Глава 6. Какие сообщества выгоднее природе? (Видовое разнообразие)
Обсуждая вопрос о гиперпространстве ниш, мы подчеркивали, что, оно, с одной стороны, зависит от комплекса чисто физических абиотических условий среды: количества осадков и ритма их поступления, обеспеченности почвы минеральными веществами, температурного режима,- а с другой - от свойств слагающих сообщество ценопопуляций растений. При появлении нового толеранта, который способен использовать ресурсы в тех условиях, когда прочие виды сообщества этого сделать не могут самого засухоустойчивого, холодостойкого, теневыносливого растения), увеличивается объем гиперпространства ниш. Это положение, кстати, ныне все шире используют рекультиваторы промышленных отвалов - вынесенных на поверхность пустых пород из шахт, скоплении шлаков и т. д. Наряду с чисто физическим увеличением гиперобъема ниш благодаря покрытию этих неуютных для растений субстратов слоем плодородной земли, изменения реакции их раствора из-за соответствующей химической обработки (нейтрализации) используют растения, наделенные высоким свойством S (это виды пустынь, солончаков и т. д.). Такие растения могут поселиться непосредственно на промышленном отвале без его обработки и начать процесс его биологического преобразования.
Таким образом, при первом, поверхностном ознакомлении с положениями концепции экологических ниш можно сделать скоропалительный вывод о том, что, чем гиперпространство ниш больше, т. е. чем условия благоприятнее, тем выше должно быть видовое богатство сообщества, или как его называют, альфа-разнообразие. В этом случае мы должны бы были выявить линейную зависимость между продуктивностью и биомассой сообщества и числом видов-продуцентов и сделать вывод, что в любом случае для природы выгоднее сообщества с максимальным количеством видов. Однако это далеко не так, поскольку вопрос о том, какое количество цепопопуляцнй делит ресурсы, зависит еще и от того, какова стратегия видов, собравшихся в сообществе. Если в сообществе имеется очень мощный виолент (точнее, если сложились условия, что у одного из видов может проявиться сильная виолентность), то он может захватить "львиную долю" ресурсов, и в этом случае альфа-разнообразие будет очень низким. В речных поймах самыми богатыми минеральными веществами и оптимальными по режиму увлажнения являются так называемые слоистые почвы вдоль русла, где ежегодно выпадает обильный плодородный наилок и паводки хорошо увлажняют почвы. Однако в этих условиях сплошь и рядом сообщества крайне обеднены видами, а абсолютное доминирование получает костер безостый, который очень хорошо специализирован для жизни на таких периодически перекрываемых свежими наносами субстратах. В этих условиях лежит его синэкологический оптимум, и он активно подавляет конкурентов. Аналогично подавляет конкурентов и пырей ползучий, который также относится к числу корневищных растений и часто на залежи в первые годы после прекращения использования пашни формирует сообщество с абсолютным доминированием.
Как ни парадоксально, но если в случае такого монодоминирования несколько ухудшить условия, введя какой-то новый фактор, например умеренный выпас, то видовое разнообразие резко возрастает, как возрастает оно и в лесах, когда на них усиливается влияние человека, использующего лес как место отдыха (рекреационные нагрузки). Уменьшение гиперобъема ниши доминанта-виолента в этих случаях происходит за счет ликвидации условий его синэкологического оптимума. Таким образом, при прочих равных абиотических условиях видовое разнообразие в сообществе тем выше, чем равномернее виолентность слагающих сообщество ценопопуляций. Этот фактор учитывается и при создании травосмесей, где поддержание стабильности видового состава возможно только при включении в травосмесь трав примерно сходной виолентности и обеспечении некоторого ценотического баланса и примерно равного раздела гиперпространства ниш.
