17.8. Синдромы опыления у Labiatae-scrophulariaceae
Galeopsis speciosa Mill.
Labiatae. Европа.
Гомогамный. Мелиттофильный. Цветок с зевом.
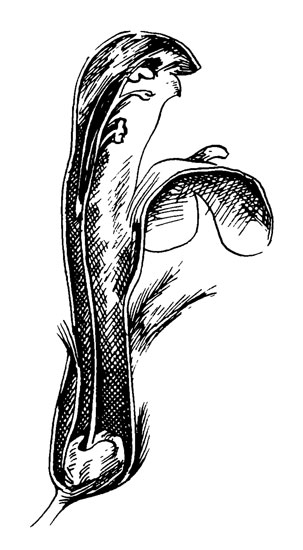
Рис. 33. Galeopsis speciosa
Венчик желтый с бросающимся в глаза темно-фиолетовым указателем нектара на нижней губе, у которой средний составляющий лепесток имеет темно-фиолетовый кончик, тогда как его проксимальная часть темно-желтая с радиально расположенными фиолетовыми пятнами. Общая длина трубки 18-22 мм, причем верхняя часть (6-8 мм) так широка, что насекомое может засунуть в нее свою голову. В нижнюю часть (12-14 мм) может проникнуть только хоботок или язык. Нектар обычно заполняет трубки на несколько миллиметров выше нектарника.
Пыльники и рыльце расположены в верхней части цветка, покрытого шлемовидной верхней губой. Так как пятая тычинка рудиментарна или вообще отсутствует, четыре оставшиеся образуют две пары, одна из которых имеет немного более длинные нити, чем другая. Связник расположен под прямым углом к нити, и тычинки открываются центрипетально (сверху вниз) створками. Они образуют крышечки с бахромчатыми краями, висящие на связнике. Нижняя губа образована тремя лепестками. В основании двух боковых долей имеются выросты, которые облегчают работу пчел в цветке (рис. 33).
Galeopsis посещают крупные шмели. Чтобы достичь той части трубки, в которой расположен нектар, опылителям нужен хоботок длиной по крайней мере 10 мм; кроме того, опылители должны просунуть головы в верхнюю, широкую часть трубки венчика. При таком положении их спинки дотронутся до концов бахромы передней теки и откроют ее. Точно так же при ударе они откроют заднюю теку. Сходным образом они дотронутся до рыльца, которое обычно выступает между двумя передними пыльниками.
Scrophularia nodosa L.
Scrophulariaceae. Европа, Северная Азия.
Протогиния. Энтомофильный цветок, опыляемый осами (?) Колокольчатый.
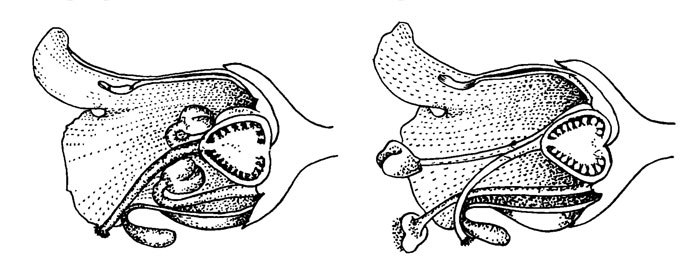
Рис. 34. Scrophularia nodosa L
Зигоморфия цветка S. nodosa (рис. 34) настолько незначительна, что ее можно не учитывать и рассматривать этот цветок как колокольчатый. В первую, или женскую, фазу цветения (слева) пылинки загнуты назад и воспринимающее рыльце находится в зеве цветка. Пятая тычинка образует стаминодий в верхней части цветка; его функция в опылении, если вообще имеется, остается неясной. Установлено, что женская фаза длится 2 дня.
В мужской фазе цветения (справа) выступающая часть столбика слегка загибается и рыльце, которое все еще способно воспринимать пыльцу, становится менее доступно, чем раньше. Тычиночные нити разгибаются и вытягиваются, а пыльники выставляются в зев цветка.
Нектар секретируется кольцевидным нектарником, расположенным в основании гинецея. Крупные насекомые садятся на венчик и просовывают головы внутрь цветка. Мелкие насекомые могут вползать в цветок. Их деятельность в качестве опылителей сомнительна. Со времен Шпренгеля S. nodosa рассматривался как прототип цветка, опыляемого осами. Возможно, что и в самом деле этот цветок чаще всего опыляется осами (по-видимому, они даже предпочитают этот цветок), однако он также часто посещается и пчелами.
При сравнении цветка S. nodosa с цветками Salvia, Pedicularis и т. д. первый может рассматриваться как обратная адаптация к стернотрибическому опылителю. Другим важным вопросом является образование постфлорального нектара, описанное Шреммером (Schremmer, 1959) для S. canina. Вероятно, постфлоральный нектар особенно привлекает ос.
Linaria vulgaris Mill.
Scrophulariaceae. Европа, Западная Азия.
Гомогамный. Мелиттофильный. Вторичное преподношение нектара. Закрытый цветок с зевом, несущий шпорец.
