17.5. Цветки, в которых выводится потомство
Calluna vulgaris L.
Ericaceae. Европа.
Цветок протандрический, энтомофильный, гемитропный. Факультативная анемофилия. Элементы аттрактации от чашевидных до колокольчатых.
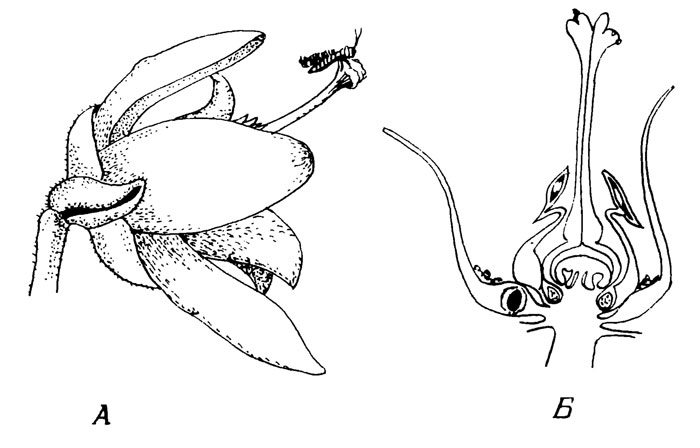
Рис. 15. Calluna vulgaris L
Цветки тетрамерные, почти радиальные, чашевидные, около 4 мм в поперечнике, с розовыми чашелистиками, более заметными, чем мелкие лепестки такого же цвета. Нижний лепесток открывается больше, чем другие. Это делает цветок слегка зигоморфным. Нижние части тычиночных нитей сочные, но их основания очень тонки. Нектарник образует непрерывное хорошо заметное кольцо вокруг завязи. Нектар доступен для насекомых, способных протискивать свои хоботки между нитями, надавливая на пыльники. Теки открываются латерально; пыльца высыпается, когда пыльники отделяются друг от друга. Роговидные отростки пыльников функционируют по типу рычага. Нектар образуется в больших количествах, поэтому Calluna посещается множеством двукрылых, перепончатокрылых и чешуекрылых. Точность и эффективность этих посещений для опыления не очень велика. Однако Calluna - важный медонос, так как содержит большое количество нектара и распространена очень широко.
В последнюю фазу цветения количество нектара уменьшается, нити подсыхают и пыльца рассеивается ветром. Следовательно, возможно ветроопыление. Пыльцы образуется очень много. По данным Пола (Pohl, 1937b), из всех исследованных растений у Calluna наблюдается наибольшая продукция пыльцы на единицу площади, а именно более чем 4*109 тетрад на 1 м2. Число пыльцевых зерен на завязях относительно низко, порядка тысячи (Hagerup, 1950а). При анализе обнаруживается, что в воздухе действительно содержится большое количество пыльцы Calluna.
Наиболее интересен тип опыления, осуществляемый Taeniothrips ericae и описанный Хагерупом на Фарерских островах, но встречающийся также и в других местах. Небольшое число Т. ericae постоянно находятся в цветках Calluna, достаточно крупных, чтобы полностью скрыть насекомых. Эти животные постоянно движутся и способны проникать между пыльниками и пестиком к нектарнику, что недоступно для более крупных опылителей. Во время движений трипсов некоторые пыльцевые зерна освобождаются из пыльников и приклеиваются к их телам, липким от нектара.
Самцы Т. ericae более редки, чем самки, и не способны летать, поэтому самки перелетают с одного цветка на другой в поисках полового партнера. Они останавливаются перед выступающими рыльцами и садятся на них, делая, таким образом, возможным само- или перекрестное опыление.
После оплодотворения самки Т. ericae влезают в цветки, поедают больше пищи в виде нектара, а также вырезают суккулентные части тычиночных нитей и лепестков; в последние они откладывают четыре яйца. Зимой яйца спрятаны в устойчивых венчиках, а личинки появляются в следующем году. Они обитают в цветках вместе со взрослой стадией насекомых. Полагают, что окукливание происходит на земле, а взрослое насекомое появляется только в следующем сезоне.