Р. Уиттекер, много занимавшийся проблемой видового разнообразия, подчеркивал, что в значительной мере оно является функцией "эволюционного времени", и, сопоставляя сходную растительность территорий, которые подвергались и не подвергались оледенению, приводил убедительные примеры большего богатства видами последней. Таким образом, кроме чисто физической величины гиперпространства ниш (экологического объема местообитания) сообщества и стратегического спектра слагающих его популяций, на видовое разнообразие оказывает влияние также абсолютный возраст сообщества, т. е. длительность его пребывания в ненарушенном состоянии. Связь этих трех параметров неопределенная, они могут усиливать или ослаблять друг друга, и потому Уиттекер делал вывод о том, что альфа-разнообразие относится к числу наиболее труднопрогнозируемых признаков сообщества.
Тем не менее если отвлечься от частностей и рассматривать общие тенденции изменения видового разнообразия в глобальном масштабе, то можно говорить о некоем главном градиенте разнообразия. Уиттекер определял его как изменения сообществ от высоких широт Арктики к тропикам на равнине и от высокогорий к равнинам. Самые богатые видами сообщества - тропические леса, саванны и самые бедные - сообщества альпийских и арктических пустынь.
Понятно, что коррективы к градиенту на равнине нужно внести при учете континентальности района, т. е. его удаления от океана, и соответственно количества осадков и характера изменения температуры в годичном цикле. Тепло без влаги, как и влага без тепла, не может служить источником улучшения условий и увеличения физического гиперпространства ресурсов, а стало быть, и альфа-разнообразия. По этой причине в низких широтах, если это пустыня, альфа-разнообразие также будет низким. Аналогично и в горах. Градиент увеличения видового разнообразия будет наблюдаться только в том случае, если в районе, где расположена горная система, соотношение тепла и влаги оптимально, т. е. если это район влажных тропиков или субтропиков. Если же, скажем, горная система расположена в пустыне, то изменение видового разнообразия не будет линейным, вначале при подъеме в горы оно будет возрастать (пустыня сменится степью или саванной), а затем пойдет на убыль. Таким образом, высказанные Уиттекером положения о главном градиенте разнообразия должны восприниматься с осторожностью.
Для измерения альфа-разнообразия предложено много различных показателей, большинство из которых учитывает не только количество видов, но еще и соотношение их значимости в сообществе, т. е. степень количественной представленности. Тем не менее число видов на единицу площади является самым простым и самым надежным показателем. Возникает лишь достаточно сложный вопрос о том, на какой площади измерять это количество видов, так как с увеличением ее размера количество видов естественно возрастает. В целом фитоценологи приходят к выводу, что по мере увеличения размера пробной площади вначале количество видов увеличивается очень быстро, а затем эта кривая начинает постепенно затухать (рис. 7). Размер площадки, соответствующий этой точке перегиба кривой, определяемой обычно чисто эмпирически, называют минимальным ареалом сообщества. Нетрудно видеть, что в нашем примере нарастание полноты выявления флористического состава более быстро происходит в сообществах, богатых видами (луговая степь), чем в бедных (пустыни). Соответственно целесообразно при описании растительности в условиях луговой степи пользоваться площадкой меньшего размера, а пустыни - большего.
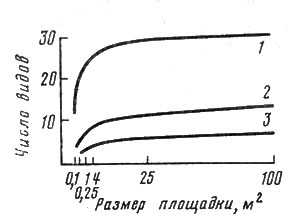
Рис. 7. Изменение числа видов с увеличением размера учетной единицы для трех типов сообществ Монголии. 1 - луговая степь с овсяницей ленской и разнотравьем; 2 - пустынная степь с змеевкой джунгарской и ковылем; 3 - пустыня с реомюрией джунгарской
Пробные площадки размера минимального ареала рекомендуется использовать для характеристики растительности. Они вполне представляют то сообщество, которое исследователь описывает, выявляя большинство видов сообщества, и позволяют оценивать количественную представленность каждого из них. Обычно фитоценолог пользуется условной шкалой учета роли видов с небольшим числом градаций. Точный учет количества вида (скажем, подсчетом числа особей или определением их массы) при такого рода описаниях обычно нецелесообразен, так как требует нереально больших затрат времени. Затратив его на один участок растительности, исследователь попросту не успеет описать еще другой, третий и т. д. А массовость исходного материала совершенно необходима для последующего выделения типов сообществ или ординации, при которых производят усреднение данных по каждому типу (классификация) или отрезку градиента (ординация).