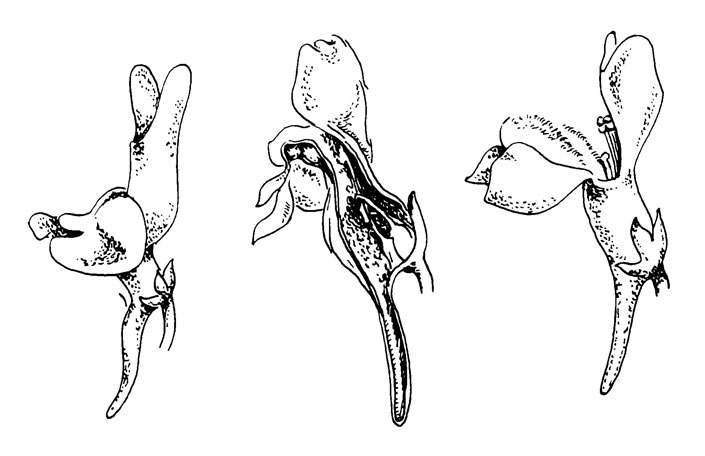
Рис. 35. Linaria vulgaris
Цветок Linaria (рис. 35) представляет собой более совершенный тип цветка с зевом по сравнению с типом такого цветка, представленного у Galeopsis. Для него характерны две главные новые черты. Во-первых, нижняя губа сильно изогнута и закрывает вход в трубку венчика. Это означает, что только насекомые, достаточно сильные для преодоления этого препятствия, могут использовать этот цветок, если они, подобно Macroglossa способны просунуть тонкий хоботок через очень маленькое отверстие между губами. Основание нижней губы формирует своего рода шарнир, поэтому при достаточном усилии вся губа отклоняется вниз.
Во-вторых, нектар находится не в нектарнике, а спускается в трубку венчика и, таким образом, становится недоступным для насекомых с короткими хоботками. В цветке расположены различные ряды волосков, их настоящая функция хорошо не изучена. Предполагается, что некоторые из них, расположенные в шпорце, функционируют как капилляры, переводя нектар из нектарника к основанию шпорца. Однако возможно также, что некоторые из этих волосков оказывают довольно неприятное действие на хоботки шмелей, поэтому едва ли их следует считать указательными структурами.
Melampyrum pratense L.
Scrophulariaceae. Европа, Северная Азия.
Гомогамный. Мелиттофильный. Цветок с зевом (кража нектара).
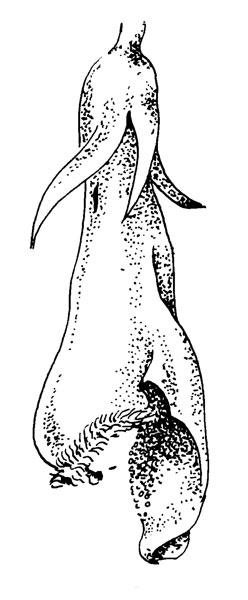
Рис. 36. Melampyrum pratense L. Показано, как воры получают доступ к нектару, прокусывая венчик у его основания. Отмеченное на рисунке отверстие соответствует отверстию, проделанному, например, Bombus lucorum (Meidell, 1945)
Цветок М. pratense относится к довольно простому в функциональном отношении типу цветков с зевом. Он обычно бледно-желтый, длиной 15-20 мм, контрастирующий с темными, коричневато-красными прицветниками. У большинства других видов прицветники и чашечки также ярко окрашены и образуют часть элемента аттрактации.
Ось горизонтальная, цветоножки довольно слабые и под тяжестью опылителя цветки склоняются вниз. Посадочная площадка, предоставляемая нижней губой, мала, но хорошо выраженная шероховатость обеспечивает великолепную точку опоры. Просовывая голову в довольно узкое отверстие цветка, опылитель должен затем его расширить, при этом рыльце наклоняется вниз, а пыльца высыпается на верхнюю сторону насекомого (голову и пронотум). Как обычно у такого типа цветка нектар находится в основании завязи.
На рис. 36 показано, как воры получают доступ к нектару, прокусывая венчик у его основания. Отмеченное на рисунке отверстие соответствует отверстию, проделанному, например, Bombus lucorum (Meidell, 1945). Некоторые воры этого типа могут также собирать пыльцу непосредственно на свои задние ноги, располагая их под пыльниками, в то время как они сами сидят на верхней части верхней губы. Энергично взмахивая крыльями, они вытряхивают пыльцу из пыльников на ноги. Во время полета к следующему цветку насекомое переносит пыльцу в корзиночках. Поскольку нектарные воры не опыляют, опыление должно происходить во время сбора пыльцы. Посещения Megachile willoughbyella, также собирающей пыльцу, отличаются от посещений других насекомых тем, что Megachile в противоположность остальным помещает под пыльником свое брюшко и затем начинает взмахивать крыльями. Megachile собирает пыльцу на нижней поверхности брюшка. Этот опылитель также может получить доступ к нектару, прокусывая венчик, хотя его хоботок достаточно длинен, чтобы собрать нектар обычным путем.