Таким образом, Т. ericae способен проводить всю свою активную жизнь в цветке Calluna. Очевидно, это насекомое может также жить и в других растениях (Hagerup, Hagerup, 1953).
На рис. 15, А изображен цветок Calluna с Т. ericae в соответствующем масштабе. На рис. 15, Б показан продольный разрез через середину цветка Calluna с нектарником (отмечен точками) и яйцом Т. ericae (черное) в основании лепестка.
Trollius europaeus L.
Ranunculaceae. Европа.
Последовательное центрипетальное созревание тычинок и пестиков.
Растение энтомофильное, цветок эутропный (?). Элемент аттрактации закрытого типа.

Рис. 16. Trollius europaeus L
Цветки желтые (рис. 16), почти шаровидные, диаметром 3 см, с большим числом перекрывающихся, ярко окрашенных чашелистиков, которые полностью закрывают внутреннюю часть. Один ряд лепестков образует оберточные нектарные листья. Число обнаруживаемых крупных опылителей невелико. Самоопыление неизбежно, но, очевидно, цветки самонесовместимы. Наиболее обычным обнаруженным до сих пор опылителем является маленькая муха Chiastochaeta trollii, которая может проникать между чашелистиками. Цветок образует полость, в которой муха живет очень долго, питаясь пыльцой и нектаром, вызывая беспорядочное и грязное опыление, так как она до этого посещала другие растения. Чаще всего самцы обнаруживаются вне цветков, а самки - в цветках Trollius.
Яйца откладываются в основании пестика. Вылупившиеся личинки пробуравливают пестик и питаются развивающимися семязачатками. Личинки заканчивают свое развитие приблизительно через 10 дней и снова забираются в проделанные ими отверстия в еще молодых и мягких плодах, а затем покидают цветки (окукливание происходит на земле?). Личинки не съедают все завязи, даже в захваченной ими части плода. Некоторые пестики свободны от насекомых.
На рис. 16 слева показан поперечный срез цветка с опылителем (Hagerup, Peterson, 1956). Возможно также, что цветки Trollius опыляются шмелями, особенно открытые цветки других видов. В середине - продольный разрез через молодой плод, в котором видна пустая оболочка яйца (внутри) и повреждения, вызванные личинкой. Справа - почти созревший плод с входным (маленьким) и выходным (большим) отверстиями, проделанными личинками.
Виды Yucca
Liliaceae. Северная Америка.
Херкогамия. Гомогамия. Эуфилия. Фаленофилия. Опыление при выведении потомства.
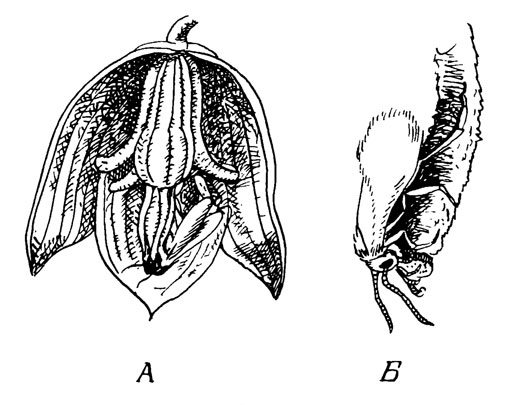
Рис. 17. Yucca
Цветки (рис. 17) висячие, колокольчатые, с тонкими желтовато-беловатыми членами околоцветника (большая часть рисунков показывает опылителей, но головой вниз). Большинство видов полностью неспособны к автогамии из-за взаимного расположения пыльников и рыльца. Нектарники расположены внутри завязи (около перегородки), они вряд ли играют заметную роль в синдроме опыления. Опылитель - ночная бабочка Tegiticula (Pronuba) yuccasella. Тычиночные нити толстые и негнущиеся, пыльники довольно маленькие, пыльца почти порошкообразна. Очевидно, ночные бабочки совсем не питаются; самки влезают в цветок и собирают пыльцу из пыльников (рис. 17, А). Наличие зубовидного отростка позволяет переносить относительно большие количества пыльцы. Пыльца транспортируется к гинецею и откладывается здесь в полости рыльца (рис. 17, Б). В это время в завязь откладываются яйца, и личинки поедают развивающиеся семязачатки. На созревших плодах видны отверстия, проделанные личинками Tegiticula, и недоразвитые части коробочки, в которой съедены все семена (обсуждение этого вопроса см. в работе Powell, Mackie, 1966).