Количество видов на такой соответствующей "минимальному ареалу" площадке может меняться в очень широких пределах, а сам размер пробной площадки может составлять от четырех (обычно травяные сообщества, луга, степи) до сотен квадратных метров (леса). В растительности нашей страны самые флористически богатые сообщества - луговые (северные) степи, где на учетной единице можно обнаружить свыше 100 видов. Все три фактора повышения разнообразия: объем гиперпространства ниш, отсутствие виолента и достаточное "эволюционное время" - действуют на эти ныне уже заповедные участки курских степей однонаправленно и параллельно. В таких сообществах очень ярко выражены дифференциация ниш вдоль оси сезонного развития, разногодичные колебания видового состава, дифференциация глубины проникновения корней и т. д. Самые бедные сообщества - это группировки растений вдоль побережий озер, где нередко распространены чистые заросли рогоза, тростника, камыша и т. д. На наших пашнях можно встретить 10-20 видов сорных растений (причем на видовую насыщенность культура агротехники может не влиять, виды могут быть представлены единичными особями, которые практически не снижают урожайности культурного растения), количество видов на лугах колеблется в пределах от 20 до 50-60. В широколиственных лесах благодаря влиянию доминанта-виолента видовом список оказывается меньшим, обычно он ограничен 20-70 видами, хотя существуют еще и мертвопокровные леса, где вследствие сильного затенения напочвенный покров почти не развит, и в этом случае число видов может падать ниже 10. Рекордсмены альфа-разнообразия - тропические леса насчитывают 1000 и более видов, причем преимущественно деревьев и лиан. Вообще, если просматривать изменение видового разнообразия лесов с севера на юг отдельно для деревьев и для напочвенного яруса, то картина получается весьма любопытной: количество видов напочвенного покрова падает, а число видов деревьев возрастает.
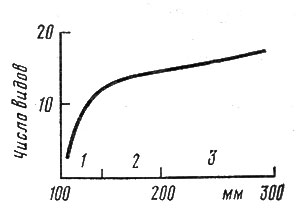
Рис. 8. Изменение видовой насыщенности (альфа-разнообразия) сообществ на градиенте пустыня - степь в Центральной Монголии. Рисунок повторяет тот же градиент, который показан на рис. 3. Указано среднее число видов на 100 м2: 1 - пустыня; 2 - опустыненная степь; 3 - сухая степь
На рис. 8 показаны результаты статистического анализа изменения числа видов в сообществах аридной полосы Монголии - от пустынь к сухим степям. При усреднении данных большого количества описаний площадок размером 100 м2 (1500) картина получилась четкой и вылилась положительная зависимость между количеством осадков и числом видов.
Альфа-разнообразие является лишь одной из форм видового разнообразия растительности. Следующей формой является бета-разнообразие, т. е. разнообразие сообществ. Для его оценки можно использовать результаты классификации (см. гл. 13) и оценить бета-разнообразие числом установленных типов сообществ. Если в одном районе установлено ассоциаций меньше, а в другом - больше, то соответственно бета-разиообразие второго выше. Чаще же для измерения бета-разнообразия пользуются показателем, который называется "полусмен" и обозначается буквами НС (от английского half change).