Виды Bombus, имеющие длинные хоботки (В. jonellus, В. pratorum и другие), наоборот влезают в цветок Melampyrum спереди и производят опыление обычным путем. Однако, если другими ворами в венчике были прокусаны дырки, они пользуются такой возможностью; сами они не могут сделать отверстие в венчике. Сходные различия были описаны (Koeman-Kwak, 1973) между коротко- и длиннохоботковыми шмелями, посещающими Pedicularis palustris.
Bartsia alpina L.
Scrophulariaceae. Европа, Северная Америка.
Протогиния. Мелиттофилия. Поздняя автогамия. Цветок с зевом.
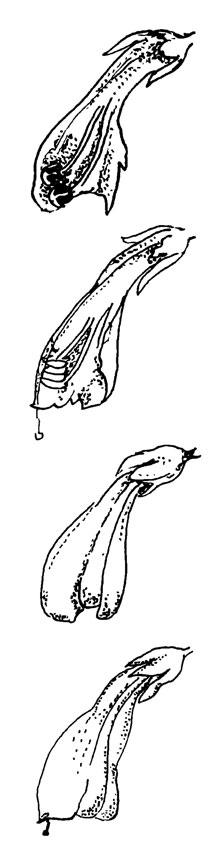
Рис. 37. Bartsia alpina L. (сверху вниз) большой и маленький цветки в женской фазе. Дальше эти же самые цветки показаны в разрезе (позднее цветение у мелких цветков)
Bartsia alpina имеет обычный цветок с зевом, характеризующийся, правда, некоторыми отклонениями. Размер цветка варьирует, причем, возможно, существуют определенные клоны(?) с большими, преимущественно энтомофильными цветками и клоны с мелкими, преимущественно автогамными цветками.
Темный грязно-фиолетовый колос (прицветники и цветки) не очень заметен для человеческого глаза. Возможность отражения ультрафиолетового излучения не исследовалась, но эффект на черно-белых фотографиях свидетельствует о способности отражать ультрафиолет. Однако зарегистрированных посещений (шмелями) мало.
По отношению к общему синдрому мелиттофилии цветок В. alpina аномальный, так как не имеет посадочной площадки. Нижняя губа очень короткая и ее доли повернуты внутрь. Шмели садятся спереди и легко отгибают в стороны доли губы, протискивая головы в цветок. Продольные складки венчика допускают его расширение, особенно в верхней части. Расширение венчика приводит, как обычно, к отделению пыльников и последующему выпадению пыльцы. Интересным является наличие шипов на нитях. Они направляют опылителя и ограничивают его деятельность нижней частью венчика.
У более мелких цветков рыльце едва выступает за пределы венчика, и растяжение последнего в период цветения очень скоро выталкивает пыльники к рыльцу. Наличие спутаннных волосков может в какой-то степени препятствовать автогамии даже в таких типах, но, по-видимому, все же в конце концов происходит самоопыление. Вопрос о том, являются ли маленькие цветки автогамными всегда, а большие - лишь в редких случаях, предстоит еще решить.
Pedicularis oederi Vahl.
Scrophulariaceae. Европа, Азия.
Протогиния (или гомогамия?). Мелиттофилия. Цветок с зевом.
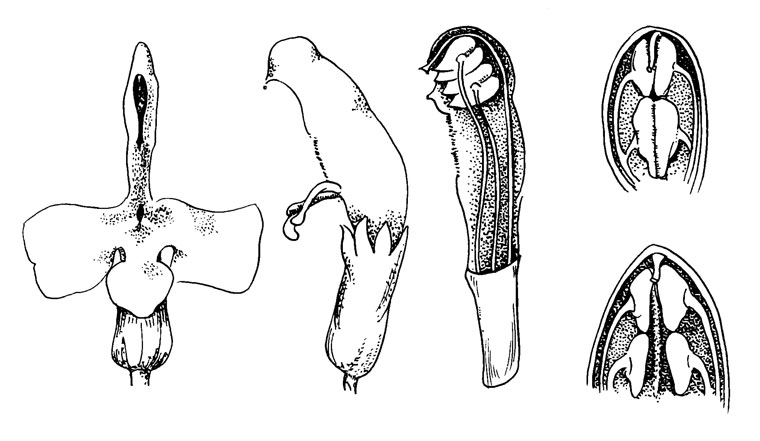
Рис. 38. Pedicularis oederi
P. oederi представляет собой примитивный тип внутри этого рода, у которого можно наблюдать ряд поразительных адаптации к опылению (Li, 1948-1949). P. oederi были изучены Шпрагу и недавно более тщательно Масиором (Macior; см. список литературы).
Цветок желтый, верхняя часть верхней губы внутри темно- красная (пятнистая), прямостоячий, в узком колосе; длина цветка 15-20 мм. Верхняя губа слегка наклонена вперед, нижняя - на 45° вниз. Верхняя губа сжата сильной жилкой, проходящей на каждой стороне от середины трубки венчика наклонно к середине свободных краев верхней губы. Края ниже этой жилки подвижны и покрыты жесткими волосками. Нижняя губа разделена на доли двумя мощными рубцами, разделяющими доли по направлению к открытой трубке венчика. Боковые доли жесткие, но гибко связанные со сторонами верхней губы.