Виды Ficus
Moraceae. Средиземноморье.
Однодомные. Вторичная протогиния. Энтомофилия. Эутропные, похожие на урну соцветия.
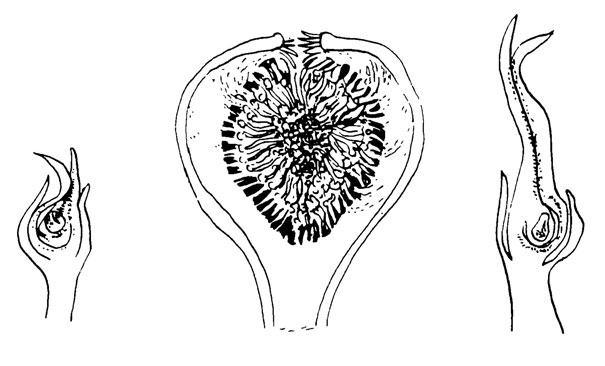
Рис. 18. Ficus
Вследствие большого экономического значения съедобный инжир, F. carica, был первым видом, который изучался с точки зрения опыления. В течение тысячелетий опыление инжира представляло собой серьезную проблему для земледельцев. Особенно тщательные исследования проводились в последней половине XIX в. Тем или иным путем эти проблемы, как научные, так и практические, решены. Однако в широком смысле слова они были решены не совсем удачно. Оказалось, что F. carica является аберрантным типом, во многих отношениях очень сложным. Только после того, как в 1969 г. Галил (Galil) с соавторами, а также Рамирец (Ramirez) опубликовали свои работы, проведенные на другом виде, оказалось возможным лучше понять то, что происходит у F. carica.
Соцветия рода Ficus напоминают по внешнему виду урну. Это так называемый циконий, внутренняя поверхность которого представляет собой ввернутую внутрь поверхность оси соцветия колос, несущую цветки (рис. 18). Цветки сильно редуцированы: тычиночный цветок состоит главным образом из одной или от одной до пяти тычинок, тогда как пестичный цветок имеет единственный пестик и три рудиментарных члена околоцветника. Эти цветки никогда не выступают из цикония, выход из которого представлен единственным узким отверстием - осциолюмом, обрамленным большим числом чешуек, представляющих собой стерильные брактеи на морфологически нижней части колоса.
Для цикония характерна четко выраженная протогиния. Галил различает пять фаз в ходе развития цикония: А. Незрелые почки. Б. Цветение пестичных* цветков. В. Пауза. Г Цветение тычиночных цветков. Д. Созревание и распространение семян.
* (Для того, чтобы избежать путаницы, мы будем называть цветки "пестичные" и "тычиночные", а термины "мужские" и "женские" употреблять только в отношении опылителей.)
Общий принцип опыления состоит в следующем: самки небольших ос, длиной всего лишь несколько миллиметров (представители сем. Agaonidae), нагруженные пыльцой, проникают в циконий на стадии Б и откладывают яйца в пестики пестичных цветков. Семязачатки образуют галлы. Во время паузы развиваются личинки, а в начале фазы Г появляются самцы и оплодотворяют самок внутри галлов. Затем нагруженные пыльцой самки перелетают в другой циконий, находящийся на стадии Б, откладывают яйца и т. д.
При изучении инжира было обнаружено, что пестичные цветки бывают двух видов: одни с короткими, другие с длинными столбиками. Яйцеклад различных ос достигает семязачатков лишь в цветках с коротким столбиком. Яйца откладываются точно в семязачаток, и никаких семян не образуется. Длинные столбики слишком длинны, и яйцеклад не достигает семязачатка. После неудачных попыток оса удаляет яйцеклад из столбика и семязачатка, который в это время был опылен, и семя развивается.
