Глава 8. Сообщества-хамелеоны (Циклическая динамика)
Говоря о горизонтальной структуре сообщества, мы подчеркивали ее динамический характер. Это одна из сторон общей циклической динамики сообщества, которая, отражая процессы дифференциации ниш по осям сезонной и разногодичной ритмики условий, может менять сообщество до неузнаваемости, нередко превращая его в "хамелеона".
В условиях наземных сообществ возможны и суточные изменения, но они играют сравнительно небольшую роль и проявляются в основном в изменении архитектоники сообщества за счет поворотов листьев, соцветий и т. д. Весьма интересным приспособлением для сбора росы является, к примеру, "сон" листьев клевера, который на ночь складывает листочки таким образом, что более опушенная нижняя сторона листа оказывается эффективным аккумулятором росы. Е. В. Белоногов, изучая этот вопрос, даже сделал вывод о целесообразности внесения специальных подкормок в ночное время, которые могут затем через лист всасываться растениями. Значительно больше роль суточной ритмики в жизни водных сообществ. В литературе имеются многочисленные примеры перераспределения планктонных водорослей в ночное и дневное время. В ночное время они концентрируются у поверхности воды, а в дневное распределяются по всему профилю водной толщи, которая хорошо освещена солнечными лучами.
Для наземных сообществ основные формы динамики - сезонная и разногодичная. Вопрос этот многократно освещался в литературе, но наиболее полно был разработан Т. А. Работновым.
В составе растительных сообществ есть группы видов с разными циклами сезонного развития, что также отражает уже отмеченную дифференциацию ниш и является причиной многократного изменения облика сообществ в течение сезона. В особенности ярко дифференциация фитоценозов проявляется в лесах, где до распускания листьев деревьев напочвенный покров представлен эфемероидами - подснежниками, видами ветрениц, хохлаткой, гусиным луком, чистяком и пр. Эти виды успевают отцвести до распускания листьев древесных пород. Неодновременность зацветания характерна и для лугов и степей, где часто говорят о смене аспектов. Так, в знаменитом заповеднике "Стрелецкая степь", сохранившем остатки красочных северных луговых степей на черноземах европейской части СССР (мы говорили о их очень высоком альфа-разнообразии), поочередно зацветают сон- трава с фиолетовыми цветами, горицвет весенний - желтыми, шалфей луговой - синими, нивяник обыкновенный - белыми цветами и т. д. Таким образом, зацветая по очереди, ценопопуляции снижают конкуренцию и более равномерно используют ресурсы среды.
Фенологические изменения сообществ - очень важный признак, показывающий их адаптацию к климату и местным почвенным условиям. Обычно для отражения феноритмики сообщества используют две основные формы графиков - фенологические спектры и кривые цветения. Первую форму мы проиллюстрируем фенологическим спектром степного сообщества на гриве в пойме Средней Лены (пример заимствован из монографии К. Е. Кононова (1982), посвященной травяной растительности этой территории (рис. 10).
![Рис. 10. Феноспектр сезонного развития сообщества остепненного луга на гриве в пойме р. Лены (по: [Кононов, 1982]) Буквами обозначены фазы фенологического развития: а - вегетация; б - бутонизация; в - цветение; г - созревание плодов и семян; д - осыпание плодов и семян; е - отмирание. Виды: 1 - прострел желтеющий; 2 - незабудка душистая; 3 - кобрезия тонколистная; 4 - ветреница лесная; 5 - остролодочник колосовидный; 6 - подорожник серый; 7 - лапчатка прилистниковая; 8 - одуванчик рогатый; 9 - щавель пирамидальный; 10 - костер иркутский; 11 - горошек мышиный; 12 - лихнис сибирский; 13 - крестовник Якоба; 14 - эспарцет сибирский; 15 - мытник миловидный; 16 - астра альпийская; 17 - тысячелистник обыкновенный; 18 - ячмень короткоостистый; 19 - подмаренник настоящий; 20 - лук блестящий; 21 - герань луговая; 22 - гвоздика разноцветная; 23 - эдельвейс Палибина; 24 - смолевка ползучая; 25 - колокольчик скученный](pic/000010.jpg)
Рис. 10. Феноспектр сезонного развития сообщества остепненного луга на гриве в пойме р. Лены (по: [Кононов, 1982]) Буквами обозначены фазы фенологического развития: а - вегетация; б - бутонизация; в - цветение; г - созревание плодов и семян; д - осыпание плодов и семян; е - отмирание. Виды: 1 - прострел желтеющий; 2 - незабудка душистая; 3 - кобрезия тонколистная; 4 - ветреница лесная; 5 - остролодочник колосовидный; 6 - подорожник серый; 7 - лапчатка прилистниковая; 8 - одуванчик рогатый; 9 - щавель пирамидальный; 10 - костер иркутский; 11 - горошек мышиный; 12 - лихнис сибирский; 13 - крестовник Якоба; 14 - эспарцет сибирский; 15 - мытник миловидный; 16 - астра альпийская; 17 - тысячелистник обыкновенный; 18 - ячмень короткоостистый; 19 - подмаренник настоящий; 20 - лук блестящий; 21 - герань луговая; 22 - гвоздика разноцветная; 23 - эдельвейс Палибина; 24 - смолевка ползучая; 25 - колокольчик скученный
Лето в Якутии короткое, и даже в пойме р. Лены, где климат более мягкий, вегетационный период ограничен двумя с половиной месяцами. Первые растения зацветают в начале июня, и последние отцветают к концу июля. Последовательность зацветания очень постепенная, отсутствуют какие-либо выраженные группы, которые бы сменяли друг друга в цикле сезонного развития. Перед нами полный "фенологический континуум".
