Глава 5. Почему растет кермек? (Типы местообитаний)
Если ценопопуляция внутри сообщества имеет нишу, то популяция в ландшафте - местообитание, т. е. определенный гиперобъем в многомерном гиперпространстве, осями которого являются факторы среды ландшафта. Каждый вид имеет в условиях ландшафта свой диапазон распределения (пределы толерантности) по каждому из факторов среды, и потому, зная пределы вариации условий среды, можно прогнозировать, какие виды могут входить в состав растительности. Таким образом, появление, скажем, красавца кермека с жесткими листьями и изящным фиолетовым соцветием свидетельствует о том, что в гиперпространстве местообитаний ландшафта есть сухие солончаковые местообитания.
Применительно к гиперпространству местообитании и популяциям могут быть повторены все те же логические конструкции, которые были использованы нами при рассмотрении ниш и ценопопуляций, как-то: дифференциация местообитаний и упаковка популяций, В конечном итоге ценопопуляция - понятие, как мы отмечали, условное и рабочее, так как определить, где кончается одно сообщество и соответствующая ему ценопопуляция и начинается другое, просто невозможно. Однако гиперпространство местообитаний несравненно шире, чем гиперпространство ниш. и потому проследить соотношение местообитаний отдельных популяций много проще, чем ниш в сообществе. Есть в этом случае и ситуация, аналогичная фундаментальной и реализованной нише, так как вид занимает не то местообитание, где он в принципе мог расти, а распространен там. где он наиболее конкурентоспособен. Центры этих распределений вида по осям условий среды носят название экологического и синэкологического оптимумов.
Этот интереснейший феномен природы неоднократно был описан в работах А. И. Прозоровского, А. П. Шенникова, X. Элленберга. В частности, было показано, что степные виды при отсутствии конкуренции значительно лучше растут в условиях более обильного лугового увлажнения и потому их правильнее называть не "сухолюбами", а "сухотерпцами". Аналогично лучше развиваются на богатых минеральных почвах вересковые кустарнички, подобные багульнику или Кассандре, которые конкуренция вытолкнула на наиболее бедные местообитания сфагновых болот. В этих условиях кустарнички оказываются в выигрышном положении как конкуренты, так как обладают наиболее соответствующей этим условиям "скаредной экономикой" патентов.
Для изучения отношения видов к факторам местообитаний используются специальные методы, которые называются ординационными.
Первые методы ординации предложил Л. Г. Раменский, причем сделал это почти одновременно с формулировкой положений о непрерывности. В этом он намного опередил Г. Глизона, который, утверждая идеи континуума и сокрушая теорию "единиц растительности", не предлагал никаких позитивных приемов обработки данных вместо классификации, которую использовали его оппоненты.
Со времени Раменского ординационные подходы "пышно расцвели", так как тонко соответствовали непрерывной природе растительности, что позволяло широко использовать математические методы и ЭВМ. Поскольку растительность отражает условия среды, то удалось разработать и ряд методов так называемой непрямой ординации, когда для изучения отношения видов к условиям среды и соответственно характеристике их местообитаний... не было необходимости вести прямые наблюдения факторов среды. Это было очень удобно, так как некоторые факторы, например такие, как влажность почвы, измерить трудно: нужно многие годы вести режимные наблюдения на стационарах, чтобы описать, как меняется влажность почвы в разные сезоны одного года и в разные по особенностям климата годы.
В основе методов непрямой ординации лежит положение об отражении составом сообществ условий среды. Стало быть, два наиболее непохожих сообщества имеют наиболее различные типы местообитаний, а два наиболее сходных по своему составу - наиболее близкие. Если ординируются не сообщества, а виды, то в этом случае сходный характер распределения видов по различным описаниям опять-таки будет интерпретирован как отражение близости их экологии, а несходный - их экологических различий. Возможности современных ЭВМ позволяют самым различным образом анализировать сходство и несходство видового состава описаний или распределения видов по различным описаниям (эти методы получили название методов R и Q), однако их рассмотрение лежит за рамками содержания этой популярной книги.
Приведем примеры результатов ординационного анализа растительности прямыми методами, в частности, прямым градиентным анализом. При этом методе для каждого описания исследователь получает оценку условий среды, далее упорядочивает описания по ряду нарастания фактора и характеризует либо изменение видового состава растительности через анализ сходства описаний (Q-метод), либо изменения постоянства отдельных видов (R-метод). На рис. 3 показаны результаты градиентного анализа изменения растительности на профиле протяженностью 350 км, который пересекал зону пустынь и степей Монголии и был заложен автором совместно с советскими и монгольскими коллегами. В данном случае экологическим фактором, который принят в качестве градиента, было расстояние от начала профиля, так как при движении на север увеличивается количество осадков. Такие схемы носят название топоклинов (от топо - положение, клин - постепенное изменение).