Цветок посещают крупные шмели, которые садятся спереди и укрепляются на долях нижней губы. Маленькие ранки показывают, где были точки опоры для ног опылителей. Избегая колючих краев, опылитель проталкивает хоботок и голову в трубку под ними, т. е. через проход, образуемый этими двумя краями на нижней губе. Это вызывает расширение верхней части трубки венчика и, вследствие этого, наклон верхней губы по направлению к спинке опылителя. Благодаря своему наружному положению (и протогинии?) рыльце первым касается спинки опылителя, и пыльца сама осыпается практически одновременно, благодаря небольшому расширению верхней губы, причем тека разрывается. После того как насекомое улетает, трубка венчика вновь сужается и верхняя губа возвращается в свое первоначальное положение, при повторном посещении все повторяется снова.
Поскольку этот механизм может срабатывать только при опылении крупными шмелями, небольшие шмели собирают пыльцу, силой раздвигая концы верхней губы и заставляя сухую пыльцу осыпаться на их брюшко, - они работают в перевернутом положении. Это поведение до некоторой степени напоминает работу, необходимую для получения нектара у Р раlustris и у видов с рострумом.
На рис. 38 (слева направо) изображено: цветок спереди и сбоку, продольное сечение верхней губы. На крайних правых рисунках показано: вверху - обычное закрытое положение вместилищ пыльцы и внизу - положение, когда пыльники силой раздвинуты в стороны.
Pedicularis silvatica (рис. 39)
Scrophulariaceae. Европа.
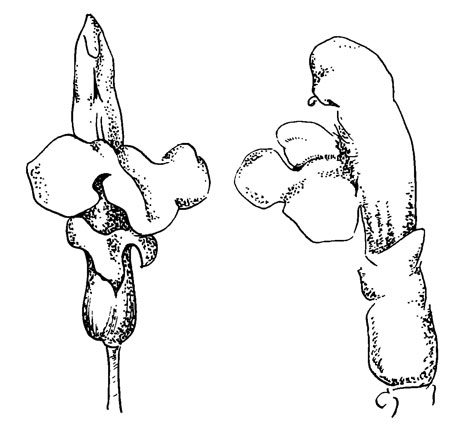
Рис. 39. Pedicularis silvatica
Цветок на рис. 39 проявляет очень слабую тенденцию к наклону, так хорошо заметную у более развитых видов. Цветки розовые, посещаются шмелями. Механизм опыления (насколько это известно) такой же, как и у Р oederi.
Pedicularis palustris (рис. 40).
Scrophulariaceae. Европа.
Протогиния (или гомогиния?). Мелиттофилия. Цветок с зевом.
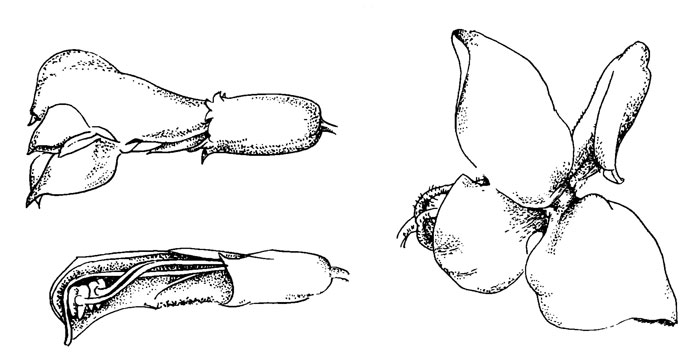
Рис. 40. Pedicularis palustris
Цветки розовые длиной 10-20 мм. Отличия от Р. oederi состоят в следующем: цветок более или менее горизонтальный, угол между верхней и нижней губой острый. Верхняя губа слегка наклонена и закручена, и края около верхушки с двумя нитевидными зубчиками поддерживают выступающий столбик. Колючие края с обеих сторон заканчиваются заметными крепкими зубчиками. Плоскость нижней губы повернута приблизительно на 45°, показывая морфологически правую сторону. Доли нижней губы менее разделены, края менее выступают.
Благодаря закручиванию нижней губы, проникнуть в цветок легче с левой стороны. Об этом свидетельствует также и способ соединения губ; с правой стороны края губ свернуты друг с другом и их нельзя разделить без применения силы. Левые края свободны, но укреплены и имеют шипы. Нектарник также наклонный.
Опылители садятся сбоку и влезают в цветок с левой стороны. Если они подходят с правой стороны (например, переползли с другого цветка), то сначала они занимают правильное положение. С помощью мандибул опылитель с силой открывает верхнюю губу над колючими краями, как раз над выступающими (крепкими) зубчиками. Затем он просовывает туда голову, оставаясь над колючими частями краев. Как и у Р. oederi при открывании цветка трубка венчика расширяется, что приводит к таким же последствиям, но в этом случае закручивание и асимметрия цветка временно нарушается из-за расширения. После посещения цветок восстанавливает свою форму.
