17.9. Синдромы опыления у орхидных
Изучение опыления у орхидных представляет собой целую науку. В этой книге невозможно описать все безграничное число вариаций, свойственных этому семейству. Мы представим лишь три главных типа опыления, чтобы показать основные пути их функционирования. Подробности можно найти в работе ван дер Пэйла и Додсона (van der Pijl, Dodson, 1966).
Cypripedium calceolus L. (рис. 50)
Северное полушарие.
Гомогамные цветки-полуловушки. Обманная аттрактация.
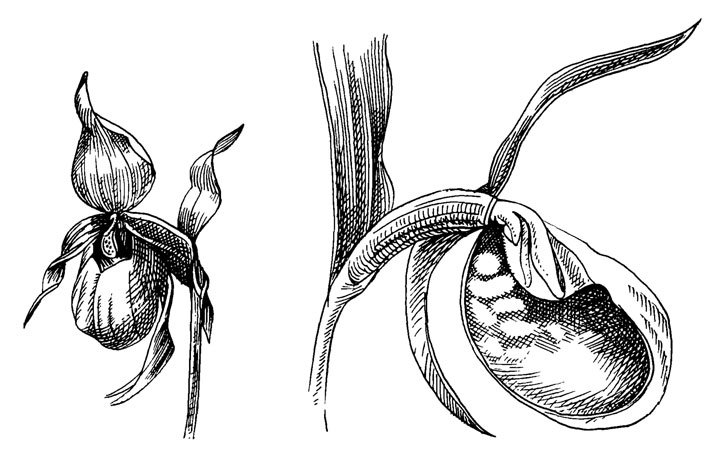
Рис. 50. Cypripedium calceolus L
У этого вида пять лепестков околоцветника настолько малы и тусклы, что они почти не имеют значения в качестве визуального аттрактанта в синдроме опыления. Напротив, губа большая блестящая желтая, похожая на мешок. У других видов губа часто менее выделяющаяся, а другие лепестки более заметны. Запах исходит от боковых лепестков (Stoutamire, 1967).
Центрипетальная часть губы сужается до канала, над которым гиностемий образует крышечку, оканчивающуюся щитовидным стаминодием, закрывающим наиболее оперативную часть. Рыльце образует другую щитовидную структуру с воспринимающей поверхностью, повернутой книзу. Два функциональных пыльника открыты - по одному на каждой стороне базальной части рыльца. Внешняя часть канала закрыта, но гиностемий сжат центрипетально, а в основании остаются выходные отверстия - по одному на каждой стороне.
Долгое время обсуждался вопрос об аттрактанте цветка Cypripedium. Вероятно, этот цветок не предоставляет опылителям никакой пищи. Полагают, что мерцание волосков (если эти наблюдения верны) может быть интерпретировано как "проявление реакции" заманивания насекомых в ловушку. По мнению Дауманна (Daumann, 1968), аттрактация происходит с помощью обмана (запаха), и опылители - мелкие одиночные пчелы рода Andrena влезают в полуловушку-губу, из которой только один выход - вдоль гиностемия. По-видимому, механизм опыления у других видов такой же [например, С. debile имитирует запах грибов (Stoutamire, 1967)].
Небольшое пространство внутри губы не позволяет насекомым летать, а внутренние поверхности настолько скользкие, что по ним нельзя выбраться из ловушки (Daumann, 1968). Итак, по крайней мере у этого вида нет необходимости, чтобы завернутый внутрь край губы функционировал в качестве барьера, препятствующего любому насекомому выбраться из цветка. Таким образом, опылитель должен протискиваться через каналы под гиностемием. Делая это, он не может не касаться спиной сначала рыльца, а затем одного из пыльников. Следовательно, имеет место функциональная протогиния. Пыльца сыпучая, но липкая и прилипает к спине опылителя.
Губа С. calceolus настолько сильно изгибается, что насекомое вряд ли видит выход, находясь на дне цветка. Но в изгибе имеется несколько светлых окон, которые будут привлекать насекомых, находящих, таким образом, позднее настоящий выход. У Сурripediinae с прямой губой (Paphiopedilum) выходы непосредственно видны со дна губы, и окон нет. В этой группе края губы прямые или даже отогнуты наружу.
