -160 мВ, но после раздражения он увеличивается на -20 мВ. Согласно нашим современным представлениям электрофизиологические явления зависят прежде всего от ионов соединений хлора и калия, концентрация которых внутри клетки регулируется с помощью ионных насосов. Возбуждение моторных клеток приводит к временному нарушению структуры плазмалеммы. С этим связаны увеличение проницаемости мембраны, временная приостановка деятельности ионных насосов, выход ионов хлора и калия, а также резкое падение тургора. Кроме того, клеточный сок выходит из вакуоли в клеточную оболочку и в межклетники. Невооруженным глазом можно видеть, что нижняя сторона первичного сочленения становится явно более темной. Одновременно с потерей тургора моторными клетками, находящимися на нижней стороне, клетки верхней стороны частично теряют свою сопротивляемость. В уравнении сосущей силы это величина А. Следовательно, на верхней стороне увеличивается сосущая сила и соответственно поглощение воды, и лист опускается. Теперь можно также понять, почему при проявлении закона "все или ничего" подпороговые раздражения не суммируются, а ответная реакция наступает лишь после того, как будет превышен порог раздражения. Очевидно, это зависит от состояния мембраны. Возможно, что в выведении (выдавливании) воды участвуют и сократительные белки. На это указывает, в частности, высокое содержание АТФ в моторных клетках, сильно падающее во время движения.
Для сейсмонастических движений типичны также стадии абсолютной и относительной невосприимчивости, в течение которых после нового раздражения возбуждение вовсе не наступает или проявляется лишь в меньшей степени. В первичном сочленении стадия абсолютной невосприимчивости продолжается от 15 до 40 с, а относительная - примерно 360 с. Во время абсолютной невосприимчивости CL- К+ выходят из клетки; при этом всякое последующее столь же сильное раздражение не оказывает действия. Во время относительной невосприимчивости снова начинают действовать ионные насосы, и ионы С1- и К+ снова поступают в клетку, но потенциалы действия еще не могут проявиться в полную силу. Специалисты различают стадии невосприимчивости не только в отношении возбудимости, но и в отношении движений. Такая стадия абсолютной невосприимчивости продолжается до тех пор, пока снова не станет проявляться ответная реакция (около 90 с). Стадия относительной невосприимчивости (в отношении движений) заканчивается лишь через 10-30 мин, когда лист вернется в первоначальное положение.
Для фазы восстановления исходного состояния характерны разные ступени. В результате осуществления процессов, требующих затрат энергии, в первую очередь восстанавливаются полупроницаемость мембран, деятельность ионных насосов и первоначальное ионное равновесие. Осмотическое же поглощение воды происходит без затрат энергии.
Особенно интересно, что продолжительность стадии невосприимчивости не зависит от того, о каком раздражении идет речь - механическом, химическом, электрическом, травматическом или каком-либо другом. Следовательно, в основе настических движений мимоз во всех случаях лежат одинаковые клеточно-физиологические процессы.
Кроме Mimosa рudiса, осуществлять сейсмонастические движения листьев могут и многие другие растения. Прежде всего это растения - обитатели тропиков. В качестве примеров можно назвать представителей семейства мимозовых Mimosa invisa и Neptunia oleracea, а также семейства кисличных - Biophytum sensitivum и Охаlis sensitiva. Особенно быстро ответные реакции происходят у Biophytum. Они могут завершиться в течение 1 с. Обратное движение продолжается 3 мин. На эти бросающиеся в глаза движения указывает и название Biophytum ("живое" растение). Очевидно, в то время, когда возникло это название, такого рода способность к движению у цветковых растений считали совершенно необычной. В небольшой степени сейсмонастичны также листья кислицы обыкновенной (Охalis acetosella) и робинии (так называемой белой акации - Robinia pseudo-acacia). Даже сильные сотрясения вызывают у этих растений нашей флоры слабые ответные реакции, и закон "все или ничего" к таким движениям неприменим. Между раздражимыми и не раздражимыми объектами существуют все переходные ступени.
Следующие примеры хорошо заметных изменений положений органов в пространстве, обусловленных сейсмонастиями, - это движения тычинок. Например, у барбариса обыкновенного (Веrberis vulgaris) тычинки резко изгибаются снаружи внутрь (рис. 35), тогда как у видов солнцецвета (Неlianthemum), у относящейся к семейству липовых комнатной липы (Sparmannia africana) и других растений они изгибаются в противоположном направлении. У тычинок Веrberis способен воспринимать раздражение только базальный участок внутренней стороны тычиночной нити, а у тычинок Неlianthemum - базальный участок ее наружной стороны. В этих местах находятся моторные клетки, и поэтому там происходит изгибание. Движение тычинок Веrberis (ср. табл. 1 и рис. 4) - это одни из самых быстрых движений, известных в мире наших растений. Проведения возбуждения от одной тычиночной нити к другой здесь не бывает. Напротив, в пучке тычиночных нитей Sparmanniа раздражение одной нити приводит к изгибанию всех нитей.