Pedicularis lapponica (рис. 41)
Scrophulariaceae. Полярные области.
Р. lapponica сходна с P. palustris за исключением того, что здесь нижняя губа (схожая с Р oederi) закручена на 90°.
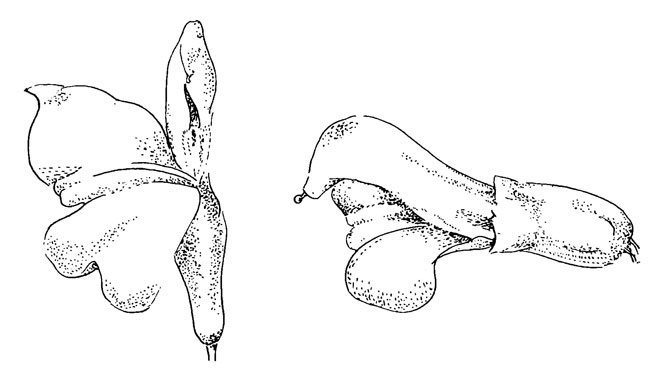
Рис. 41. Pedicularis lapponica
Работа опылителей на этих цветках грубая и оставляет отметки на лепестках, особенно заметные у P. lapponica; незначительные отметки коготков остаются на нижней губе, на которую опылитель ставит ноги во время расширения трубки венчика, и значительные - на верхней губе, которую он с силой выворачивает. После нескольких посещений, цветки, особенно хрупкие цветки P. lapponica, выглядят сильно поврежденными и плохо функционируют; но к этому времени они бывают уже опылены.
Pedicularis racemosa Dougl. (рис. 42)
Scrophulariaceae. Северо-запад США.
Протогиния(?) Мелиттофилия. Модифицированный цветок с зевом.
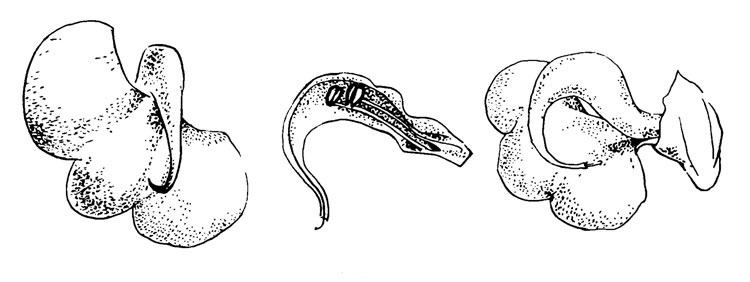
Рис. 42. Pedicularis racemosa
По своему обычному положению цветок напоминает Р. раlustris и P. lapponica, но отличается длинным рострумом на шлеме. Рострум служит каналом, который поддерживает только столбик: пыльники расположены вблизи рострума; таким образом, в этом цветке не наблюдается близкого расположения рыльца и пыльников, что характерно для других видов.
Опыление у этого вида было описано Масиором (Macior, 1970). Цветок не имеет нектара и посещается только сборщиками пыльцы. Шмели приближаются к венчику с вытянутыми антеннами, захватывают венчик передними и средними ногами, а шлем сбоку - мандибулами. Оставаясь против широкой нижней губы венчика, они висят в перевернутом положении и собирают пыльцу, вибрируя крыльями. Спиральный рострум цветка с заключенным в него столбиком и торчащим рыльцем контактируют с нижней стороной головы и передней частью торакса, тогда как пыльца, вытряхиваемая из пыльников, расположенных в шлеме, падает на перевернутое вверх брюшко насекомого. Иногда сборщики счищают пыльцу в корбикулы, продолжая висеть на венчике при помощи своих мандибул. (Из Macior, с сокращениями).
Pedicularis groenlandica Retz.
Scrophulariaceae. Северная Америка.
Гомогамные(?). Мелиттофильные. Пыльцевая аттрактация.
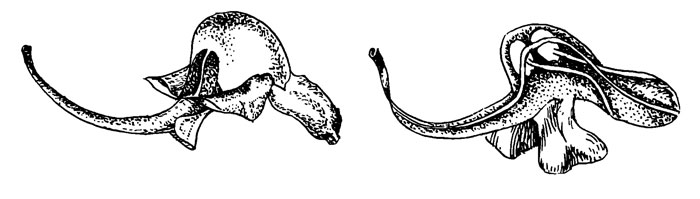
Рис. 43. Pedicularis groenlandica
У этого вида (рис. 43) шлем не только клювовидный, как у P. racemosa, но вытянут наподобие хоботка, который при полном цветении делает один завиток, образованный главным образом дистальной частью рострума (Macior, 1968 а, 1977). Как у P. racemosa, в роструме находится только столбик, причем пыльники остаются в выпуклой базальной части шлема. Опыляющие насекомые (различные виды Bombus) садятся на верхушку рострума и цепляются передними ногами за боковые нижние лепестки. Мандибулами насекомые захватывают средний край передней поверхности выпуклой части шлема. Рострум частично поддерживает опылителя (последний опирается на рострум средними ногами) и обвивает его тело между грудью и брюшком, ориентируя рыльце "точно на центр передней абдоминальной поверхности".
