Движения хлоропластов
Как уже упомянуто в рассказе о растительной клетке, в хлоропластах происходит фотосинтез. Речь идет о важнейшем биохимическом процессе, осуществляющемся на Земле, которому мы обязаны энергетической основой нашей жизни - пищей и топливом. Эволюционируя, растения выработали механизмы регуляции, которые, во-первых, обеспечивают оптимальное использование света, необходимого для фотосинтеза, и, во-вторых, защищают фотосинтезирующий аппарат от слишком сильного освещения. Этой цели, в частности, служат так называемые ориентировочные движения хлоропластов, обнаруживающиеся у водорослей, мхов, папоротников и семенных растений. В листочках мхов и заростках (геметофитах) папоротников при слабом освещении хлоропласту располагаются у физически верхних и нижних стенок клеточных оболочек, где можно абсорбировать больше света (рис. 9). При сильном освещении хлоропласты отходят к боковым стенкам и тем самым оказываются, "защищенными". В связи с этим говорят о позитивной и негативной ориентировках хлоропластов. Что же касается механики движений, то она может быть двоякой, Если, как, например, у рдеста (Роtаmogeton) направление движения хлоропластов не совпадает с направлением движения других компонентов протоплазмы, мы можем сделать вывод о том, что их перемещение активно. Напротив, результаты исследования водоросли вошерии (Vаuсhеria), у которой как хлоропласты, так и другие компоненты протоплазмы перемещаются в одном и том же направлении, свидетельствуют о пассивном их движении. При исследовании влияния света на перемещение хлоропластов прежде всего следует попытаться выявить соответствующий фоторецептор. За исключением водорослей класса конъюгат, или сцеплянок (Conjugatophyceae), у которых красный свет вызывает позитивные ориентировочные движения хлоропластов, а синий свет - негативные, у всех прочих исследованных объектов только коротковолновая часть спектра видимого света обусловливает перемещение хлоропластов. Поэтому только желтые пигменты можно считать фоторецепторами. При этом с очень большой степенью вероятности здесь можно говорить об одном из флавинов, вернее - флавопротеидов, и большинство исследователей в связи с этим называет рибофлавин. Но в пластидах флавины не обнаруживаются. Сравнение спектра поглощения этого пигмента со спектрами проявления как позитивных и негативных ориентировочных движений хлоропластов, так и движения цитоплазмы, а также ее вязкости показывает, что все эти спектры имеют очень большое сходство. По-видимому, этот рецептор способен вызывать не одну, а многие ответные реакции.
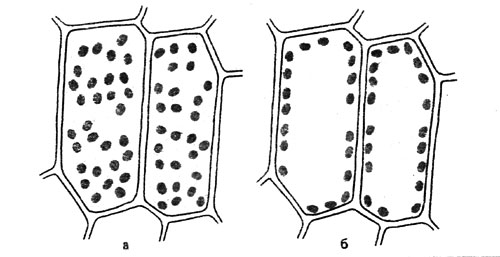
Рис. 9. Движение хлоропластов у Funaria hygrometrica: а - положение хлоропластов при слабой освещенности (около 50 лк); б - их положение при сильной освещенности (около 1000 лк)
Фоторецептор синего света до сих пор еще не удалось уверенно выявить, а упомянутое действие красного света на водоросли-сцеплянки мужоцию (Мougeotia) и мезотениум (Мesotaenium) свидетельствует в пользу того, что в роли фоторецептора здесь выступает фитохром.
Что же касается локализации рецепторов, то опыты, проведенные как на ряске(Lemna), плаунке (Sеlaginеllа), валлиснерии (Vаllisneria), так и на водоросли-сцеплянке Mesotaenium, показали, что оба фоторецептора - флавин и фитохром - сосредоточены в эктоплазме, находящейся в состоянии, подобном гелю. Как показывают исследования, проведенные с применением поляризованного света, молекулы фитохрома 560 расположены вдоль поверхности в виде спирали под углом 45°, а молекулы фитохрома 730 - перпендикулярно поверхности.
Наряду с механизмом восприятия световых сигналов и связанных с ним вопросов большой интерес представляет также проблема ориентирования хлоропластов при сильном и слабом освещении. Посредством многочисленных экспериментов удалось показать, что управление движением хлоропластов зависит от градиентов поглощения. При этом речь идет о различиях в яркости света, которые возникают по-разному. Например, при прохождении света сквозь клетку происходит его ослабление (принцип затенения). Но клетки, между прочим, могут вести себя и как собирательные линзы, причем сзади" света оказывается больше, чем "спереди". Затем к появлению градиентов поглощения может вести специфическое расположение молекул фоторецептора. У зеленой водоросли Hormidium обнаружен эффект действия линзы, тогда как у сцеплянок Mougeotia и Мesotaenium градиент поглощения обусловлен положением фоторецепторов. Между движением хлоропластов и фотодинезами существуют тесные связи.
О возможности проявления пассивных и активных движений хлоропластов мы уже говорили. И в тех и в других движениях, вероятно, участвуют белковые фибриллы типа актомиозина. При пассивных перемещениях сократительные белки находятся между экто- и эндоплазмой. Таким образом, цитоплазма перемещается вдоль фибрилл. Напротив, активное движение хлоропластов происходит в результате сокращения фибриллярных структур, контактирующих с поверхностью каждого из них. Для наглядности объяснения иногда используют действие весла, которое может вызвать, как скольжение гребной шлюпки вперед, так и движение окружающей шлюпку воды.
Движение хлоропластов, как и движение цитоплазмы, связано с реакциями, освобождающими энергию. Как можно заключить исходя из результатов опытов с Lemna и Mougeotia, АТФ в первую очередь образуется при процессах дыхания, соответственно окислительного фосфорилирования. Происходящее во время фотосинтеза фотофосфорилирование также может быть причастно к освобождению энергии.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'