![Рис. 11. Кривые цветения трех типов луга в пойме Средней Лены (по: [Кононов, 1982]) 1 - остепненный луг; 2 - настоящий луг; 3 - болотистый луг](pic/000011.jpg)
Рис. 11. Кривые цветения трех типов луга в пойме Средней Лены (по: [Кононов, 1982]) 1 - остепненный луг; 2 - настоящий луг; 3 - болотистый луг
Вторую форму - кривую цветения, которая показывает количество цветущих видов в разное время вегетационного сезона,- мы проиллюстрируем тремя рисунками - 11, 12, 13. Рис. 11 и 12 заимствованы из той же монографии К. Е. Кононова и показывают, как влияет на фенологическое развитие положение участка в рельефе и особенности погодных условий конкретного года. Три типа луга: остепненный, настоящий и болотистый - связаны с местообитаниями различного гидротермического режима. Нетрудно видеть, что первым достигает фенологического типа остепненный луг хорошо прогреваемых и сухих местообитаний, а последним- болотистый, связанный с холодными переувлажненными почвами при близком уровне вечной мерзлоты.
![Рис. 12. Влияние различных погодных условий на феноритмику сообществ (по: [Кононов, 1982]) 1 - 1973 г.; 2 - 1972; 3 - 1971](pic/000012.jpg)
Рис. 12. Влияние различных погодных условий на феноритмику сообществ (по: [Кононов, 1982]) 1 - 1973 г.; 2 - 1972; 3 - 1971
То же сообщество остепненного луга, которое характеризовалось на рис. 10 и 11, имеет меняющуюся год от года ритмику. В год с более ранним паводком и теплым летом (1973) пик цветения наступает раньше (см. рис. 12).
На рис. 13 показано несколько совмещенных кривых цветения для разных типов растительности по работе В. Н. Голубева (1971). Этот исследователь по данным литературы и по собственным наблюдениям, которые он вел в разных районах нашей страны, проанализировал десятки кривых цветения.
![Рис. 13. Примеры кривых цветения сообществ разных типов растительности (по: [Голубев, 1971]) 1 - дубрава; 2 - луговая степь; 3 - полынно-ковыльная степь; 4 - тундра](pic/000013.jpg)
Рис. 13. Примеры кривых цветения сообществ разных типов растительности (по: [Голубев, 1971]) 1 - дубрава; 2 - луговая степь; 3 - полынно-ковыльная степь; 4 - тундра
На рисунке наглядно показано, что продолжительность периода вегетации сообщества, когда какие-то виды находятся в фазе цветения, зависит от климата; этот период для тундры много короче, чем для дубравы или луговой степи. Если вегетационный период сопровождается перерывами в поступлении осадков, то кривая цветения многовершинна - в периоды засухи число цветущих видов резко сокращается. Такая многовершинная кривая характерна для сезонного развития полупустынных сообществ и южных вариантов степей (кривая 3 характеризует ритмику цветения полынно-ковыльной степи).
Изучение сезонной ритмики не только интересный аспект теории, показывающий общую адаптацию сообществ и дифференциацию ниш, но и очень важный момент при организации сенокосного и пастбищного хозяйства. Известно, что накопление питательных веществ в растениях тесно связано с циклами сезонного развития. К моменту цветения в растениях отмечается наибольшее количество протеина, являющегося важнейшим элементом корма животных, а в период плодоношения происходит массовый отток питательных веществ из листьев в семена, листья и стебли грубеют и их питательная ценность понижается. Поэтому определение оптимального срока первого скашивания проводится с учетом сезонной ритмики преобладающих по массе растений. Соответственно, когда создается искусственный сенокос с несколькими компонентами из числа многолетних трав, составители травосмеси стараются включить в ее состав компоненты со сходными циклами сезонного развития, чтобы зацветание компонентов было более или менее одновременным. Тем самым, разумеется, усиливается конкуренция между компонентами за счет наложения их экологических ниш по оси сезонной ритмики, но это окупается более высоким качеством сена, в составе которого нет растений с уже огрубевшими стеблями.