Orchis maculata и родственные виды (рис. 51)
Европа.
Гомогамный. Мелиттофильный. Открытый цветок с зевом. Аттрактант не известен.
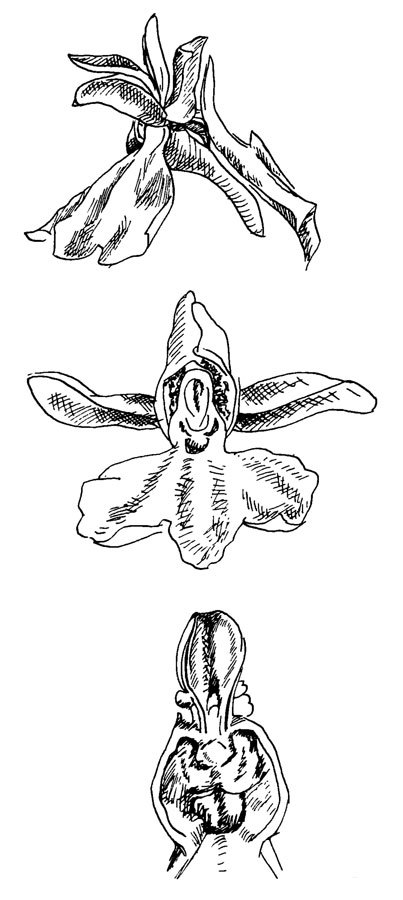
Рис. 51. Orchis maculata
Действующие части цветка - губа и гиностемий. Пять других лепестков образуют только часть аттрактивного аппарата. Опылители - главным образом пчелы и иногда мухи - садятся на губу так что их ось параллельна оси цветка, и ориентируются в направлении центра цветка, полости рыльца и входа в шпорец, расположенного в основании губы.
Один оставшийся пыльник образует верхушку гиностемия. Он имеет сравнительно широкий связник; две теки открываются щелями, причем их содержимое образует две сцементированные массы - поллинии. Поллиний прбдолжается в тонкую ножку. направленную к носику.
Из всех органов в цветке орхидных носик - самый спорный, и о его морфологическом происхождении не высказано никаких окончательных мнений. Он интерпретировался и как измененная лопасть рыльца, и как измененная тычинка, и как орган, возникший de novo. Поскольку рыльце у многих орхидных трехлопастное, это наводит на мысль, что у орхидных все три лопасти рыльца все еще сохраняют свою функцию.
На самом деле носик представляет собой выступающий бугорок, внутренние клетки которого частично дегенерируют и образуют два липких шарика, окруженных вязкой жидкостью. Ножка прилипает к эпидермису верхней поверхности носика. Легко разрываясь, он оставляет два очень маленьких диска эпидермиса, прикрепленных к ножкам. Липкие шарики прикреплены к нижней стороне дисков.
Затронутый насекомым, проникающим в цветок (это можно имитировать острым карандашом или другим удобным предметом), эпидермис носика разрывается, как описано выше, и остаток отгибается назад, выставляя вязкие шарики, которые немедленно прикрепляются к поверхности вторгшегося опылителя. Качество склеивающих веществ великолепно: они так прочно прикрепляют диски на животных, что те, улетая, вытаскивают целый поллинарий (прилипальце, ножку и поллиний) из пыльника.
Первоначально поллинарий выступает прямо и обычно в этом положении ударяет пыльник следующего посещаемого цветка. Вследствие несимметричного высыхания клеток, ножка в течение полминуты изгибается вперед и поллиний принимает такое положение, при котором он ударяет рыльце следующего цветка. Поверхность рыльца очень липкая, и большая часть, иногда все поллинии прилипают, оставляя на теле насекомого только ножку и диски как следы успешной опылительной активности.