Значительное различие между двумя классами пестичных цветков, - возможно, более позднее явление; примитивные виды инжира содержат пестичные цветки с непостоянной длиной столбика. Таким образом, мы можем предположить, что этот синдром развивается в ровном, тропическом климате, где циконии созревают непрерывно, в результате чего появившиеся женские особи всегда находят циконии на стадии Б. На рис. 19, А приведена схема опыления и образования семян в трех поколениях инжира этого типа, так называемого однодомного инжира.
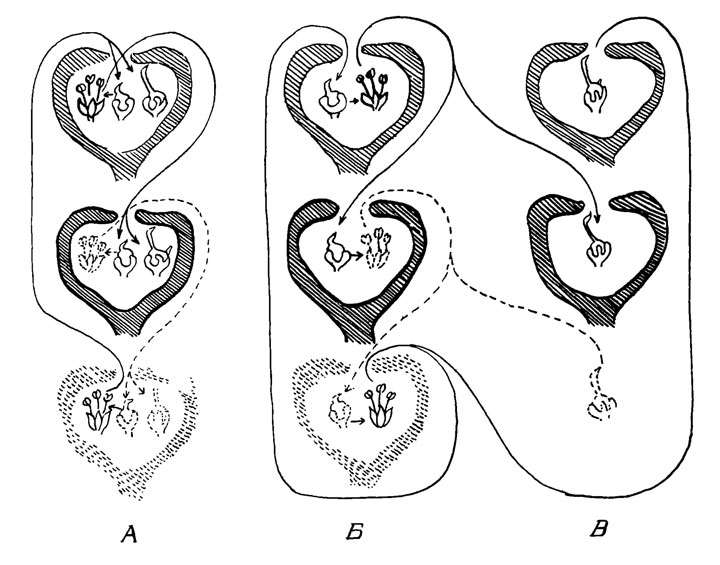
Рис. 19. Схема опыления и образования семян в трех поколениях инжира этого типа, так называемого однодомного инжира
Однако имеются и другие виды инжира (рис. 19, Б и В), у которого два типа цикониев: одни с тычиночными цветками и с короткими столбиками, другие с пестичными цветками и с длинными столбиками. Эти растения (морфологически женские растения, имеющие женские и двуполые цветки на разных растениях) - функционально двудомные, так как в цикониях одного типа образуется только пыльца и никогда не образуется семян, а в цикониях другого - образуются только семена, а не пыльца. Размножающаяся популяция ос поддерживается галлами, развивающимися в тычиночных цикониях.
За исключением крайней протогинии, этот синдром не очень далек от опыления Trollius и Yucca. Главное значение открытий Галила и Рамиреца состояло в том, что у изученных ими видов Ficus пыльца переносится активно, так же как и у Yucca. Самки опыляющих ос на обеих сторонах торакса имеют карманы; в них они активно собирают пыльцу перед тем, как покинуть циконии, в которых они развивались.
Число и положение тычиночных цветков по отношению к пестичным весьма различны в зависимости от вида. У некоторых видов тычиночные цветки распределены случайно, тогда как у других они сконцентрированы в отдельной зоне около осциолюма. У некоторых видов последнего типа самцы разрезают тычиночные нити перед тем, как покинуть циконий. Отделенные пыльники падают и располагаются среди пестичных цветков, из которых должны появиться самки. В некоторых случаях самки активно открывают пыльники и выгребают пыльцу. Галил и др. (Galil et al., 1973) наблюдали также процесс опыления у F. costaricana и F. hemsleyana. Пыльца захватывается ложкообразными крайними суставами передних ног - ареолами - и откладывается главным образом на брюшке осы. Отсюда она вычесывается щеточками на передних коксах и откладывается в вентральных карманах на обеих сторонах торакса. Искривления торакса расширяют отверстия в карманах и облегчают помещение в них пыльцы. Опорожнение карманов повторяет процесс в обратной последовательности, причем пыльца снова переносится к ариолам (Galil, Eisikowitch, 1973). Опыление достигается за счет активного переноса к рыльцу небольшого числа пыльцевых зерен после каждой яйцекладки.