Нектара нет, пыльца собирается при помощи вибрации крыльев, которая вызывает появление около шлема небольшого желтого облака, оседающего на теле насекомого. Частота взмахов крыльев на цветке отличается от частоты взмахов при полете. Пыльца, прилипшая к телу насекомого, затем счесывается и переносится в корбикулы. Остаток пыльцы на передней абдоминальной поверхности попадает на рыльце следующего цветка.
Очевидно, работа этого очень точного механизма зависит от правильного соотношения размеров опылителя и цветка. Длина головы и груди опыляющих рабочих шмелей, 7-8 мм, соответствует по размеру цветку, имеющему 8 мм между передней частью выпуклой части шлема и петлей рострума. Слишком крупные матки не приспособлены к опылению этого цветка. По сравнению с видами, упомянутыми выше, P. groenlandica характеризуется 1) безнектарными цветками с пыльцой. В развитии синдрома опыления у этого рода для большего совершенства и точности происходит возврат к аттрактантам, имеющим более раннее происхождение; 2) происходит также возврат к стернотрибическому опылению (если оно более примитивно, чем нототрибическое), но в противоположность стернотрибическому опылению у Р. racemosa рыльце в этом случае сначала касается спинки опылителя, а затем уже брюшка - своего рода супернототрибическое опыление.
Pedicularis sceptrum-carolinum L.
Scrophulariaceae. Северная Европа.
Протогиния. Мелиттофильный. Закрытый цветок.
Цветок прямостоячий, желтый, с красными долями нижней губы, около 30 мм длиной.
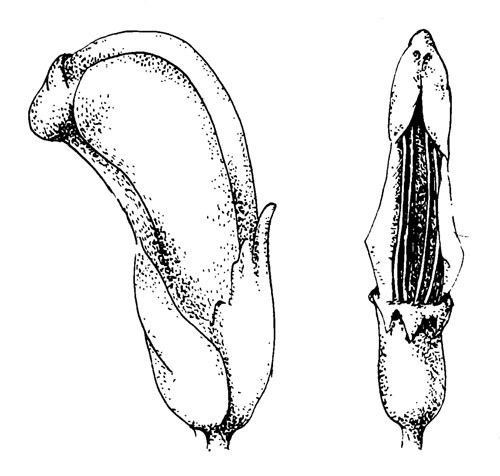
Рис. 44. Pedicularis sceptrum-carolinum L. Справа показан шлем, снизу: видно, что загнутые внутрь края лепестков образуют нечто вроде настила, на котором находится свободно лежащая пыльца.
Для этих цветков характерны те же самые общие черты, что и для P. oederi и P. palustris, однако у P. sceptrum-carolinum цветки постоянно закрыты. Цветки Р. capitata представляют собой промежуточную форму (Macior, 1975), которые еще открыты для рабочих шмелей. Верхняя губа шире, чем у этих двух видов, и пыльца высыпается из пыльников перед посещением. Так как края верхней губы загнуты внутрь, пыльца не высыпается на нижнюю губу или в трубку венчика. Колючих краев нет. Свободные доли нижней губы очень короткие и загнуты внутрь, захватывая конец верхней губы и включая также рыльце, которое слегка выдается из верхней губы. Изгиб нижней губы, особенно характерный для Р. palustris имеется также у Р. sceptrum- carolinum; его можно увидеть, если попытаться открыть цветок; с правой стороны края двух губ скручены вместе так, что фактически их нельзя разделить, не повредив при этом цветок. Левая сторона может быть открыта и такой открытый цветок напоминает цветок P. palustris.
Только очень сильные и "интеллектуальные" шмели могут обращаться с этим цветком. Они садятся на верхушку прямостоячего цветка, перемещаются на левую сторону и с силой оттягивают нижнюю губу, определенным образом упираясь в цветок ногами. Открыв цветок, насекомое сбоку вползает в широкую трубку венчика, и скрывается в нем. У этого вида при расширении цветка верхняя губа также опускается, а ее загнутые внутрь концы разделяются, что приводит к опылению и отложению пыльцы.
Иногда менее "интеллектуальные" опылители пытаются влезть в цветок с "неправильной" стороны. Забавно наблюдать, как цветок "отталкивает" насекомое, если последнему удается хотя бы немного просунуть голову с правой стороны. Несмотря на имеющиеся трудности, цветки интенсивно посещаются шмелями, достаточно сильными и достаточно "интеллектуальными", чтобы уметь с ними обращаться. Без сомнения, это происходит благодаря большому количеству нектара, доступному для опылителей.
Pedicularis lanceolata Michx.
Scrophulariaceae. Северная Америка.
Протандрический. Пыльцевая аттрактация. Мелиттофильньш. Закрытый цветок.