Резервами для снижения конкуренции остаются другие оси, слагающие гиперпространство экологических ниш, и в первую очередь характер размещения подземных частей. В травостой включают, скажем, корневищный и рыхлокустовый злаки, а также бобовые с корневой системой кистекорневого типа. Однако требование одновременности зацветания всех компонентов делает сенокосные травосмеси потенциально менее богатыми видами, чем пастбищные. В последнем случае хозяйство заинтересовано в растянутости времени цветения и неоднородности зацветания компонентов, нужно чтобы нарастание зеленой массы происходило непрерывно на протяжении всего пастбищного сезона. Дифференциация ниш по оси сезонных изменений помогает цветоводам создавать декоративные ансамбли с многократным цветением без трудоемкой замены растений на клумбах в течение лета.
Разногодичные изменения сообщества (флюктуации) являются показателем его пластичности по отношению к колебанию различных факторов среды, определяющих характер сообщества. Т. А. Работнов подчеркивает, что в этом случае изменения касаются в основном соотношения компонентов сообщества и его общей продуктивности. Видовой и ценопопуляционный состав при этом остаются постоянными, хотя в отдельные годы некоторые элементы сообщества не развиваются и остаются в покоящемся состоянии. По причинам, вызывающим флюктуации, Т. А. Работнов делит их на экотопические, связанные с особенностями климата года, характером разлива рек, и антропогенные - колебания интенсивности использования, например влияние срока пастьбы при использовании пастбищ в системе пастбищеоборота, когда каждый год пастьба начинается с разных участков и после стравливания травостою предоставляется возможность восстановиться, а раз в несколько лет он полностью изолируется от выпаса для отдыха. Кроме того, различаются фитоциклические ("клеверные" годы, когда клевер массово цветет), фитопаразитарные (нападение нематод или других паразитов растений) и зооциклические флюктуации.
Очень интересные флюктуации последнего рода были описаны академиком Е. М. Лавренко и профессором А. А. Юнатовым в монгольских степях. В отдельные годы там массово размножается полевка Брандта и почти нацело поедает дернины ковылей. После этого из покоящихся почек на корневищах массово развивается вострец ложнопырейный и степь меняется до неузнаваемости. Однако спустя 4-5 лет ковыли вновь восстанавливаются, а вострец снова переходит в положение второстепенного вида. Таким образом, степная экосистема поддерживает устойчивость и не разрушается после повышения численности грызунов.
Разногодичные флюктуации выражены и в пустынях, где периодически в наиболее благоприятные периоды года дают вспышки "подпольщики"-однолетники. Уже упоминались данные Р. Уиттекера об однолетниках в пустыне Сонора, в их составе имеются "комплекты" для разных вариантов благоприятных периодов. Таким образом, для выявления всего потенциально возможного видового состава пустыни требуется срок по крайней мере в десять лет.
Однако наряду с пластичностью состава сообщества проявляются и антагонистические - по направленности механизма стабильности. Исследованием устойчивости травяных экосистем занимался К. А. Куркин, который показал, что два фактора поддержания продуктивности - увлажнение и содержание нитратного азота - антагонистичны. Во влажный год повышению урожая способствует оптимальность режима увлажнения почвы, а в сухой год дефицит влаги в некоторой степени компенсируется появлением значительно большего количества нитратного азота. Устойчивость флористического (ценопопуляционного) состава противоположна устойчивости продуктивности. Таким образом, в разные годы за счет развития разных видов также в известной степени погашаются климатические колебания.
Амплитуда колебаний продуктивности сообщества - весьма характерный признак его функции. Обычно чем меньше продуктивность сообщества зависит от климата (т. е. чем больше оно связано с питанием за счет грунтовых вод), тем амплитуда колебаний меньше. На рис. 14, составленном по данным Г. С. Сабардиной, показаны кривые изменения двух характерных сообществ для лесной зоны европейской части СССР, и нетрудно видеть, что постоянно связанный с грунтовыми водами черноосочник в меньшей степени зависит от колебаний климата, и потому его урожайность меняется от года к году довольно незначительно. Суходольный луг, напротив, связан с сухими почвами, которые увлажняются в основном только за счет атмосферных осадков. По этой причине его урожайность следует за количеством осадков в вегетационный период и в разные по климату годы сильно меняется.
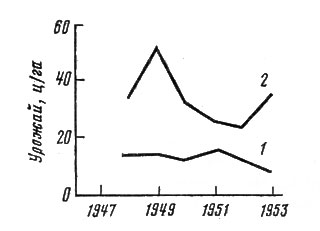
Рис. 14. Разногодичные колебания урожайности двух луговых сообществ лесной зоны (по: Сабардина, 1955). 1 – черноосочник; 2 - суходольный луг
Таким образом, колебания климата в разные годы бывают столь велики, что далеко не всегда стабилизирующие механизмы могут обеспечить высокий выход фитомассы. По этой причине нарастание стволов деревьев происходит с неодинаковой скоростью год от года и разным оказывается урожай сенокосов или пастбищ. В этом случае фактором стабилизации может оказаться и человек, который, организуя полив и удобрение, часто делает продуктивность сообществ вполне автономной по отношению к погодным условиям.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'