У некоторых цикониев рыльца образуют непрерывную поверхность - синстигму - и самки прокусывают окружающие рыльца после откладывания яиц в один семязачаток. В результате многие бесполезные обитатели цикония и паразиты не могут откладывать яйца около галлов или в галлы, в которых обитают опылители.
У некоторых видов во время фазы В внутри цикония значительно возрастает содержание двуокиси углерода. Когда появляются самцы, содержание СО2 может достичь 10%. При такой концентрации самцы сохраняют свою активность, а самки - нет. После оплодотворения самок, находящихся еще в галлах, и срезания пыльников самцы проделывают отверстия в стенке цикония (но не пользуются осциолюмом!) и покидают циконий. Концентрация двуокиси углерода постепенно снижается, и самки становятся активными, начинают собирать пыльцу и нагружать карманы, расширяют, если необходимо, проделанное самцами отверстие и тоже покидают циконий, чтобы найти другой, находящийся в фазе Б, где они могут отложить яйца. В тонкостенных цикониях других видов во время процесса созревания атмосфера сохраняется "нормальной".
Этот процесс опыления был назван Галилом "этодинамическим". Как и у Yucca, он основан на активной работе опылителя. Опылитель F. carica, Blastophaga psenes не имеет торакальных карманов, и большая часть пыльцы транспортируется пассивно и беспорядочно между тергитами осы; пыльца пассивно отлагается на рыльце пестичного растения, куда насекомые откладывают яйца. Такое опыление Галил называет "топоцентрическим". Из-за менее точного механизма переноса в топоцентрическом циконии значительно больше пестичных цветков, чем в этодинамическом. Излишек пыльцы с поверхности тела самок счесывается перед тем, как они покинут циконий. F. carica - растение с очень длительной историей одомашнивания, оно имеет очень много клонов, причем некоторые из них партенокарпические. Кроме того, для культурного инжира характерно наличие особей противоположного пола - каприфиг. И инжир и каприфига - высоко специализированные культурные растения, которые могут размножаться вегетативно. Размножение семенами дает особи "дикого" типа так называемый эриноцис. Культивируемый съедобный инжир и каприфига функционируют вместе как двудомный вид, тогда как эриноцис в основном соответствует однодомному типу. Процесс "капрификации", т. е. набрасывание веток каприфиги в крону фиговых деревьев, - метод для получения гарантированной продукции плодов - хорошо известен уже в течение нескольких тысяч лет. Понятно, что эта техника затруднительна для обычного практического применения в областях, где не существует традиционной культуры инжира, даже после того как были разработаны теоретические основы. Предложение Кондита (Condit, 1947), одного из пионеров в этой области, было встречено неодобрительно, когда он предложил для получения урожая в Калифорнии ввести в культуру каприфигу и Blastophaga.
Другая сложность в опылении F. carica - сезонный характер средиземноморского климата. Цветение непрерывное, с тремя главными синхронизированными сезонами. Последний выводок личинок перезимовывает в галлах и появляется следующей весной.
Род Ficus огромный, содержит более 1000 видов; представители этого рода распространены по всем тропикам, а также в субтропических областях; среди них встречаются самые толстые в мире деревья и ползучие растения, крепко цепляющиеся за скалы. Циконии различаются более чем по 10 признакам, и лишь немногие виды изучены в настоящее время. Многочисленные вариации одного и того же типа, описанные Галилом и его сотрудниками, свидетельствуют о том, что еще слишком рано делать какие-либо выводы. Единственный вывод, который представляется вполне надежным, состоит в том, что для всего рода характерен один и тот же синдром опыления, в котором участвуют видоспецифические осы - представители семейства Agaonidae (Wiebes, 1963, 1966).
Семейство Moraceae, к которому принадлежит род Ficus, в основном анемофильно. Существование этого в высшей степени сложного энтомофильного синдрома удивительно, и с возможными оговорками на плохо изученное пока опыление у Dorstenia (Carauta, 1972), эволюцию этого синдрома проследить крайне трудно.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'