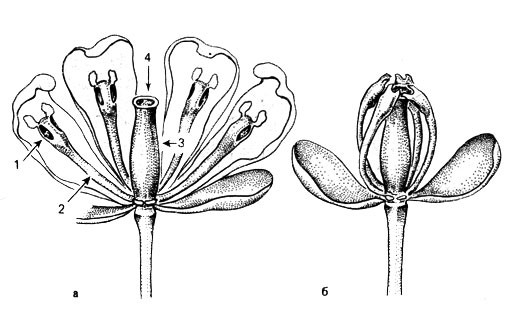
Рис. 35. Цветки Berberis vulgaris (чашелистики и лепестки частично удалены) с нераздраженными (а) и раздраженными (б) тычинками, 1 - пыльник, 2 - тычиночная нить, 3 - столбик, 4 - рыльце
Несколько по-иному происходит движение сейсмонастически раздражимых тычинок многих сложноцветных. Например, тычиночные нити васильков (виды Centaurea), весьма обычных растений наших лугов, полей и других местообитаний, отвечают на прикосновение сильным сокращением. Потеря тургора симметрично расположенными моторными клетками приводит к укорочению тычинок на 20-30% первоначальной их длины. При этом столбик с еще нераскрывшимся рыльцем, находящийся внутри трубки, образованной пыльниками, и действующий как поршень, выдавливает пыльцу из трубки наружу. Затем пыльцу разносят насекомые-опылители. Если латентный (скрытый) период продолжается менее секунды, то на осуществление ответной реакции уходит несколько секунд, а для возвращения к исходному положению требуются уже минуты.
В удержании пыльцы могут принимать участие и раздражимые рыльца, что, например, можно видеть у растущих у нас видов губастика (Mimulus). Моторная ткань находится на внутренней стороне каждой из лопастей рыльца. Если насекомое к ней прикоснется, то лопасти сближаются и при этом приставшая к насекомому пыльца может быть снята. Возбуждение может передаваться от одной лопасти к другой. Ответная реакция происходит в течение нескольких секунд, а возвращение лопастей рыльца в исходное положение продолжается до 45 мин.
Во всех названных случаях настические изменения положения происходят в результате потери тургора моторными клетками, расположение которых обусловливает разные способы осуществления ответных реакций в виде движений тычинок и рылец. Как и при движениях листьев мимоз, речь здесь также идет об ответных реакциях типа "все или ничего" с явно выраженными стадиями невосприимчивости. Но в отличие от Mimosa в тычинках и рыльцах обычно не обнаруживается проведение возбуждения. В качестве исключений уже были названы Sparmannia и Mimulus.
В некоторых специальных приспособлениях, служащих для ловли животных, например, таких, как листья росянок, тоже проявляются сейсмонастические движения. Известные примеры этого - растущая в наших водоемах альдрованда пузырчатая (Aldrovanda vesiculosa) и встречающаяся в приатлантических областях Северной Америки венерина мухоловка (Dionaea muscipula). Именно у последней в 1769 г. была впервые обнаружена насекомоядность растений. Захватывающие листья Diоnaeа могут очень быстро складываться. На верхних, то есть внутренних сторонах половин листовой пластинки, имеющих по краям зубцы, находятся пищеварительные железки, раздражимые сосочки и по три раздражимых щетинки. Когда половины пластинки сближаются, зубцы одной из половин заходят в промежутки между зубцами другой, и добыча удерживается надежно. Раздражение вызывает потерю тургора моторных клеток, имеющихся на верхней стороне, и сильное растяжение клеток нижней стороны листовой пластинки. Вслед за этим быстрым ловчим движением еще происходит более медленное дальнейшее сближение половин пластинки. Здесь речь идет уже о процессах роста на наружной стороне, и половины листовой пластинки еще крепче смыкаются вокруг насекомого. Если пойманным оказывается живой объект, то половины пластинки остаются сблизившимися в течение дней и даже недель; если же объект "несъедобен", то через несколько часов они снова расходятся. Очевидно, при этом также действуют химические и контактные раздражения. Открывание, т. е. отхождение половин пластинки друг от друга, происходит в результате восстановления осмотического потенциала, а также усиления роста клеток верхней стороны.
Служащие стимуляторами раздражимые щетинки передают раздражение специфическим чувствительным клеткам, находящимся в основаниях щетинок. Половины листовой пластинки, лишенные щетинок, теряют способность воспринимать раздражения. Как правило, для проявления движений требуется несколько раздражений. Вопросы, касающиеся суммирования раздражений, еще не выяснены. Aldrovanda и Dionaea обнаруживают потенциал действия примерно в 100 мВ. Он распространяется от основания раздраженной щетинки во все стороны со скоростью от 6 до 20 см/с. Здесь перед нами самое быстрое проведение возбуждений из всех измеренных у растений до сих пор. Как и при движениях тычинок Веrberis, латентное время у Dionaea составляет 0,1 с, время ответной реакции - 0,2 с, а ее продолжительность - 0,1 с. Закон "все или ничего" у Aldrovanda и Dionaea проявляется только при благоприятных условиях. Обычно степень проявления ответной реакции зависит от интенсивности раздражения. Наконец, следует отметить четырехсекундную стадию невосприимчивости у Dionaea.
В заключение рассказа о сейсмонастиях рассмотрим еще и вопрос об их биологическом значении. В отличие от объяснений значения движений раздражимых тычинок и рылец, способствующих воспроизведению растений, и ловчих листьев, выполняющих задачи, связанные с физиологией питания, на вопрос о значении движений листьев у мимоз ответить труднее. Однако следует исходить из того, что они развились в течение филогенеза и сохранились при естественном отборе как дающие растениям определенные преимущества. Какие же преимущества предоставляет бросающееся в глаза изменение внешнего вида мимозы, которая после сотрясения выглядит как увядшее растение? Поскольку одно только складывание листочков приводит к уменьшению поверхности примерно наполовину, а ответные реакции в первичном и вторичных сочленениях еще больше уменьшают общую поверхность растения, ответ на вопрос "для чего" должен лежать в этой плоскости. По-видимому, положение испытавших раздражение листьев вызывает ограничение транспирации (пассивное испарение воды живым растением). Особенно при сильном, иссушающем ветре это может защитить от излишней потери воды. Это, пожалуй, согласуется и с раздражающим действием ветра.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'