Рис. 3. Континуум на границе пустынь и степей Монголии. Встречаемость видов: а - 5-49%; б - 50-79%; в - 80-100%; г - каждое деление соответствует 25 км. Показано также изменение среднегодового количества осадков в мм Виды: 1 - драгантемум гобийский; 2 - солянка воробьиная; 3 - парнолистник желтодревесный; 4 - парнолистник Розова; 5 - анабазис коротколистный; 6 - реомюрия джунгарская; 7 - потаниния монгольская; 8 - змеевка джунгарская; 9 - ковыль гобийский; 10 - ирис Бунге; 11 - лук монгольский; 12 - бассия шерстолистная; 13 - лагохилюс иглолистный; 14 - лук многокорневой; 15 - птилотрихум сизый; 16 - крашенинниковия; 17 - гармала чернушковая; 18 - вьюнок Аммана; 19 - чий блестящий; 20 - цельнолистник даурский; 21 - карагаиа желтокорая; 22 - змеевка растопыренная; 23 - ковыль Крылова; 24 - полынь холодная; 25 - доктостемон цельнолистный; 26 - гетеропагшус шерстинистоволосистый; 27 - осочка твердоватая; 28 - карагана карликовая; 29 - хамеродос прямой; 30 - серпуха васильковая; 31 - сибальдианта сплюснутая; 32 - лапчатка двувильчатая; 33 - пырей гребневидный; 34 - полынь Сиверса; 35 - элисум китайский; 36 - володушка двустебельная; 37 - тонконог гребенчатый; 38 - цимбария даурская; 39 - мятлик вытянутый; 40 - карагана мелколистная
Следует обратить внимание на отсутствие каких-либо резких границ между зонами пустынь и степей и подзонами, которые также отражают изменения климата. Границы зон и подзон по этой причине проводятся с большим или меньшим элементом условности в результате коллективной договоренности. Всегда есть основания для споров о смещении границ на юг или на север на 50, а иногда даже на 75 или 100 км.
![Рис. 4. Пример континуума растительности дельты Волги вдоль оси концентрации солей в почве (по: [Голуб]). 1 - частуха подорожниковая; 2 - молочай Бородина; 3 - канареечник тростниковидный; 4 - алтей лекарственный; 5 - ажрек; 6 - аргузия сибирская; 7 - клубнекамыш морской; 8 - триполиум обыкновенный](pic/000004.jpg)
Рис. 4. Пример континуума растительности дельты Волги вдоль оси концентрации солей в почве (по: [Голуб]). 1 - частуха подорожниковая; 2 - молочай Бородина; 3 - канареечник тростниковидный; 4 - алтей лекарственный; 5 - ажрек; 6 - аргузия сибирская; 7 - клубнекамыш морской; 8 - триполиум обыкновенный
На рис. 4 и 5 показаны результаты градиентного анализа растительности дельты р. Волги по отношению к двум главным акологическим факторам этой территории - засолению и увлажнению. Рисунки наглядно показывают, что виды, подобные канареечнику тростниковидному и частухе подорожниковой, снижают свою встречаемость при повышении концентрации солей в почве. Триполиум, напротив, достигает максимума встречаемости при самом высоком засолении. Остальные виды имеют оптимум в средней части градиента. Рисунок хорошо иллюстрирует принцип континуума: распределения всех видов индивидуальны.
![Рис. 5. Распределение некоторых видов дельты р. Волги по градиенту относительного превышения (по: [Голуб]). 1 - ежеголовник прямой; 2 - канареечник тростниковидный; 3 - риция; 4 - бодяк полевой; 5 - манник тростниковидный; 6 - молочай Бородина; 7 - алтей лекарственный; 8 - ажрек; 9 - свинорой пальчатый; 10 - мортук пшеничный; 11 - солерос европейский; 12 - клубнекамыш морской](pic/000005.jpg)
Рис. 5. Распределение некоторых видов дельты р. Волги по градиенту относительного превышения (по: [Голуб]). 1 - ежеголовник прямой; 2 - канареечник тростниковидный; 3 - риция; 4 - бодяк полевой; 5 - манник тростниковидный; 6 - молочай Бородина; 7 - алтей лекарственный; 8 - ажрек; 9 - свинорой пальчатый; 10 - мортук пшеничный; 11 - солерос европейский; 12 - клубнекамыш морской
Аналогична картина изменения характера растительности в связи с высотой над меженью (т. е. средним летним уровнем) реки. Относительное превышение коррелирует с увлажнением, причем поскольку климат в низовьях Волги соответствует полупустыне, то на интервале всего трех метров высоты происходит смена видов от типичных прибрежно-водных (ежеголовник, канареечник) через виды влажных солончаковатых лугов (клубнекамыш морской, алтей лекарственный) до растений полупустыни (ажрек).
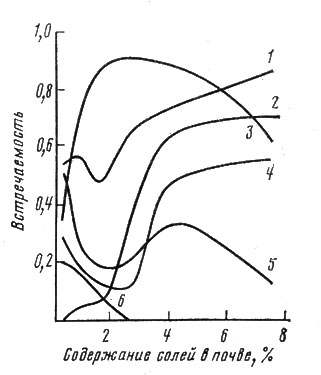
Рис. 6. Распределение некоторых видов поймы р. Амударьи по градиенту засоления. 1 - тамарикс удлиненный; 2 - карабарак; 3 - туранга; 4 - ажрек; 5 - солодка гладкая; 6 - вейник сомнительный
На рис. 6 приведены результаты градиентного анализа отношения некоторых видов к засолению в условиях поймы р. Амударьи. Проанализированная растительность представляет собой прирусловые ивняки и тугайные леса. Для светолюбивых видов характерен провал в распределении многих видов при средней концентрации солей в почве, что соответствует наиболее полному развитию тополя туранги, который перехватывает ресурсы света. В этом проявилась хорошо известная экологам двувершинность распределения конкурентно-слабых видов, которые на части диапазона снижают свое участие в растительности за счет того, что там развивается более мощный конкурент.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'