Для измерения бета-разнообразия в полусменах проводят ординацию сообществ вдоль градиентов ведущих факторов и для каждого градиента устанавливают свою меру бета-разнообразия. Скажем, выполнив ординацию по оси увлажнения, смотрят, как меняется видовой состав от влажного полюса градиента к сухому. Естественно, что постепенно происходит выпадение одних видов и появление других. Как только произойдет смена половины состава, то разнообразие этой части градиента оценивается в 1 НС, когда состав растительности обновится целиком, разнообразие достигнет 2 НС и начнется новая смена состава сообщества. Разнообразие всего изученного градиента может составить 3, 4 и даже 5 или 6 НС.
В Центральной Якутии широко распространены аласы - безлесные котловины в тайге, где благодаря аккумуляции солей развиваются сообщества солончаковых лугов. В центре аласа обычно озеро, по верхней части - степь. Если просматривать изменение бета-разнообразия от прибрежно-водных группировок до степи, то список видов сменится два раза - вначале на смену прибрежно-водным растениям, таким, как манник, тростник и осоки, придут настоящие луговые - ячмень короткоостый, пырей, бескильница и др., а затем они заменятся на степной набор - типчак, ковыль тырсу, ломкоколосник и др. Таким образом, бета-разнообразие аласа в Центральной Якутии по градиенту увлажнения оценивается в 4 НС. Показать полностью смену флористического состава на этом градиенте - дело довольно сложное (и вряд ли это интересно читателям). На рис. 9 показана смена лишь некоторых видов, наиболее характерных для разных типов растительности якутских аласов.
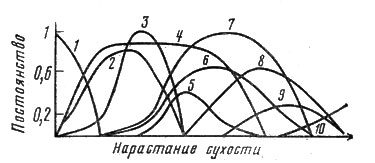
Рис. 9. Пример экологического ряда высокого бета-разнообразия на аласах Центральной Якутии. 1 - ломкоколосник ситниковидный; 2 - типчак ленский; 3 - прострел желтеющий; 4 - полынь замещающая; 5 - гвоздика разноцветная; 6 - ячмень короткоостый; 7 - бескильница тонкоцветковая; 8 - лисохвост тростниковидный; 9 - бекмания восточная; 10 - крестовник арктический
Наконец, все флористическое богатство района называется гамма-разнообразием. Оно слагается из альфа-разнообразия и бета-разнообразия. Практически это произведение среднего числа видов в сообществе на число типов сообществ, и потому одно и то же гамма-разнообразие может быть достигнуто разным путем: может быть сравнительно немного сообществ, богатых видами, и много сообществ, бедных видами. Таким образом, видовое разнообразие района является следствием "упаковок" двух порядков - ниш ценопопуляций и местообитаний популяций - и вклад этих "упаковок" в разных природных районах различен, хотя в целом, альфа- и бета-разнообразие меняются более или менее параллельно. В экстремальных условиях (пустыни) снижается альфа-разнообразие и климат "давит", нивелируя различия склонов разной экспозиции при невысоких превышениях, различия содержания в почве солей и т. д. При более благоприятных условиях, скажем лесостепной зоне, когда альфа-разнообразие возрастает, растительность наиболее чутко реагирует на малейшие нюансы рельефа, эдафических факторов и т. д.
Совокупность видов растений одного района называют также конкретной флорой, которая практически является синонимом гамма-разнообразия, хотя выявляющий конкретную флору ботанико-географ преследует иные цели и обычно изучает ее состав не экологически или синтаксономически, а, как систематик, выявляет роль различных семейств, исследует ареалы входящих в конкретную флору видов и т. д.
Таким образом, на поставленный в заголовке главы вопрос о том, какие сообщества выгоднее для природы (многовидовые или маловидовые), нельзя дать однозначный ответ. Все зависит от особенностей условий среды и собравшихся в сообществе видов. Тем не менее общей тенденцией эволюции растительности является, безусловно, повышение плотности упаковки ценопопуляций в сообществе и популяций в растительности. Не признав этого, мы не сможем объяснить возможности сосуществования на нашей планете того гигантского количества видов растений, которые создала природа.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'