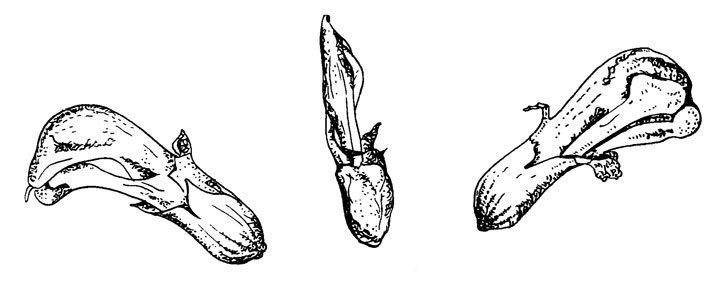
Рис. 45. Pedicularis lanceolata
Этот цветок (рис. 45) образует пыльцу в качестве аттрактанта, опыление его также было описано Масиором (Macior, 1969). Цветок похож на цветок Р. groenlandica, но без рострума, характерного для другой секции (Li, 1948, 1949). Как и родственный ему Р. sceptrum-carolinum, он закрыт, но в отличие от содержащих нектар полностью закрытых цветков этого типа, цветок Р. lanceolata открыт у основания. В этом отношении наблюдается очень интересная параллель с цветком Coronilla emerus, который отличается от родственных нектароносных видов открытым цветком. Однако сростнолепестный, венчик у Pedicularis не может открыться так же легко, как раздельнолепестный венчик Papilionaceae, и эффект у Р. lanceolata главным образом достигается укорочением трубки венчика.
Опылители (рабочие шмели) садятся на верхушку шлема, поворачиваются к нижней стороне венчика и выравниваются с главной осью цветка, головой к основанию. В этом положении они захватывают свободный край шлема мандибулами, а верхнюю часть - средними ногами. Пыльца извлекается передними ногами. При сборе пыльцы крылья не вибрируют.
Опыление стернотрибическое, т. е. рыльце касается оставшейся массы пыльцы в более или менее недоступных частях брюшка насекомого.
Salvia patens Cav.
Labiatae. Мексика.
Протандрический-гомогамный. Мелиттофильный. Цветок с зевом.
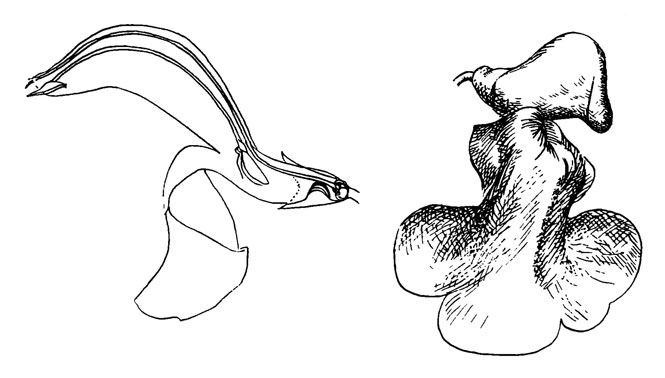
Рис. 46. Salvia patens
Цветки темноголубые, образуют открытый колос, длина венчика 25-30 мм (рис. 46). Очень похожи на цветки Galeopsis speciosa, но отличаются от него более горизонтальным положением цветка, более широкой и короткой (колоколовидной) трубкой венчика, числом тычинок, редуцированным до двух. Однако главное различие заключается в морфологии тычинок. Тычиночные нити срослись с венчиком за исключением короткого, жесткого конца, который выступает более или менее перпендикулярно к общему направлению цветка. Пыльник соединен с нитью связником таким образом, что может закручиваться приблизительно на 180°. Связник 15-18 мм длиной с очень неравными плечами. Более длинное направлено вверх и несет фертильную теку, положение которой соответствует положению пыльников у Galeopsis. Другое плечо очень короткое и несет на нижнем конце стерильную, похожую на ложку теку. Его положение перпендикулярно положению цветка. Две сросшиеся стерильные теки между ними более или менее плотно закрывают вход в трубку венчика.
Нектарник находится в основании завязи. Непосредственно над ним полость венчика сужается в результате образования выпуклости в нижней части трубки.
Шмели садятся на очень большую нижнюю губу и продвигаются к нектарнику, расположенному на дне трубки венчика. Для того чтобы достичь его, они должны раздвинуть створки, образованные двумя сросшимися стерильными теками, которые преграждают путь. Это приводит в действие рычаги, заставляющие фертильные теки опускаться и ударять по спине опылителя, оставляя на ней пыльцу. Рыльце независимо от этого механизма; оно выступает из цветка и во всяком случае в последнюю фазу цветения опускается настолько низко, что задевает спину опылителя даже перед тем, как опустится пыльник.
Этот общий тип опыления обнаружен у многих видов Salvia, причем главным образом у видов с голубыми цветками. Вообще имеются как редуцированные, так и нормальные цветки, более мелкие и(или) однополые (женские с более или менее редуцированным андроцеем).
Salvia splendens Sellow
Labiatae. Бразилия.