Карман в нижней части носика немедленно отключается и с вторгшимся объектом больше нет контакта. Таким образом, чтобы защитить прилипальце оставшихся поллинариев - как это часто случается, - только один из них должен быть удален первым опылителем.
У всех базитонических орхидных имеются незначительные изменения, связанные главным образом с расстоянием между прилипальцами и местом их слияния в один орган, соответствующим расстоянию между двумя боковыми поверхностями органа насекомого, к которому прилипает поллинарий, от тонкого хоботка до широких частей тела, а также от верхушки до бокового положения и дальше до положения под хоботком, как в неперевернутых цветках Nigritella. У большинства видов аттрактантом является нектар, у Ophrys аттрактация половая. Нектар собирается в шпорец, длина которого варьирует в соответствии с размером опылителя, предпочитающего этот цветок. У Ophrys нет шпорца.
Orchis представляет собой другое исключение. Его аттрактант неизвестен; нектара не имеется в шпорцах многих видов (Daumann, 1941, 1971).
Во многих случаях род Orchis и родственные ему роды не стабильны. Сомнительно, являются ли шмели основными опылителями данной группы. Известно, что дневными и ночными бабочками опыляются различные виды, даже внутри самого рода Orchis (О. ustulata), а на Фарерских островах, где род Bombus не участвует в опылении, Хагерупом было показано, что некоторые орхидные опыляются мухами (Eristalis).
Виды Cattleya (рис. 52)
Тропическая Южная и Центральная Америка.
Гомогамный. Мелиттофильный. Нектарная аттрактация.

Рис. 52. Cattleya
Cattleya является хорошим представителем группы акротонических орхидных, которые отличаются от базитонических (например, Orchis) расположением пыльника и деталями механизма удаления поллиниев.
Как и у Orchis, к оперативным частям цветка относятся губа и гиностемий, которые в этом случае образуют единицу аттрактации с зевом внутри цветка. Остальные лепестки (некоторые из них очень заметны) образуют только органы аттрактации. Для приведенного здесь типа цветка (у отдельных видов Cattleya тип цветка отклоняется от обычного) адаптированными опылителями являются крупные полуобщественные пчелы. Они садятся на губу и ориентируются по направлению к центру цветка. Нектар находится в нектарнике, расположенном в завязи.
Пыльник не прямой, как у Orchis, а изогнут, причем его главная ось перпендикулярна оси гиностемия. Его дорсальная половина более или менее погружена в ткань гиностемия (морфологическое значение которого мы обсуждать здесь не будем). Имеются ножки, которые слегка выступают из пыльника, но ни к чему не прикреплены. В передней части теки щелей нет.
Носик образует широкую губу, верхняя часть которой состоит из прочной ткани с интактным эпидермисом, тогда как нижняя часть дегенерирует с образованием вязкого материала, свободно выступающего под носиком.
Влезая в цветок, насекомое не будет контактировать с этой вязкой массой, так как верхняя жесткая часть носика отогнута назад и скрывает вязкое вещество, защищая его. Однако, вылезая из цветка, насекомое вначале выгнет эту жесткую часть носика таким образом, что выступающая нижняя часть и "неожиданное количество вязкого материала оттягивает края в стороны и в то же самое время втягивает кончик пыльника" (Дарвин). Эффект состоит в том, чтобы приклеить ножки к насекомому. Когда поллиний удаляется из пыльника, верхняя часть его должна подняться подобно крышке, и у многих видов пыльник разрушается.
Дальнейшие стадии, высыхание ножек, положение поллиниев по отношению к полости рыльца и т. д. в принципе аналогичны соответствующим чертам в опылении Orchis.
Самые важные операциональные различия по сравнению с Orchis следующие: 1) эффект крышки пыльника и 2) работа механизма при выходе из цветка. Если влезающее в цветок насекомое уже имело бы на спине приклеенные поллинии, то они оседали бы в полости рыльца перед удалением пыльника. Общий эффект должен заключаться в функциональной протогинии, обеспечивающей аллогамию.
У акротонических орхидных вариации касаются главным образом числа, формы и связывания поллиниев, а также формы и прикрепления ножек.