Протандрический - гомогамный. Орнитофильный. Трубчатый цветок.
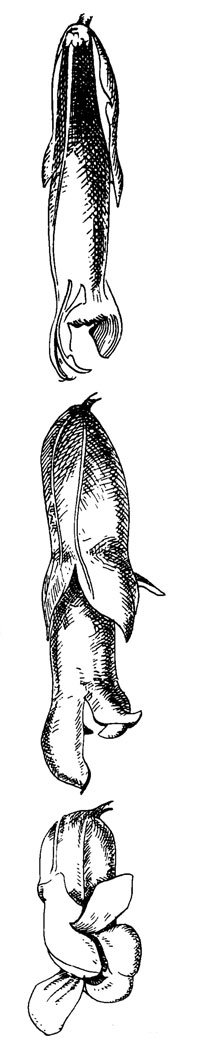
Рис. 47. Salvia splendens
Этот цветок (рис. 47) с огненно-красным венчиком и чашечкой без какой-либо посадочной площадки обнаруживает типичные черты орнитофилии или синдрома опыления колибри. Для полной оценки этого синдрома цветок Salvia splendens следует сравнить с мелиттофильным цветком S. patens.
Механизм рычагов не функционирует, и доступ внутрь цветка не загорожен. Оседание пыльцы неизбежно, когда птица всовывает клюв в цветок (Trelease, 1881).
Виды Mentha
Labiatae. Европа.
Протандрический. Миофильный. Щетковидный цветок.

Рис. 48. Mentha
Вместе с близкими родами Mentha (рис. 48) представляет тип щетковидного цветка в семействе Labiatae. Более простые цветки этих родов, по-видимому развились из типа, представленного цветком Galeopsis, в результате редукции отдельных частей. Цветки еще сохраняют следы зигоморфии, а пятая тычинка, функционирующая в цветке с зевом, также редуцирована в этом псевдоактиноморфном цветке. Сильно выступающие тычинки и рыльца характерны для синдрома щетковидных цветков.
Кроме гермафродитных цветков, имеются также исключительно женские цветки, иногда расположенные на раздельных стеблях. Они мельче, чем цветки гермафродитного типа (длина трубки венчика 2-3 мм по сравнению с 3-5 мм), и полагают, что опылители посещают их после гермафродитных цветков.
Опылителями являются главным образом жесткокрылые и особенно двукрылые, даже если обильное выделение нектара в основании завязи может случайно привлечь опылителей более высокого класса. Thymus с похожими цветками очень активно опыляют пчелы.
Coleus frederici Taylor
Labiatae. Ангола.
Мотыльковый (перевернутый с зевом) цветок.
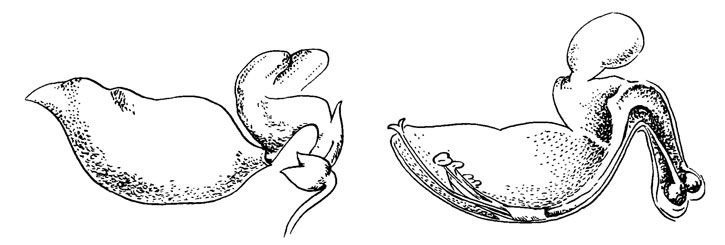
Рис. 49. Coleus frederici
О естественных опылителях видов Coleus известно очень мало. Чрезвычайно сложный цветок с несомненностью предполагает посещение этого цветка эутропными насекомыми и в некоторых случаях птицами. Однако показанный на рис. 49 цветок вряд ли орнитофилен. Ни одна из птиц не могла бы просунуть клюв в изогнутую трубку этого цветка. Голубой цвет также предполагает другие классы опылителей.
С. frederici, изображенный на рисунке, представлен как пример перевернутого цветка с зевом, а функционально - как мотылькового цветка. И если реверсия у Centrosema обусловлена действительным переворачиванием, то у Coleus это результат перенесения тычинок и столбика из верхней в нижнюю часть цветка.
В этом случае общая модель цветка Labiatae - Scrophulariaceae сильно изменена. Две доли, обычно образующие шлем, малы и разделены. Они плоско выступают над остальным цветком и создают видимый фон. Боковые доли нижней губы почти неразвиты, тогда как центральная доля образует большой сдавленный с боков карман, из которого выступают репродуктивные органы. Эта доля подвешена к остальной части цветка. Трубка венчика сравнительно жесткая и сильно изогнута.
Тычинки срослись с венчиком до того места, где подвешена нижняя губа. Отсюда и далее приблизительно наполовину сросшиеся тычинки образуют жесткий канал, окружающий столбик. Эта структура, не похожая на что-либо в обычных цветках с зевом, придает необходимую жесткость репродуктивным органам. Следовательно, пыльники и столбик остаются на месте, когда средняя доля отгибается вниз под тяжестью тела опылителя.
Изгиб происходит там, где средняя доля подвешена к основанию цветка. На рис. 49 (справа) показано, что венчик и трубка тычиночных нитей разделены почти до конца.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'