У Listera ovata ножки нет, и поллиний непосредственно приклеивается к насекомому. В этом цветке вязкое вещество закупорено и извергается из носика только при прикосновении. Затем оно в течение нескольких секунд высыхает. Поскольку ножка отсутствует, поллиний не может осуществлять движений, необходимых для достижения такого положения, которое позволяет ему ударять рыльце следующего цветка. Напротив, в течение дня носик совершает определенные движения, оставляя свободным проход к рыльцу.
Представляется весьма важным введение промежуточного звена, поскольку ножки во многих родах сами постоянно прикрепляются к эпидермису верхней стороны носика. Вместе с подстилающей тканью ножка образует так называемый стебелек, который отделяется от носика, когда удаляющийся опылитель приводит механизм в рабочее состояние. Вязкое (липкое) вещество, находящееся под или на верхушке носика, приклеивает стебелек к насекомому, которое и удаляет поллиний из теки. Весьма изменчивый стебелек несет ту же самую функцию, что и ножки у Orchis; среди орхидей, имеющих стебельки, ножки очень короткие или рудиментарные. Натяжение в стебельке создает основу для взрывного механизма у Catasetinae, в результате действия которого (после его включений поллинарий выбрасывается в воздух и с большой точностью попадает на опылителя. Эти виды частично имеют цветки только с тычинками или только с пестиками.
Для иллюстрации этого явления приведен культурный сорт Tityus (рис. 52). На продольном срезе поллинарий показан черным цветом. Отчетливо виден канал пестика, выходящий из полости рыльца.
Представляется очень интересным наблюдение Додсона о том, что из-за отсутствия сбалансированности запаха, размеров и липкости, цветок некоторых гибридов в природе становится настоящей смертельной ловушкой, убивающей посещающих его пчел.
Nigritella nigra L. (рис. 53)
Психофилия. Щетковидный цветок.
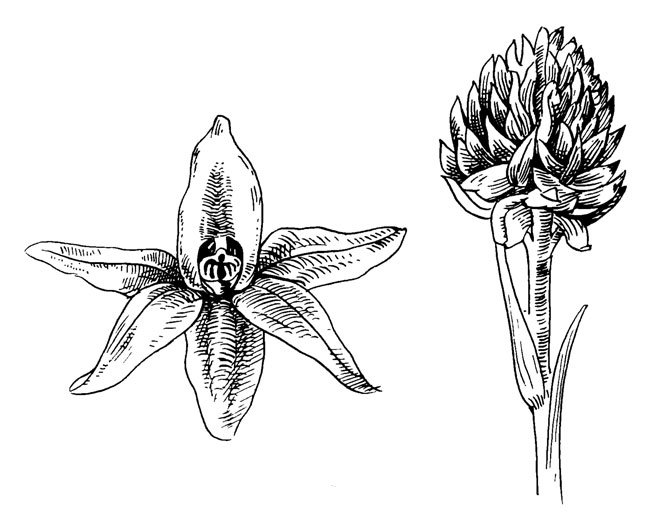
Рис. 53. Nigritella nigra L
Среди орхидных Nigritella представляет собой тип щетковидного соцветия. Дифференциация между членами околоцветника, характерная для орхидных, в основном утрачена. Губа незначительно отличается от других лепестков. Переворачивание цветка - незначительное в щетковидных цветках - также исчезло.
На губе все еще имеется короткий нектароносный шпорец. Хотя длина его всего около 2 мм, нектар может использоваться только дневными бабочками, поскольку отверстие, ведущее в шпорец, слишком узко для более грубого хоботка других насекомых. Было отмечено, что цветки посещаются многими бабочками несмотря на то, что для синдрома психофилии не типичен темный пурпурно-коричневый цвет.
Принцип опыления аналогичен классическому принципу, характерному для орхидных, за исключением того, что положение цветка дает возможность поллиниям самим прикрепляться к нижней стороне хоботка опылителя, и в соответствии с этим меняется движение ножки, приводящей поллиний в положение для удара по рыльцу.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'